
18.4: Linfócitos B e anticorpos
- Page ID
- 181432

Objetivos de
- Descreva a produção e maturação das células B
- Compare a estrutura dos receptores de células B e receptores de células T
- Compare a ativação dependente de T e independente de T de células B
- Compare as respostas de anticorpos primários e secundários
A imunidade humoral se refere aos mecanismos das defesas imunes adaptativas que são mediados por anticorpos secretados pelos linfócitos B, ou células B. Esta seção se concentrará nas células B e discutirá sua produção e maturação, receptores e mecanismos de ativação.
Produção e maturação de células B
Como as células T, as células B são formadas a partir de células-tronco hematopoiéticas multipotentes (HSCs) na medula óssea e seguem um caminho através das células-tronco linfoides e dos linfoblastos (ver Figura 17.3.1). Ao contrário das células T, entretanto, os linfoblastos destinados a se tornarem células B não saem da medula óssea e viajam até o timo para maturação. Em vez disso, eventuais células B continuam a amadurecer na medula óssea.
A primeira etapa da maturação das células B é uma avaliação da funcionalidade de seus receptores de ligação ao antígeno. Isso ocorre por meio da seleção positiva de células B com receptores funcionais normais. Um mecanismo de seleção negativa é então usado para eliminar as células B autorreativas e minimizar o risco de autoimunidade. A seleção negativa de células B autorreativas pode envolver a eliminação por apoptose, edição ou modificação dos receptores para que não sejam mais autorreativos ou a indução de anergia na célula B. As células B imaturas que passam pela seleção na medula óssea viajam para o baço para seus estágios finais de maturação. Lá, elas se tornam células B maduras ingênuas, ou seja, células B maduras que ainda não foram ativadas.
Exercício\(\PageIndex{1}\)
Compare a maturação das células B com a maturação das células T.
Receptores de células B
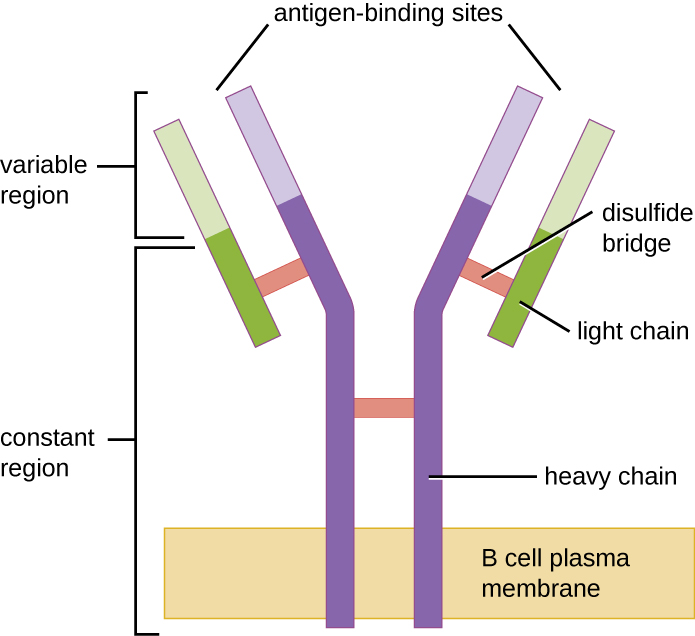
Como as células T, as células B possuem receptores antígenos específicos com especificidades diversas. Embora dependam de células T para um funcionamento ideal, as células B podem ser ativadas sem a ajuda das células T. Os receptores de células B (BCRs) para células B maduras ingênuas são formas monoméricas de IgD e IgM ligadas à membrana. Eles têm duas cadeias pesadas idênticas e duas cadeias leves idênticas conectadas por ligações de dissulfeto em uma forma básica de “Y” (Figura\(\PageIndex{1}\)). O tronco da molécula em forma de Y, a região constante das duas cadeias pesadas, atravessa a membrana da célula B. Os dois sítios de ligação ao antígeno expostos ao exterior da célula B estão envolvidos na ligação de epítopos específicos do patógeno para iniciar o processo de ativação. Estima-se que cada célula B madura ingênua tenha mais de 100.000 BCRs em sua membrana, e cada uma dessas BCRs tenha uma especificidade idêntica de ligação ao epítopo.
