
17.2: Defesas químicas
- Page ID
- 181146

objetivos de aprendizagem
- Descreva como as enzimas nos fluidos corporais fornecem proteção contra infecções ou doenças
- Listar e descrever a função dos peptídeos antimicrobianos, componentes do complemento, citocinas e proteínas de fase aguda
- Descreva semelhanças e diferenças entre as vias clássicas, alternativas e de complemento de lectina
Além das defesas físicas, o sistema imunológico inato inespecífico usa vários mediadores químicos que inibem os invasores microbianos. O termo “mediadores químicos” abrange uma grande variedade de substâncias encontradas em vários fluidos e tecidos corporais em todo o corpo. Os mediadores químicos podem trabalhar sozinhos ou em conjunto para inibir a colonização e a infecção microbiana.
Alguns mediadores químicos são produzidos endogenamente, o que significa que são produzidos pelas células do corpo humano; outros são produzidos exogenamente, o que significa que são produzidos por certos micróbios que fazem parte do microbioma. Alguns mediadores são produzidos continuamente, banhando a área com a substância antimicrobiana; outros são produzidos ou ativados principalmente em resposta a algum estímulo, como a presença de micróbios.
Mediadores químicos e enzimáticos encontrados em fluidos corporais
Os fluidos produzidos pela pele incluem exemplos de mediadores endógenos e exógenos. As glândulas sebáceas da derme secretam um óleo chamado sebo que é liberado na superfície da pele através dos folículos pilosos. Esse sebo é um mediador endógeno, fornecendo uma camada adicional de defesa ao ajudar a selar os poros do folículo piloso, impedindo que bactérias na superfície da pele invadam as glândulas sudoríparas e os tecidos circundantes (Figura\(\PageIndex{1}\)). Certos membros do microbioma, como a bactéria Propionibacterium acnes e o fungo Malassezia, entre outros, podem usar enzimas lipases para degradar o sebo, usando-o como fonte de alimento. Isso produz ácido oleico, que cria um ambiente levemente ácido na superfície da pele que é inóspito para muitos micróbios patogênicos. O ácido oleico é um exemplo de mediador produzido exogenamente porque é produzido por micróbios residentes e não diretamente pelas células do corpo.
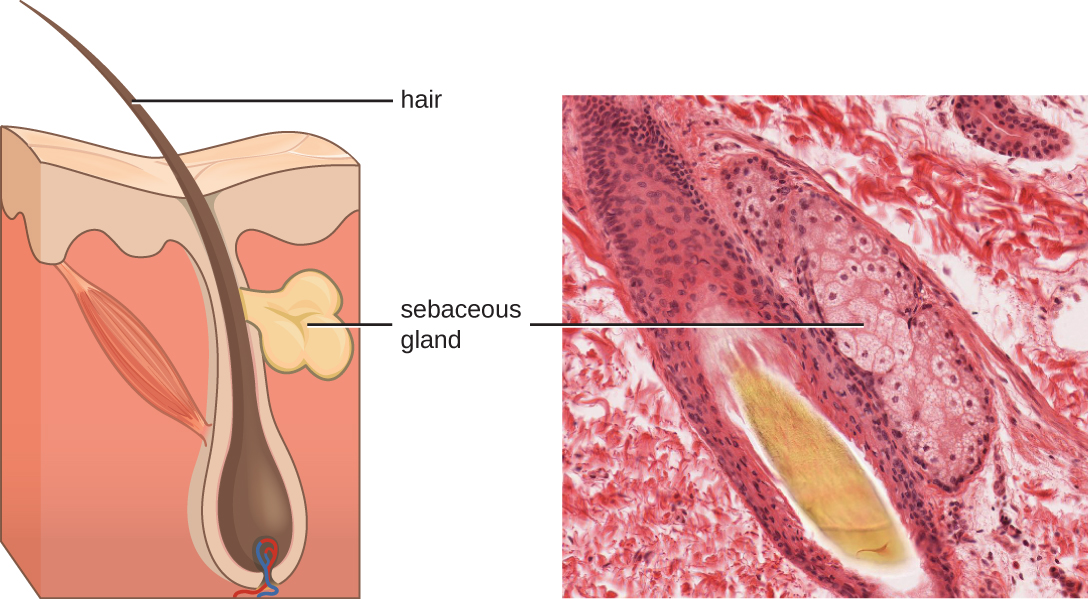
Fatores ambientais que afetam a microbiota da pele podem ter um impacto direto na produção de mediadores químicos. A baixa umidade ou a diminuição da produção de sebo, por exemplo, podem tornar a pele menos habitável para micróbios que produzem ácido oleico, tornando a pele mais suscetível a patógenos normalmente inibidos pelo baixo pH da pele. Muitos hidratantes para a pele são formulados para combater esses efeitos, restaurando a umidade e os óleos essenciais da pele.
