
11.6: Como os procariontes assexuados alcançam a diversidade genética
- Page ID
- 181344

Objetivos de
- Compare os processos de transformação, transdução e conjugação
- Explicar como a transferência assexual de genes resulta na diversidade genética procariótica
- Explicar a estrutura e as consequências para a diversidade genética bacteriana dos transposons
Normalmente, quando consideramos a transferência genética, pensamos na transferência vertical de genes, a transmissão da informação genética de geração em geração. A transferência vertical de genes é, de longe, o principal modo de transmissão da informação genética em todas as células. Em organismos que se reproduzem sexualmente, eventos de cruzamento e uma variedade independente de cromossomos individuais durante a meiose contribuem para a diversidade genética da população. A diversidade genética também é introduzida durante a reprodução sexual, quando as informações genéticas de dois pais, cada um com diferentes complementos de informação genética, são combinadas, produzindo novas combinações de genótipos parentais na prole diploide. A ocorrência de mutações também contribui para a diversidade genética em uma população. A diversidade genética da prole é útil em ambientes variáveis ou inconsistentes e pode ser uma das razões para o sucesso evolutivo da reprodução sexual.
Quando procariontes e eucariotos se reproduzem assexuadamente, eles transferem uma cópia quase idêntica de seu material genético para seus filhos por meio da transferência vertical de genes. Embora a reprodução assexuada produza mais filhos mais rapidamente, quaisquer benefícios da diversidade entre esses filhos são perdidos. Como então os organismos cujo modo reprodutivo dominante é o assexual criam diversidade genética? Em procariontes, a transferência horizontal de genes (HGT), a introdução de material genético de um organismo para outro na mesma geração, é uma forma importante de introduzir a diversidade genética. O HGT permite que até mesmo espécies distantes compartilhem genes, influenciando seus fenótipos. Acredita-se que o HGT seja mais prevalente em procariontes, mas que apenas uma pequena fração do genoma procariótico pode ser transferida por esse tipo de transferência a qualquer momento. À medida que o fenômeno é investigado mais detalhadamente, pode se revelar ainda mais comum. Muitos cientistas acreditam que o HGT e a mutação são fontes significativas de variação genética, a matéria-prima para o processo de seleção natural, em procariontes. Embora o HGT seja mais comum entre organismos relacionados à evolução, ele pode ocorrer entre quaisquer duas espécies que vivam juntas em uma comunidade natural.
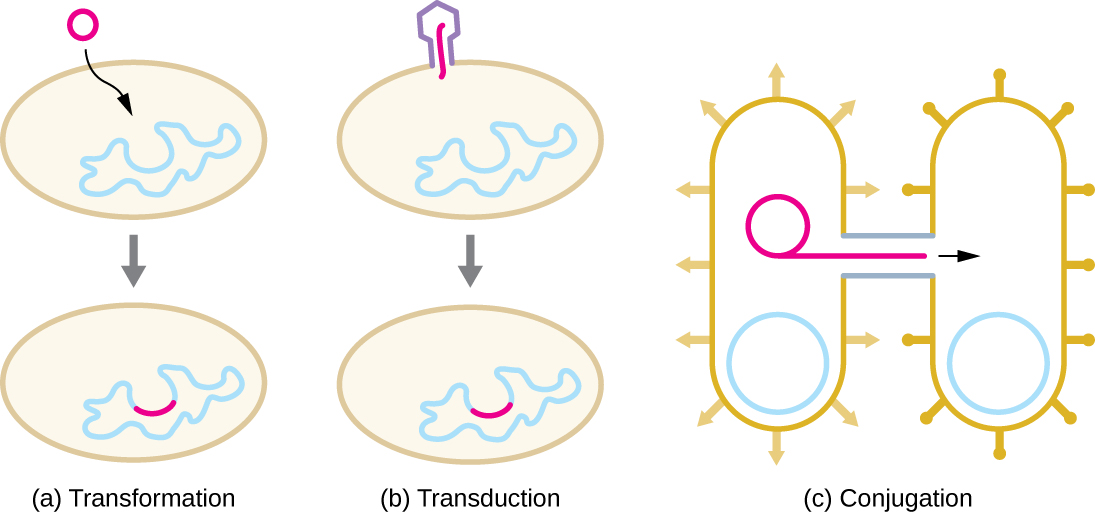
Sabe-se que o HGT em procariontes ocorre pelos três mecanismos principais ilustrados na Figura\(\PageIndex{1}\):
- Transformação: DNA nu é retirado do meio ambiente
- Transdução: genes são transferidos entre células em um vírus (veja O Ciclo de Vida Viral)
- Conjugação: uso de um tubo oco chamado pilus de conjugação para transferir genes entre células
Exercício\(\PageIndex{1}\)
- Quais são as três maneiras pelas quais a reprodução sexual introduz variação genética na prole?