Para estarem preparadas para reagir a uma ampla variedade de epítopos microbianos, as células B, como as células T, usam o rearranjo genético de centenas de segmentos de genes para fornecer a diversidade necessária de especificidades do receptor. A região variável da cadeia pesada BCR é composta por segmentos V, D e J, semelhantes à cadeia β do TCR. A região variável da cadeia leve BCR é composta por segmentos V e J, semelhantes à cadeia α do TCR. O rearranjo genético de todas as combinações possíveis de V-J-D (cadeia pesada) e V-J (cadeia leve) fornece milhões de sítios únicos de ligação ao antígeno para o BCR e para os anticorpos secretados após a ativação.
Uma diferença importante entre BCRs e TCRs é a maneira como eles podem interagir com epítopos antigênicos. Enquanto os TCRs só podem interagir com epítopos antigênicos que são apresentados na fenda de ligação ao antígeno do MHC I ou MHC II, os BCRs não requerem apresentação de antígeno com o MHC; eles podem interagir com epítopos em antígenos livres ou com epítopos exibidos na superfície de patógenos intactos. Outra diferença importante é que os TCRs reconhecem apenas epítopos proteicos, enquanto os BCRs podem reconhecer epítopos associados a diferentes classes moleculares (por exemplo, proteínas, polissacarídeos, lipopolissacarídeos).
A ativação das células B ocorre por meio de diferentes mecanismos, dependendo da classe molecular do antígeno. A ativação de uma célula B por um antígeno proteico requer que a célula B funcione como um APC, apresentando os epítopos proteicos com MHC II para auxiliar as células T. Devido à sua dependência das células T para ativação das células B, os antígenos proteicos são classificados como antígenos dependentes de T. Em contraste, polissacarídeos, lipopolissacarídeos e outros antígenos não proteicos são considerados antígenos independentes de T porque podem ativar células B sem processamento e apresentação de antígenos às células T.
Exercício\(\PageIndex{2}\)
- Quais tipos de moléculas servem como BCR?
- Quais são as diferenças entre TCRs e BCRs em relação ao reconhecimento de antígenos?
- Quais classes de moléculas são antígenos dependentes de T e quais são antígenos independentes de T?
Ativação independente de células T de células B
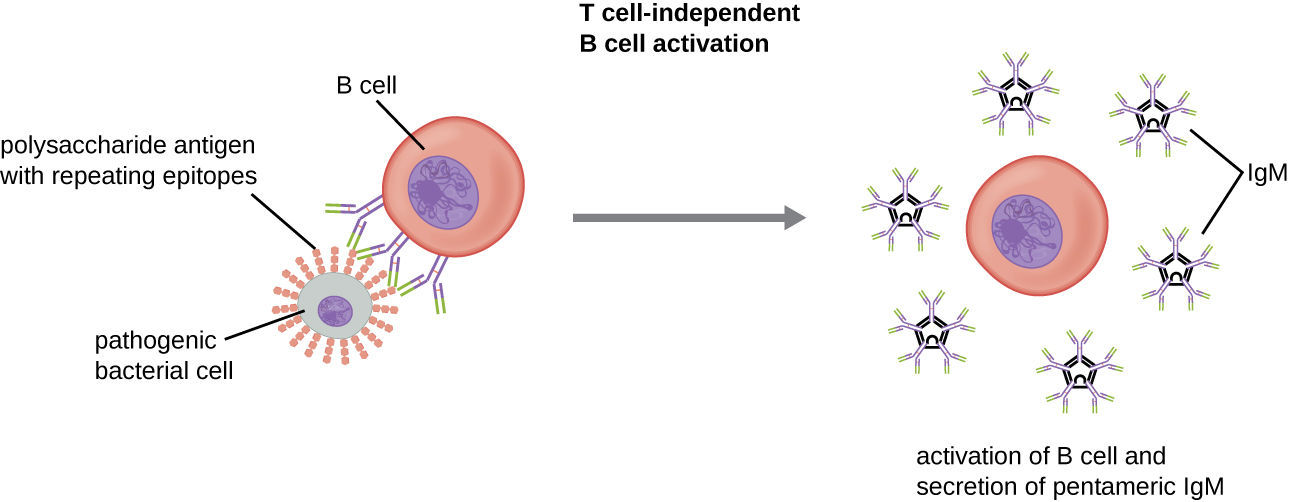
A ativação de células B sem a cooperação de células T auxiliares é chamada de ativação independente de células T e ocorre quando os BCRs interagem com antígenos independentes de T. Os antígenos independentes de T (por exemplo, cápsulas de polissacarídeos, lipopolissacarídeos) têm unidades de epítopo repetitivas dentro de sua estrutura, e essa repetição permite a ligação cruzada de vários BCRs, fornecendo o primeiro sinal de ativação (Figura\(\PageIndex{2}\)). Como as células T não estão envolvidas, o segundo sinal deve vir de outras fontes, como interações de receptores do tipo Toll com PAMPs ou interações com fatores do sistema complemento.