O trato digestivo também produz um grande número de mediadores químicos que inibem ou matam micróbios. Na cavidade oral, a saliva contém mediadores, como as enzimas lactoperoxidase, e o muco secretado pelo esôfago contém a enzima antibacteriana lisozima. No estômago, o fluido gástrico altamente ácido mata a maioria dos micróbios. No trato digestivo inferior, os intestinos têm enzimas pancreáticas e intestinais, peptídeos antibacterianos (criptinas), bile produzida pelo fígado e células Paneth especializadas que produzem lisozima. Juntos, esses mediadores são capazes de eliminar a maioria dos patógenos que conseguem sobreviver ao ambiente ácido do estômago.
No trato urinário, a urina expulsa os micróbios do corpo durante a micção. Além disso, a leve acidez da urina (o pH médio é de cerca de 6) inibe o crescimento de muitos micróbios e patógenos potenciais no trato urinário.
O sistema reprodutor feminino emprega lactato, um mediador químico produzido exogenamente, para inibir o crescimento microbiano. As células e camadas de tecido que compõem a vagina produzem glicogênio, um polímero de glicose ramificado e mais complexo. Os lactobacilos na área fermentam o glicogênio para produzir lactato, diminuindo o pH da vagina e inibindo a microbiota transitória, patógenos oportunistas como a Candida (uma levedura associada a infecções vaginais) e outros patógenos responsáveis por doenças sexualmente transmissíveis.
Nos olhos, as lágrimas contêm os mediadores químicos lisozima e lactoferrina, ambos capazes de eliminar micróbios que chegaram à superfície dos olhos. A lisozima cliva a ligação entre NAG e NAM no peptidoglicano, um componente da parede celular das bactérias. É mais eficaz contra bactérias gram-positivas, que não possuem a membrana externa protetora associada às bactérias gram-negativas. A lactoferrina inibe o crescimento microbiano ao se ligar quimicamente e sequestrar o ferro. Isso efetivamente mata muitos micróbios que precisam de ferro para crescer.
Nas orelhas, o cerúmen (cera de ouvido) apresenta propriedades antimicrobianas devido à presença de ácidos graxos, que reduzem o pH para entre 3 e 5.
O trato respiratório usa vários mediadores químicos nas vias nasais, na traquéia e nos pulmões. O muco produzido nas passagens nasais contém uma mistura de moléculas antimicrobianas semelhantes às encontradas nas lágrimas e na saliva (por exemplo, lisozima, lactoferrina, lactoperoxidase). As secreções na traquéia e nos pulmões também contêm lisozima e lactoferrina, bem como um grupo diversificado de mediadores químicos adicionais, como o complexo lipoproteico chamado surfactante, que tem propriedades antibacterianas.
Exercício\(\PageIndex{1}\)
- Explicar a diferença entre mediadores endógenos e exógenos
- Descreva como o pH afeta as defesas antimicrobianas
Peptídeos antimicrobianos
Os peptídeos antimicrobianos (AMPs) são uma classe especial de mediadores não específicos derivados de células com propriedades antimicrobianas de amplo espectro. Alguns AMPs são produzidos rotineiramente pelo corpo, enquanto outros são produzidos principalmente (ou produzidos em maiores quantidades) em resposta à presença de um patógeno invasor. A pesquisa começou a explorar como os AMPs podem ser usados no diagnóstico e tratamento de doenças.