- Qual é o benefício da reprodução assexuada?
- Quais são os três mecanismos de transferência horizontal de genes em procariontes?
Transformação
Frederick Griffith foi o primeiro a demonstrar o processo de transformação. Em 1928, ele mostrou que a bactéria Streptococcus pneumoniae viva e não patogênica poderia ser transformada em bactérias patogênicas por meio da exposição a uma cepa patogênica morta pelo calor. Ele concluiu que algum tipo de agente, que ele chamou de “princípio transformador”, havia sido passado da bactéria patogênica morta para a bactéria viva, não patogênica. Em 1944, Oswald Avery (1877—1955), Colin MacLeod (1909—1972) e Maclyn McCarty (1911—2005) demonstraram que o princípio transformador era o DNA (veja Usando a microbiologia para descobrir os segredos da vida).
Em transformação, o procarioto absorve o DNA nu encontrado em seu ambiente e que é derivado de outras células que se listaram ao morrer e liberaram seu conteúdo, incluindo seu genoma, no meio ambiente. Muitas bactérias são naturalmente competentes, o que significa que elas se ligam ativamente ao DNA ambiental, o transportam através de seus envelopes celulares até o citoplasma e o tornam de fita simples. Normalmente, o DNA estranho de fita dupla dentro das células é destruído pelas nucleases como defesa contra infecções virais. No entanto, essas nucleases geralmente são ineficazes contra o DNA de fita simples, então esse DNA de fita simples dentro da célula tem a oportunidade de se recombinar no genoma bacteriano. Uma molécula de DNA que contém fragmentos de DNA de diferentes organismos é chamada de DNA recombinante. (O DNA recombinante será discutido com mais detalhes em Micróbios e as ferramentas da engenharia genética.) Se a bactéria incorporar o novo DNA em seu próprio genoma por meio de recombinação, a célula bacteriana pode ganhar novas propriedades fenotípicas. Por exemplo, se uma bactéria não patogênica absorve o DNA de um gene de toxina de um patógeno e o incorpora em seu cromossomo, ela também pode se tornar patogênica. O DNA plasmídico também pode ser absorvido por bactérias competentes e conferir novas propriedades à célula. No geral, a transformação na natureza é um processo relativamente ineficiente porque os níveis de DNA ambiental são baixos devido à atividade das nucleases que também são liberadas durante a lise celular. Além disso, a recombinação genética é ineficiente na incorporação de novas sequências de DNA ao genoma.
Na natureza, a transformação bacteriana é um mecanismo importante para a aquisição de elementos genéticos que codificam fatores de virulência e resistência a antibióticos. Foi demonstrado que os genes que codificam a resistência a compostos antimicrobianos são difundidos na natureza, mesmo em ambientes não influenciados por humanos. Esses genes, que permitem que micróbios que vivem em comunidades mistas compitam por recursos limitados, podem ser transferidos dentro de uma população por transformação, bem como por outros processos do HGT. Em laboratório, podemos explorar o processo natural de transformação bacteriana para a engenharia genética para fazer uma grande variedade de medicamentos, conforme discutido em Micróbios e as Ferramentas da Engenharia Genética.