Uma vez que uma célula B é ativada, ela sofre proliferação clonal e as células-filhas se diferenciam em células plasmáticas. As células plasmáticas são fábricas de anticorpos que secretam grandes quantidades de anticorpos. Após a diferenciação, os BCRs de superfície desaparecem e a célula plasmática secreta moléculas pentaméricas de IgM que têm a mesma especificidade de antígeno que as BCRs (Figura\(\PageIndex{2}\)).
A resposta independente da célula T é de curta duração e não resulta na produção de células B de memória. Portanto, não resultará em uma resposta secundária às exposições subsequentes a antígenos independentes de T.
Exercício\(\PageIndex{3}\)
- Quais são os dois sinais necessários para a ativação independente de células T das células B?
- Qual é a função de uma célula plasmática?
Ativação dependente de células T de células B
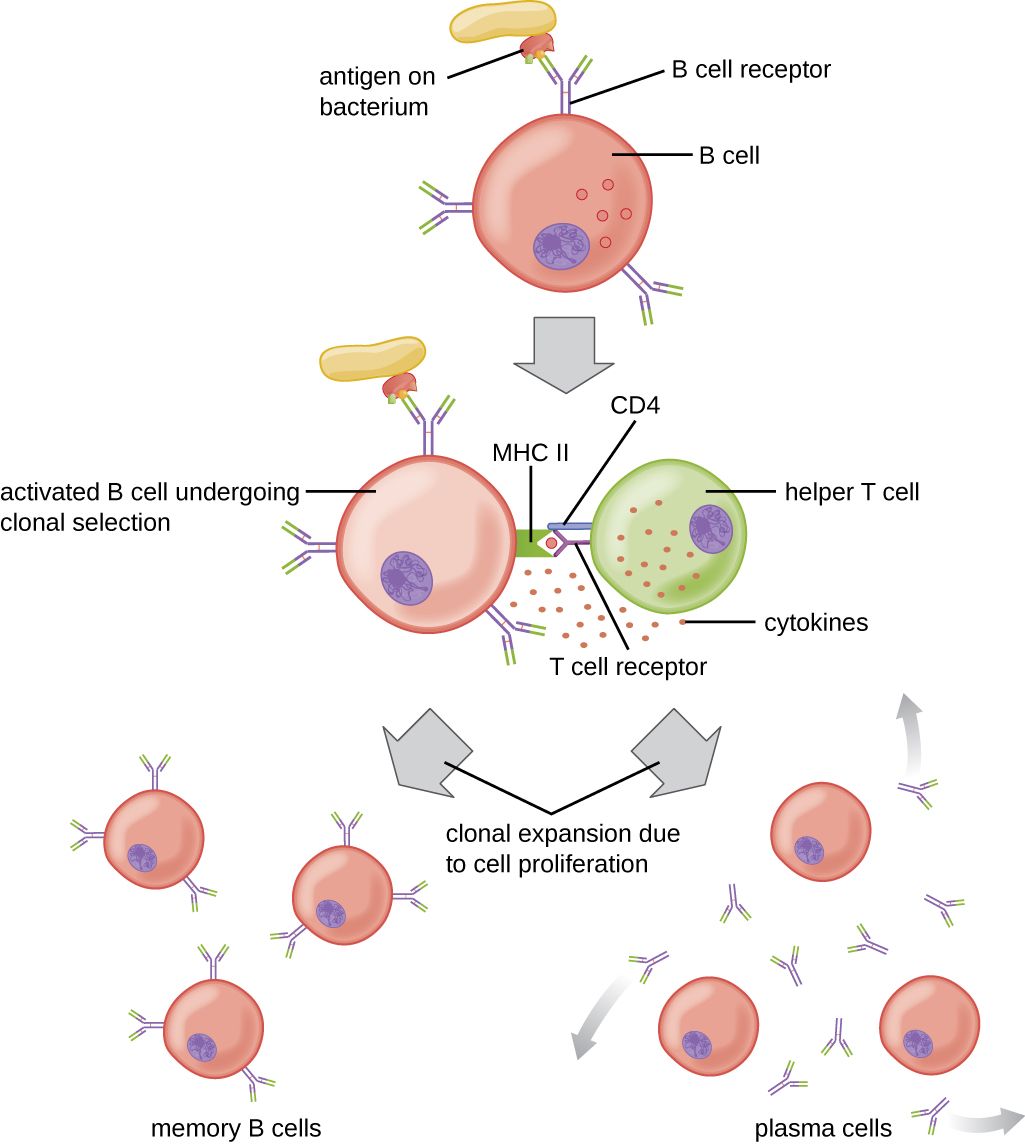
A ativação das células B dependente da célula T é mais complexa do que a ativação independente da célula T, mas a resposta imune resultante é mais forte e desenvolve a memória. A ativação dependente de células T pode ocorrer em resposta a antígenos proteicos livres ou a antígenos proteicos associados a um patógeno intacto. A interação entre os BCRs em uma célula B madura ingênua e um antígeno proteico livre estimula a internalização do antígeno, enquanto a interação com antígenos associados a um patógeno intacto inicia a extração do antígeno do patógeno antes da internalização. Uma vez internalizado dentro da célula B, o antígeno proteico é processado e apresentado com o MHC II. O antígeno apresentado é então reconhecido pelas células T auxiliares específicas do mesmo antígeno. O TCR da célula T auxiliar reconhece o antígeno estranho e a molécula CD4 da célula T interage com o MHC II na célula B. A coordenação entre células B e células T auxiliares que são específicas do mesmo antígeno é chamada de reconhecimento vinculado.
Uma vez ativadas por reconhecimento ligado, as células T H 2 produzem e secretam citocinas que ativam a célula B e causam proliferação em células-filhas clonais. Após várias rodadas de proliferação, citocinas adicionais fornecidas pelas células T H 2 estimulam a diferenciação de clones de células B ativadas em células B de memória, que responderão rapidamente às exposições subsequentes ao mesmo epítopo proteico, e células plasmáticas que perdem suas membranas BCRs e inicialmente secretam IgM pentamérico (Figura\(\PageIndex{3}\)).