Os AMPs podem induzir danos celulares em microrganismos de várias maneiras, inclusive infligindo danos às membranas, destruindo DNA e RNA ou interferindo na síntese da parede celular. Dependendo do mecanismo antimicrobiano específico, um AMP específico pode inibir apenas certos grupos de micróbios (por exemplo, bactérias gram-positivas ou gram-negativas) ou pode ser mais amplamente eficaz contra bactérias, fungos, protozoários e vírus. Muitos AMPs são encontrados na pele, mas também podem ser encontrados em outras regiões do corpo.
Uma família de AMPs chamada defensinas pode ser produzida por células epiteliais em todo o corpo, bem como por defesas celulares, como macrófagos e neutrófilos (consulte Defesas celulares). As defensinas podem ser secretadas ou agir dentro das células hospedeiras; elas combatem os microrganismos danificando suas membranas plasmáticas. As AMPs chamadas bacteriocinas são produzidas exogenamente por certos membros da microbiota residente no trato gastrointestinal. Os genes que codificam esses tipos de AMPs são frequentemente transportados em plasmídeos e podem ser transmitidos entre diferentes espécies dentro da microbiota residente por meio de transferência lateral ou horizontal de genes.
Existem vários outros AMPs em todo o corpo. As características de alguns dos AMPs mais significativos estão resumidas na Tabela\(\PageIndex{1}\).
| AMP | Secretado por | Site do corpo | Patógenos inibidos | Modo de ação |
|---|---|---|---|---|
| Bacteriocinas | Microbiota residente | Tracto gastrointestinal | Bactérias | Interromper a membrana |
| Catelicidina | Células epiteliais, macrófagos e outros tipos celulares | Pele | Bactérias e fungos | Interrompe a membrana |
| Defensas | Células epiteliais, macrófagos, neutrófilos | Em todo o corpo | Fungos, bactérias e muitos vírus | Interromper a membrana |
| Dermicidina | glândulas sudoríparas | Pele | Bactérias e fungos | Interrompe a integridade da membrana e os canais iônicos |
| Histatinas | Glândulas salivares | Cavidade oral | Fungos | Interromper a função intracelular |
Exercício\(\PageIndex{2}\)
Por que os peptídeos antimicrobianos (AMPs) são considerados defesas não específicas?
Mediadores de proteína plasmática
Muitos fatores imunes inatos não específicos são encontrados no plasma, a porção fluida do sangue. O plasma contém eletrólitos, açúcares, lipídios e proteínas, cada um dos quais ajuda a manter a homeostase (ou seja, o funcionamento interno estável do corpo) e contém as proteínas envolvidas na coagulação do sangue. Proteínas adicionais encontradas no plasma sanguíneo, como proteínas de fase aguda, proteínas do complemento e citocinas, estão envolvidas na resposta imune inata não específica.
Plasma versus soro
Existem dois termos para a porção fluida do sangue: plasma e soro. Como eles diferem se ambos são fluidos e não têm células? A porção fluida do sangue que sobra após a coagulação (coagulação das células sanguíneas) é soro. Embora moléculas como muitas vitaminas, eletrólitos, certos açúcares, proteínas do complemento e anticorpos ainda estejam presentes no soro, os fatores de coagulação estão amplamente esgotados. O plasma, por outro lado, ainda contém todos os elementos de coagulação. Para obter plasma do sangue, um anticoagulante deve ser usado para evitar a coagulação. Exemplos de anticoagulantes incluem heparina e ácido etilenodiamina tetraacético (EDTA). Como a coagulação é inibida, uma vez obtida, a amostra deve ser gentilmente girada para baixo em uma centrífuga. As células sanguíneas mais pesadas e densas formam uma pastilha na parte inferior de um tubo de centrífuga, enquanto a porção de plasma fluido, que é mais leve e menos densa, permanece acima do sedimento celular.