Exercício\(\PageIndex{2}\)
Por que uma célula bacteriana produz DNA ambiental trazido para a célula em uma forma de fita simples?
Transdução
Os vírus que infectam bactérias (bacteriófagos) também podem mover pequenos pedaços de DNA cromossômico de uma bactéria para outra em um processo chamado transdução (veja a Figura 6.2.3). Lembre-se de que, na transdução generalizada, qualquer pedaço de DNA cromossômico pode ser transferido para uma nova célula hospedeira por embalagem acidental de DNA cromossômico em uma cabeça de fago durante a montagem do fago. Por outro lado, a transdução especializada resulta da excisão imprecisa de um profago lisogênico do cromossomo bacteriano, de modo que ele carregue consigo um pedaço do cromossomo bacteriano de ambos os lados do local de integração do fago para uma nova célula hospedeira. Como resultado, o hospedeiro pode adquirir novas propriedades. Esse processo é chamado de conversão lisogênica. De importância médica, um fago lisogênico pode levar consigo um gene de virulência para seu novo hospedeiro. Uma vez inserido no cromossomo do novo hospedeiro, o novo hospedeiro pode ganhar patogenicidade. Várias bactérias patogênicas, incluindo Corynebacterium diphtheriae (o agente causador da difteria) e Clostridium botulinum (o agente causador do botulismo), são virulentas devido à introdução de genes codificadores de toxinas por bacteriófagos lisogênicos, afirmando a relevância da transdução na troca de genes envolvidos em doenças infecciosas. As arqueias têm seus próprios vírus que translocam material genético de um indivíduo para outro.
Exercício\(\PageIndex{3}\)
- Qual é o agente de transdução de células procarióticas?
- Na transdução especializada, de onde vem a parte transdutora do DNA?
As consequências clínicas da transdução
Paul, um trabalhador humanitário de 23 anos de Atlanta, viajou para o Haiti em 2011 para prestar ajuda após o terremoto de 2010. Depois de trabalhar lá por várias semanas, de repente, ele começou a sentir problemas abdominais, incluindo cólicas severas, náuseas, vômitos e diarreia aquosa. Ele também começou a sentir cãibras musculares intensas. Em uma clínica local, o médico suspeitou que os sintomas de Paul foram causados pela cólera porque houve um surto de cólera após o terremoto. Como a cólera é transmitida pela via fecal-oral, brechas na infraestrutura de saneamento, como geralmente ocorrem após desastres naturais, podem precipitar surtos. O médico confirmou o diagnóstico presuntivo usando um teste de colera. Ele então prescreveu a Paul uma dose única de doxiciclina, bem como sais de reidratação oral, instruindo-o a beber quantidades significativas de água limpa.
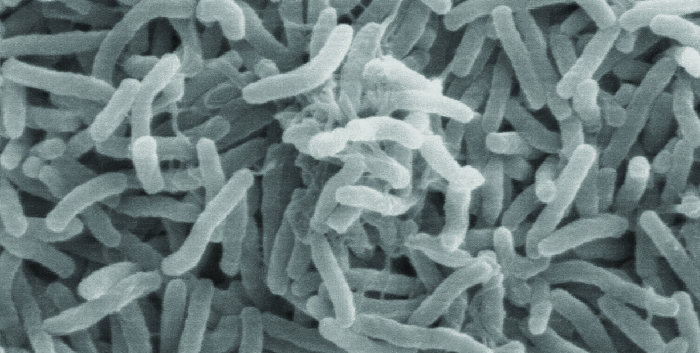
A cólera é causada pela haste curvada gram-negativa Vibrio cholerae (Figura\(\PageIndex{2}\)). Seus sintomas resultam em grande parte da produção da toxina da cólera (TC), que acaba ativando um transportador de cloreto para bombear íons cloreto das células epiteliais para o lúmen intestinal. A água então segue os íons cloreto, causando a prolífica diarreia aquosa característica da cólera. O gene que codifica a toxina da cólera é incorporado ao cromossomo bacteriano V. cóleras por meio da infecção da bactéria pelo fago filamentoso lisogênico CTX, que carrega o gene da tomografia computadorizada e o introduz no cromossomo na integração do profago. Assim, cepas patogênicas de V. cholerae resultam da transferência horizontal de genes por transdução especializada.