Após a secreção inicial de IgM, as citocinas secretadas pelas células T H 2 estimulam as células plasmáticas a mudarem da produção de IgM para a produção de IgG, IgA ou IgE. Esse processo, chamado de troca de classe ou troca de isótipos, permite que células plasmáticas clonadas da mesma célula B ativada produzam uma variedade de classes de anticorpos com a mesma especificidade do epítopo. A mudança de classe é realizada pelo rearranjo genético dos segmentos genéticos que codificam a região constante, o que determina a classe de um anticorpo. A região variável não é alterada, então a nova classe de anticorpo mantém a especificidade original do epítopo.
Exercício\(\PageIndex{4}\)
- Quais etapas são necessárias para a ativação das células B dependente de células T?
- O que é a mudança de classe de anticorpos e por que ela é importante?
Respostas primárias e secundárias
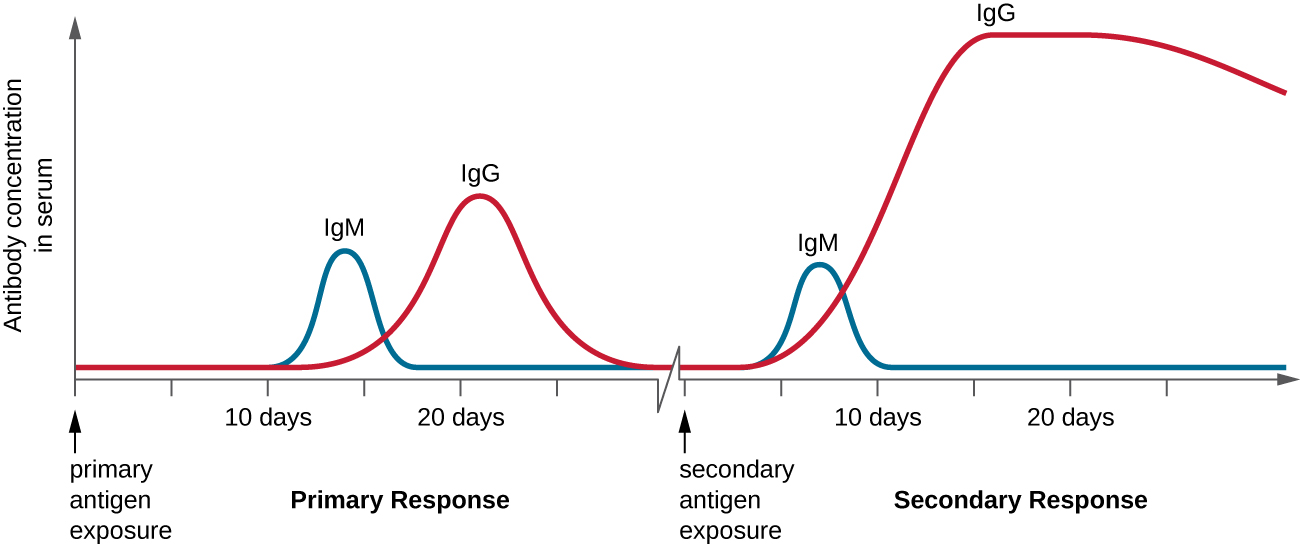
A ativação das células B dependente de células T desempenha um papel importante nas respostas primárias e secundárias associadas à imunidade adaptativa. Com a primeira exposição a um antígeno proteico, ocorre uma resposta de anticorpo primário dependente de células T. O estágio inicial da resposta primária é um período de atraso, ou período latente, de aproximadamente 10 dias, durante o qual nenhum anticorpo pode ser detectado no soro. Esse período de atraso é o tempo necessário para todas as etapas da resposta primária, incluindo a ligação do antígeno às células B maduras ingênuas com BCRs, processamento e apresentação do antígeno, ativação de células T auxiliares, ativação de células B e proliferação clonal. O final do período de defasagem é caracterizado por um aumento nos níveis de IgM no soro, pois as células T H 2 estimulam a diferenciação das células B em células plasmáticas. Os níveis de IgM atingem seu pico cerca de 14 dias após a exposição primária ao antígeno; mais ou menos nessa mesma época, o T H 2 estimula a mudança de classe de anticorpos e os níveis de IgM no soro começam a diminuir. Enquanto isso, os níveis de IgG aumentam até atingirem um pico cerca de três semanas após a resposta primária (Figura\(\PageIndex{4}\)).