Proteínas de fase aguda
As proteínas de fase aguda são outra classe de mediadores antimicrobianos. As proteínas de fase aguda são produzidas principalmente no fígado e secretadas no sangue em resposta às moléculas inflamatórias do sistema imunológico. Exemplos de proteínas de fase aguda incluem proteína C reativa, amilóide A sérica, ferritina, transferrina, fibrinogênio e lectina de ligação à manose. Cada uma dessas proteínas tem uma estrutura química diferente e inibe ou destrói micróbios de alguma forma (Tabela\(\PageIndex{1}\)).
| Algumas proteínas de fase aguda e suas funções | |
|---|---|
| Proteína C-reativa | Reveste bactérias (opsonização), preparando-as para ingestão por fagócitos |
| Amilóide sérica A | |
| Ferritina | Ligar e sequestrar ferro, inibindo assim o crescimento de patógenos |
| Transferrina | |
| Fibrinogênio | Envolvido na formação de coágulos sanguíneos que capturam patógenos bacterianos |
| Lectina de ligação à manose | Ativa a cascata do complemento |
O sistema Complement
O sistema complemento é um grupo de mediadores de proteínas plasmáticas que podem atuar como uma defesa inata inespecífica e, ao mesmo tempo, servir para conectar a imunidade inata e adaptativa (discutida no próximo capítulo). O sistema complemento é composto por mais de 30 proteínas (incluindo C1 a C9) que normalmente circulam como proteínas precursoras no sangue. Essas proteínas precursoras são ativadas quando estimuladas ou desencadeadas por uma variedade de fatores, incluindo a presença de microrganismos. As proteínas do complemento são consideradas parte da imunidade inata inespecífica porque estão sempre presentes no sangue e nos fluidos teciduais, permitindo que sejam ativadas rapidamente. Além disso, quando ativadas por meio da via alternativa (descrita posteriormente nesta seção), as proteínas do complemento têm como alvo os patógenos de maneira não específica.
O processo pelo qual os precursores circulantes do complemento se tornam funcionais é chamado de ativação do complemento. Esse processo é uma cascata que pode ser desencadeada por um dos três mecanismos diferentes, conhecidos como vias alternativa, clássica e da lectina.
A via alternativa é iniciada pela ativação espontânea da proteína C3 do complemento. A hidrólise do C3 produz dois produtos, C3a e C3b. Quando nenhum micróbio invasor está presente, o C3b é rapidamente degradado em uma reação de hidrólise usando a água do sangue. No entanto, se micróbios invasores estiverem presentes, o C3b se fixa à superfície desses micróbios. Uma vez conectado, o C3b recrutará outras proteínas do complemento em uma cascata (Figura\(\PageIndex{2}\)).
A via clássica fornece um mecanismo mais eficiente de ativação da cascata do complemento, mas depende da produção de anticorpos pelas defesas imunes adaptativas específicas. Para iniciar a via clássica, um anticorpo específico deve primeiro se ligar ao patógeno para formar um complexo anticorpo-antígeno. Isso ativa a primeira proteína na cascata do complemento, o complexo C1. O complexo C1 é um complexo proteico com várias partes, e cada componente participa da ativação total do complexo geral. Após o recrutamento e ativação do complexo C1, as demais proteínas do complemento da via clássica são recrutadas e ativadas em uma sequência em cascata (Figura\(\PageIndex{2}\)).
A via de ativação da lectina é semelhante à via clássica, mas é desencadeada pela ligação da lectina de ligação à manose, uma proteína de fase aguda, aos carboidratos na superfície microbiana. Como outras proteínas de fase aguda, as lectinas são produzidas pelas células do fígado e geralmente são reguladas positivamente em resposta aos sinais inflamatórios recebidos pelo corpo durante uma infecção (Figura\(\PageIndex{2}\)).