Exercício\(\PageIndex{4}\)
- Por que os surtos de cólera são mais comuns como resultado de um desastre natural?
- Por que as cãibras musculares são um sintoma comum da cólera? Por que o tratamento com sais de reidratação oral é tão importante para o tratamento da cólera?
- Em áreas atingidas pela cólera, quais são algumas estratégias que as pessoas poderiam usar para prevenir a transmissão de doenças?
Conjugação
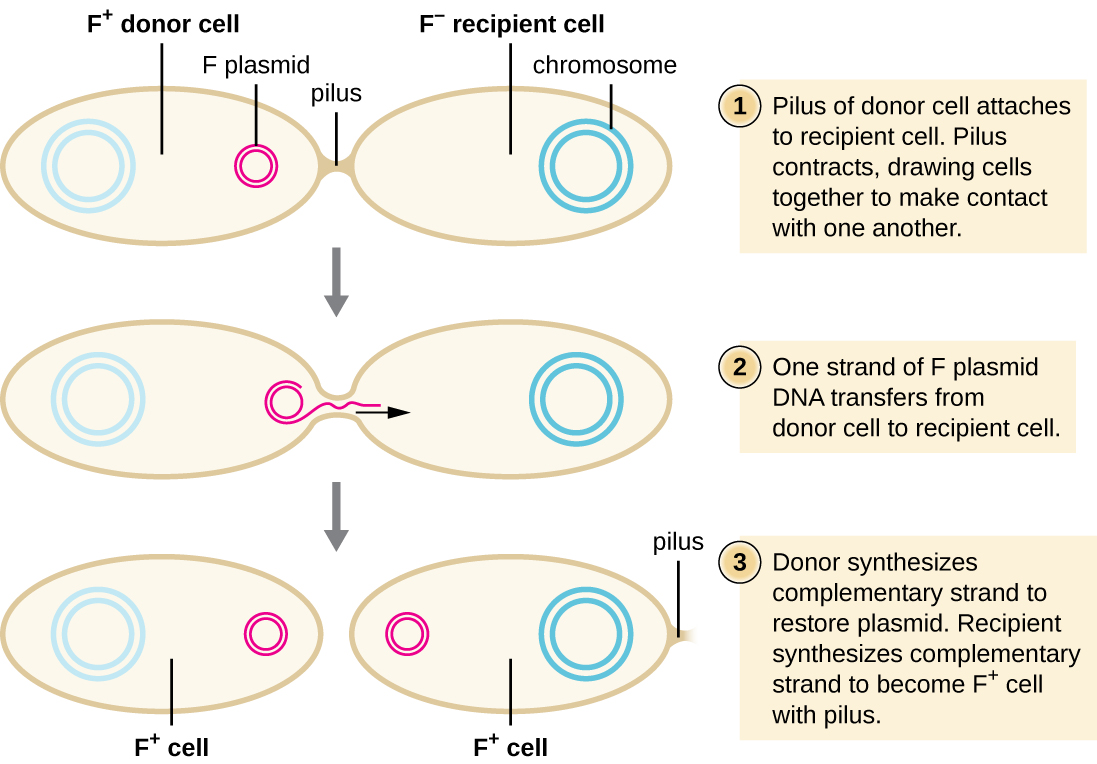
Na conjugação, o DNA é transferido diretamente de um procarioto para outro por meio de um pilus de conjugação, que coloca os organismos em contato uns com os outros. Em E. coli, os genes que codificam a capacidade de conjugação estão localizados em um plasmídeo bacteriano chamado plasmídeo F, também conhecido como fator de fertilidade, e o pilus de conjugação é chamado de F pilus. Os genes do plasmídeo F codificam tanto as proteínas que compõem o pilus F quanto as envolvidas na replicação circular do plasmídeo. As células que contêm o plasmídeo F, capazes de formar um pilus F, são chamadas de células F + ou células doadoras s, e aquelas sem plasmídeo F são chamadas de células F − ou células receptoras s.