Durante a resposta primária, algumas das células B clonadas são diferenciadas em células B de memória programadas para responder às exposições subsequentes. Essa resposta secundária ocorre com mais rapidez e força do que a resposta primária. O período de defasagem é reduzido para apenas alguns dias e a produção de IgG é significativamente maior do que a observada para a resposta primária (Figura\(\PageIndex{4}\)). Além disso, os anticorpos produzidos durante a resposta secundária são mais eficazes e se ligam com maior afinidade aos epítopos alvo. As células plasmáticas produzidas durante as respostas secundárias vivem mais do que as produzidas durante a resposta primária, portanto, os níveis de anticorpos específicos permanecem elevados por um longo período de tempo.
Exercício\(\PageIndex{5}\)
- Quais eventos ocorrem durante o período de atraso da resposta primária do anticorpo?
- Por que os níveis de anticorpos permanecem elevados por mais tempo durante a resposta secundária de anticorpos?
Conceitos principais e resumo
- Os linfócitos B ou células B produzem anticorpos envolvidos na imunidade humoral. As células B são produzidas na medula óssea, onde ocorrem os estágios iniciais de maturação, e viajam para o baço para as etapas finais de maturação em células B maduras ingênuas.
- Os receptores de células B (BCRs) são formas monoméricas de IgD e IgM ligadas à membrana que ligam epítopos de antígenos específicos com suas regiões de ligação ao antígeno Fab. A diversidade da especificidade de ligação ao antígeno é criada pelo rearranjo genético dos segmentos V, D e J semelhante ao mecanismo usado para a diversidade de TCR.
- Os antígenos proteicos são chamados de antígenos dependentes de T porque só podem ativar células B com a cooperação de células T auxiliares. Outras classes de moléculas não requerem cooperação com células T e são chamadas de antígenos independentes de T.
- A ativação independente de células B por células T envolve a ligação cruzada de BCRs por epítopos repetitivos de antígenos não proteicos. É caracterizada pela produção de IgM pelas células plasmáticas e não produz células B de memória.
- A ativação de células B dependente de células T envolve processamento e apresentação de antígenos proteicos para células T auxiliares, ativação das células B por citocinas secretadas por células T H 2 ativadas e células plasmáticas que produzem diferentes classes de anticorpos como resultado da classe trocando. Células B de memória também são produzidas.
- Exposições secundárias a antígenos dependentes de T resultam em uma resposta secundária de anticorpos iniciada pelas células B de memória. A resposta secundária se desenvolve mais rapidamente e produz níveis mais altos e mais sustentados de anticorpos com maior afinidade pelo antígeno específico.