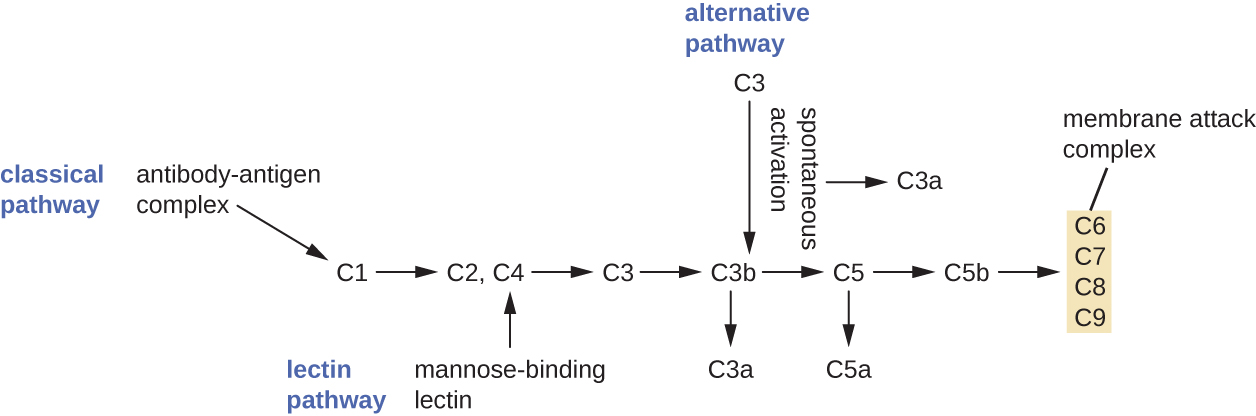
Embora cada via de ativação do complemento seja iniciada de forma diferente, todas elas fornecem os mesmos resultados protetores: opsonização, inflamação, quimiotaxia e citólise. O termo opsonização se refere ao revestimento de um patógeno por uma substância química (chamada opsonina) que permite que as células fagocíticas o reconheçam, engolfem e destruam com mais facilidade. As opsoninas da cascata do complemento incluem C1q, C3b e C4b. Outras opsoninas importantes incluem proteínas e anticorpos de ligação à manose. Os fragmentos do complemento C3a e C5a são anafilatoxinas bem caracterizadas com potentes funções pró-inflamatórias. As anafilatoxinas ativam os mastócitos, causando a degranulação e a liberação de sinais químicos inflamatórios, incluindo mediadores que causam vasodilatação e aumento da permeabilidade vascular. O C5a também é um dos quimioatrativos mais potentes para neutrófilos e outros glóbulos brancos, defesas celulares que serão discutidas na próxima seção.
As proteínas do complemento C6, C7, C8 e C9 se reúnem em um complexo de ataque de membrana (MAC), que permite que o C9 se polimerize em poros nas membranas de bactérias gram-negativas. Esses poros permitem que água, íons e outras moléculas entrem e saiam livremente das células-alvo, levando eventualmente à lise celular e à morte do patógeno (Figura\(\PageIndex{2}\)). No entanto, o MAC só é eficaz contra bactérias gram-negativas; ele não consegue penetrar na espessa camada de peptidoglicano associada às paredes celulares das bactérias gram-positivas. Como o MAC não representa uma ameaça letal aos patógenos bacterianos gram-positivos, a opsonização mediada pelo complemento é mais importante para sua eliminação.
Citocinas
As citocinas são proteínas solúveis que atuam como sinais de comunicação entre as células. Em uma resposta imune inata inespecífica, várias citocinas podem ser liberadas para estimular a produção de mediadores químicos ou outras funções celulares, como proliferação celular, diferenciação celular, inibição da divisão celular, apoptose e quimiotaxia.