Conjugação do plasmídeo F
Durante a conjugação típica em E. coli, o pilus F de uma célula F + entra em contato com uma célula F e se retrai, colocando os dois envelopes celulares em contato (Figura\(\PageIndex{3}\)). Em seguida, uma ponte citoplasmática se forma entre as duas células no local do pilus de conjugação. À medida que a replicação circular do plasmídeo F ocorre na célula F +, uma cópia de fita simples do plasmídeo F é transferida através da ponte citoplasmática para a célula F −, que então sintetiza a fita complementar, tornando-a de fita dupla. A célula F − agora se torna uma célula F + capaz de fazer seu próprio pilus de conjugação. Eventualmente, em uma população bacteriana mista contendo células F + e F −, todas as células se tornarão células F +. Os genes do plasmídeo F de E. coli também codificam proteínas que impedem a conjugação entre células F +.
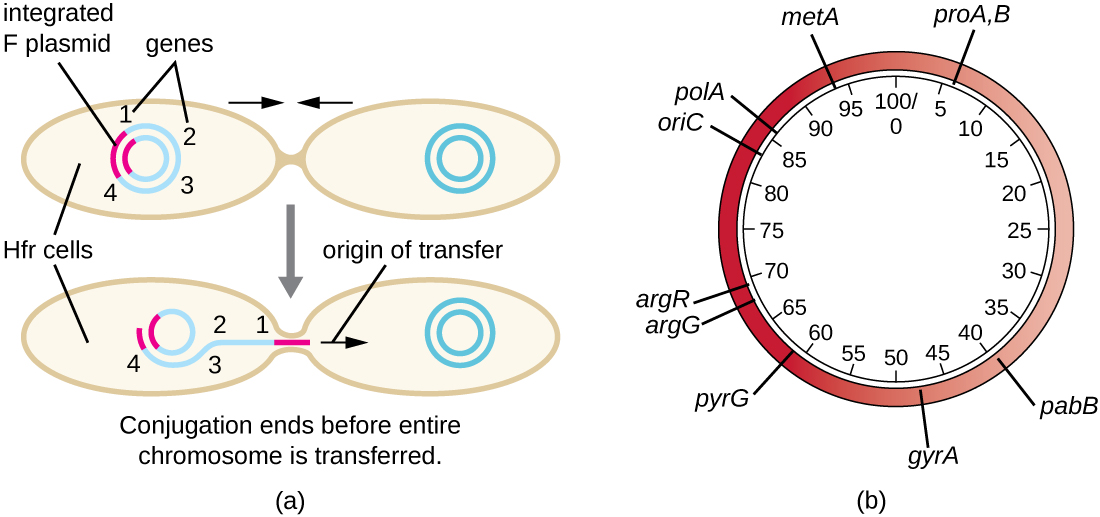
Conjugação de células F' e Hfr
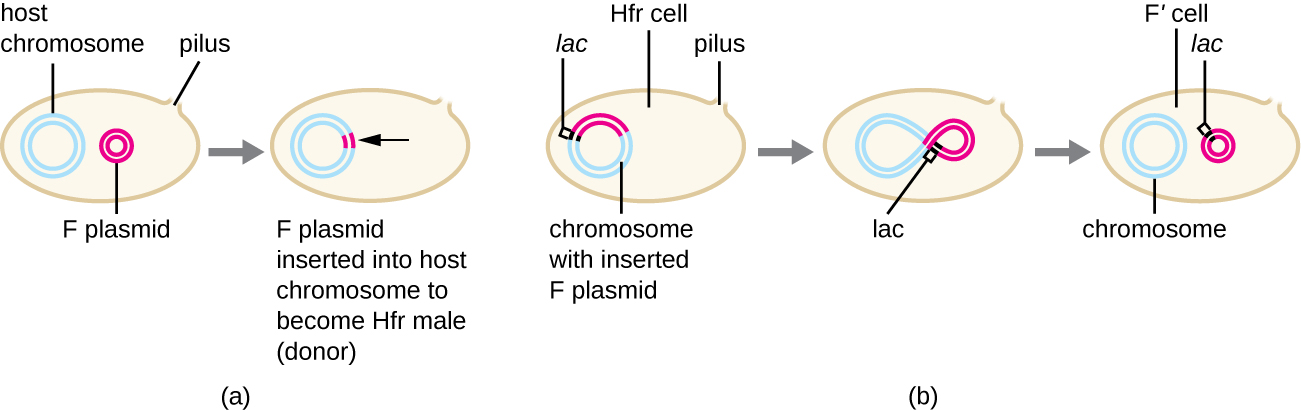
Embora a conjugação típica em E. coli resulte apenas na transferência do DNA do plasmídeo F, a conjugação também pode transferir DNA cromossômico. Isso ocorre porque o plasmídeo F ocasionalmente se integra ao cromossomo bacteriano por meio da recombinação entre o plasmídeo e o cromossomo, formando uma célula Hfr (Figura\(\PageIndex{4}\)). “Hfr” refere-se à alta frequência de recombinação observada quando as células F − receptoras recebem informações genéticas das células Hfr por meio de conjugação. Semelhante à excisão imprecisa de um profago durante a transdução especializada, o plasmídeo F integrado também pode ser excisado imprecisamente do cromossomo, produzindo um plasmídeo F' que carrega consigo algum DNA cromossômico adjacente ao local de integração. Na conjugação, esse DNA é introduzido na célula receptora e pode ser mantido como parte do plasmídeo F' ou ser recombinado no cromossomo bacteriano da célula receptora.