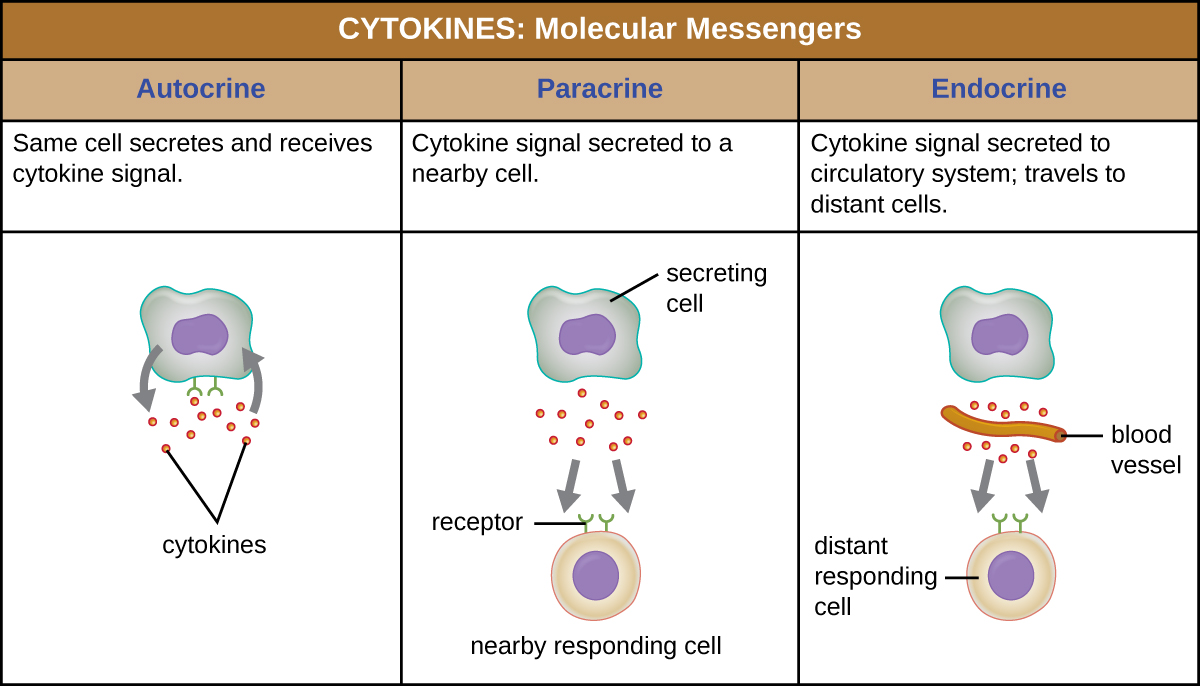
Quando uma citocina se liga ao seu receptor alvo, o efeito pode variar amplamente, dependendo do tipo de citocina e do tipo de célula ou receptor ao qual ela se ligou. A função de uma citocina específica pode ser descrita como autócrina, parácrina ou endócrina (Figura\(\PageIndex{3}\)). Na função autócrina, a mesma célula que libera a citocina é a receptora do sinal; em outras palavras, a função autócrina é uma forma de autoestimulação por uma célula. Em contraste, a função parácrina envolve a liberação de citocinas de uma célula para outras células próximas, estimulando alguma resposta das células receptoras. Por último, a função endócrina ocorre quando as células liberam citocinas na corrente sanguínea para serem transportadas para as células-alvo muito mais distantes.
Três classes importantes de citocinas são as interleucinas, quimiocinas e interferons. Originalmente, pensava-se que as interleucinas eram produzidas apenas por leucócitos (glóbulos brancos) e estimulavam apenas os leucócitos, daí as razões de seu nome. Embora as interleucinas estejam envolvidas na modulação de quase todas as funções do sistema imunológico, seu papel no corpo não se restringe à imunidade. As interleucinas também são produzidas e estimulam uma variedade de células não relacionadas às defesas imunológicas.
As quimiocinas são fatores quimiotáticos que recrutam leucócitos para locais de infecção, danos nos tecidos e inflamação. Em contraste com fatores quimiotáticos mais gerais, como o fator de complemento C5a, as quimiocinas são muito específicas nos subconjuntos de leucócitos que recrutam.
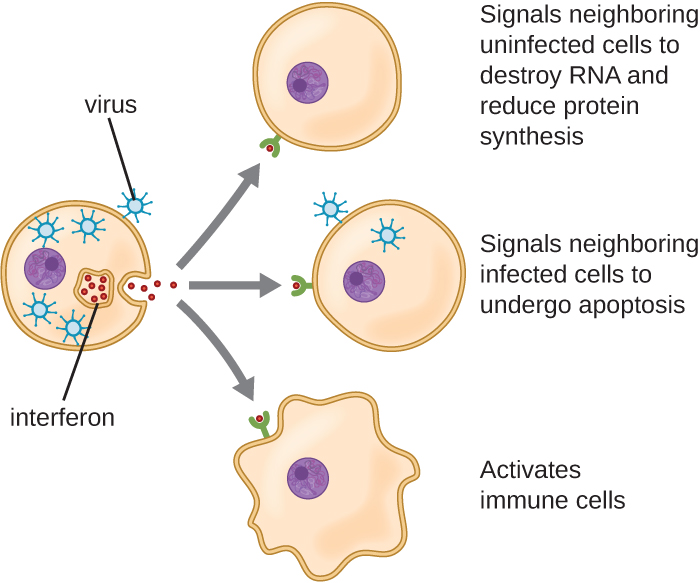
Os interferões são um grupo diverso de moléculas de sinalização imune e são especialmente importantes em nossa defesa contra vírus. Os interferões do tipo I (interferon-α e interferon-β) são produzidos e liberados por células infectadas com vírus. Esses interferons estimulam as células próximas a interromper a produção de mRNA, destruir o RNA já produzido e reduzir a síntese protéica. Essas alterações celulares inibem a replicação viral e a produção de vírus maduros, retardando a propagação do vírus. Os interferões do tipo I também estimulam várias células imunológicas envolvidas na depuração viral a atacar de forma mais agressiva as células infectadas pelo vírus. O interferon tipo II (interferon-γ) é um importante ativador das células imunes (Figura\(\PageIndex{4}\)).
Mediadores que provocam inflamação
Muitos dos mediadores químicos discutidos nesta seção contribuem de alguma forma para a inflamação e a febre, que são respostas imunes inespecíficas discutidas com mais detalhes em Inflamação e Febre. As citocinas estimulam a produção de proteínas de fase aguda, como proteína C reativa e lectina de ligação à manose, no fígado. Essas proteínas de fase aguda atuam como opsoninas, ativando as cascatas do complemento pela via da lectina.
Algumas citocinas também se ligam aos mastócitos e basófilos, induzindo-os a liberar histamina, um composto pró-inflamatório. Os receptores de histamina são encontrados em uma variedade de células e mediam eventos pró-inflamatórios, como broncoconstrição (contração das vias aéreas) e contração do músculo liso.
Além da histamina, os mastócitos podem liberar outros mediadores químicos, como os leucotrienos. Os leucotrienos são mediadores pró-inflamatórios à base de lipídios que são produzidos a partir do metabolismo do ácido araquidônico na membrana celular de leucócitos e células teciduais. Em comparação com os efeitos pró-inflamatórios da histamina, os dos leucotrienos são mais potentes e duradouros. Juntos, esses mediadores químicos podem induzir tosse, vômito e diarreia, que servem para expulsar patógenos do corpo.
Certas citocinas também estimulam a produção de prostaglandinas, mediadores químicos que promovem os efeitos inflamatórios das cininas e histaminas. As prostaglandinas também podem ajudar a elevar a temperatura corporal, levando à febre, que promove as atividades dos glóbulos brancos e inibe levemente o crescimento de micróbios patogênicos (ver Inflamação e Febre).
Outro mediador inflamatório, a bradicinina, contribui para o edema, que ocorre quando fluidos e leucócitos vazam da corrente sanguínea para os tecidos. Ele se liga aos receptores nas células das paredes capilares, fazendo com que os capilares se dilatem e se tornem mais permeáveis aos fluidos.
Exercício\(\PageIndex{3}\)
- O que as três vias de ativação do complemento têm em comum?
- Explique os sinais autócrinos, parácrinos e endócrinos.
- Cite dois mediadores importantes que provocam inflamação.
Foco clínico: Parte 2
Para aliviar a constrição das vias aéreas, Angela é imediatamente tratada com anti-histamínicos e administrada corticosteróides por meio de um inalador e, em seguida, monitorada por um período de tempo. Embora sua condição não piore, as drogas não parecem estar aliviando sua condição. Ela é internada no hospital para posterior observação, testes e tratamento.
Após a admissão, um médico realiza testes de alergia para tentar determinar se algo em seu ambiente pode estar desencadeando uma resposta inflamatória alérgica. Um médico solicita análises de sangue para verificar os níveis de citocinas específicas. Uma amostra de escarro também é coletada e enviada ao laboratório para coloração microbiana, cultura e identificação de patógenos que podem estar causando uma infecção.