As células Hfr também podem tratar o cromossomo bacteriano como um enorme plasmídeo F e tentar transferir uma cópia dele para uma célula F − receptora. Como o cromossomo bacteriano é muito grande, a transferência de todo o cromossomo demora muito tempo (Figura\(\PageIndex{5}\)). No entanto, o contato entre as células bacterianas durante a conjugação é transitório, por isso é incomum que todo o cromossomo seja transferido. O DNA cromossômico hospedeiro próximo ao local de integração do plasmídeo F, deslocado pelo processo unidirecional de replicação circular, tem maior probabilidade de ser transferido e recombinado no cromossomo da célula receptora do que os genes hospedeiros mais distantes. Assim, a localização relativa dos genes bacterianos no genoma da célula Hfr pode ser mapeada com base em quando eles são transferidos por conjugação. Como resultado, antes da era do sequenciamento generalizado do genoma bacteriano, as distâncias nos mapas do genoma procariótico eram frequentemente medidas em minutos.
Consequências e aplicações da conjugação
Os plasmídeos são um importante tipo de elemento de DNA extracromossômico em bactérias e, nas células que os abrigam, são considerados parte do genoma bacteriano. Do ponto de vista clínico, os plasmídeos geralmente codificam genes envolvidos na virulência. Por exemplo, genes que codificam proteínas que tornam uma célula bacteriana resistente a um antibiótico específico são codificados em plasmídeos R. Os plasmídeos R, além de seus genes de resistência antimicrobiana, contêm genes que controlam a conjugação e a transferência do plasmídeo. Os plasmídeos R são capazes de se transferir entre células da mesma espécie e entre células de diferentes espécies. Os plasmídeos R únicos geralmente contêm vários genes que conferem resistência a vários antibióticos.