Exercício\(\PageIndex{4}\)
- Quais aspectos do sistema imunológico inato podem estar contribuindo para a constrição das vias aéreas de Angela?
- Por que Angela foi tratada com anti-histamínicos?
- Por que o médico estaria interessado nos níveis de citocinas no sangue de Angela?
\(\PageIndex{3}\)A tabela fornece um resumo das defesas químicas discutidas nesta seção.
| Defesa | Exemplos | Função |
|---|---|---|
| Produtos químicos e enzimas nos fluidos corporais | Sebo das glândulas sebáceas | Fornece barreira de óleo que protege os poros do folículo piloso contra patógenos |
| Ácido oleico do sebo e da microbiota da pele | Diminui o pH para inibir patógenos | |
| Lisozima nas secreções | Mata bactérias atacando a parede celular | |
| Ácido no estômago, urina e vagina | Inibe ou mata bactérias | |
| Enzimas digestivas e bile | Mate bactérias | |
| Lactoferrina e transferrina | Ligar e sequestrar ferro, inibindo o crescimento bacteriano | |
| Surfactante nos pulmões | Mata bactérias | |
| Peptídeos antimicrobianos | Defeninas, bacteriocinas, dermicidina, catelicidina, histatinas, | Mate as bactérias atacando as membranas ou interferindo nas funções celulares |
| Mediadores de proteínas plasmáticas | Proteínas de fase aguda (proteína C reativa, amilóide A sérica, ferritina, fibrinogênio, transferrina e lectina de ligação à manose) | Inibe o crescimento de bactérias e auxilia na captura e morte de bactérias |
| Complementa C3b e C4b | Opsonização de patógenos para auxiliar na fagocitose | |
| Complemento C5a | Quimioatrativo para fagócitos | |
| Complementa C3a e C5a | Anafilatoxinas pró-inflamatórias | |
| Citocinas | Interleucinas | Estimule e module a maioria das funções do sistema imunológico |
| Quimiocinas | Recrutar glóbulos brancos para a área infectada | |
| Interferões | Alerte as células sobre a infecção viral, induza a apoptose das células infectadas por vírus, induza defesas antivirais em células infectadas e não infectadas próximas, estimule as células imunes a atacar as células infectadas por vírus | |
| Mediadores que provocam inflamação | Histamina | Promove vasodilatação, broncoconstrição, contração do músculo liso, aumento da secreção e produção de muco |
| Leucotrienos | Promover a inflamação; mais forte e duradouro do que a histamina | |
| Prostaglandinas | Promover inflamação e febre | |
| Bradicinina | Aumenta a vasodilatação e a permeabilidade vascular, levando ao edema |
Conceitos principais e resumo
- Numerosos mediadores químicos produzidos de forma endógena e exógena exibem funções antimicrobianas inespecíficas.
- Muitos mediadores químicos são encontrados em fluidos corporais, como sebo, saliva, muco, fluidos gástricos e intestinais, urina, lágrimas, cerúmen e secreções vaginais.
- Os peptídeos antimicrobianos (AMPs) encontrados na pele e em outras áreas do corpo são amplamente produzidos em resposta à presença de patógenos. Isso inclui dermcidina, catelicidina, defensinas, histatinas e bacteriocinas.
- O plasma contém várias proteínas que servem como mediadores químicos, incluindo proteínas de fase aguda, proteínas do complemento e citocinas.
- O sistema complemento envolve várias proteínas precursoras que circulam no plasma. Essas proteínas são ativadas em uma sequência em cascata na presença de micróbios, resultando na opsonização de patógenos, quimioatração de leucócitos, indução de inflamação e citólise por meio da formação de um complexo de ataque de membrana (MAC).
- As citocinas são proteínas que facilitam várias respostas não específicas das células imunes inatas, incluindo produção de outros mediadores químicos, proliferação celular, morte celular e diferenciação.
- As citocinas desempenham um papel fundamental na resposta inflamatória, desencadeando a produção de mediadores que provocam a inflamação, como proteínas de fase aguda, histamina, leucotrienos, prostaglandinas e bradicinina.