Genes necessários para a produção de várias toxinas e moléculas importantes para a colonização durante a infecção também podem ser encontrados codificados em plasmídeos. Por exemplo, cepas de E. coli produtoras de verotoxina (VTEC) parecem ter adquirido os genes que codificam a toxina Shiga de sua parente gram-negativa Shigella dysenteriae por meio da aquisição de um grande plasmídeo que codifica essa toxina. O VTEC causa doença diarreica grave que pode resultar na síndrome hemolítica urêmica (SHU), que pode levar à insuficiência renal e à morte.
Em ambientes não clínicos, genes bacterianos que codificam enzimas metabólicas necessárias para degradar compostos atípicos especializados, como hidrocarbonetos aromáticos policíclicos (PAHs), também são frequentemente codificados em plasmídeos. Além disso, certos plasmídeos têm a capacidade de se mover de células bacterianas para outros tipos de células, como as de plantas e animais, por meio de mecanismos distintos da conjugação. Esses mecanismos e seu uso na engenharia genética são abordados em Aplicações Modernas da Genética Microbiana.
Exercício\(\PageIndex{5}\)
- Que tipo de replicação ocorre durante a conjugação?
- O que ocorre para produzir uma célula Hfr E. coli?
- Quais tipos de características são codificadas nos plasmídeos?
Transposição
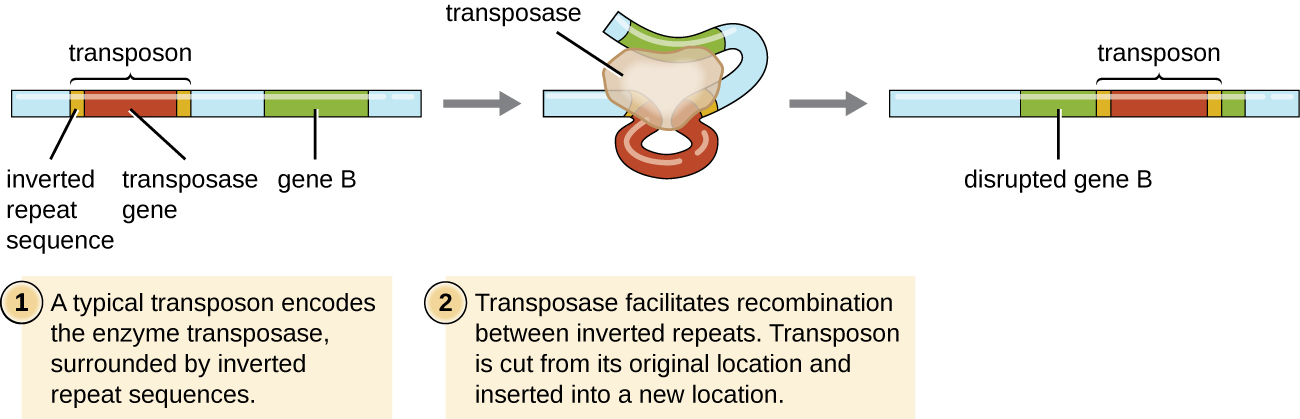
Elementos genéticos chamados transposons (elementos transponíveis), ou “genes saltadores”, são moléculas de DNA que incluem sequências repetidas invertidas especiais em suas extremidades e um gene que codifica a enzima transposase (Figura\(\PageIndex{6}\)). Os transposons permitem que toda a sequência seja extirpada de forma independente de um local em uma molécula de DNA e se integre ao DNA em outro lugar por meio de um processo chamado transposição. Os transposons foram originalmente descobertos no milho (milho) pela geneticista americana Barbara McClintock (1902—1992) na década de 1940. Desde então, os transposons foram encontrados em todos os tipos de organismos, tanto procariontes quanto eucariotos. Assim, diferentemente dos três mecanismos discutidos anteriormente, a transposição não é específica do procarioto. A maioria dos transposons não é replicativa, o que significa que eles se movem de forma “recortada e colada”. Alguns podem ser replicativos, no entanto, mantendo sua localização no DNA enquanto fazem uma cópia para ser inserida em outro lugar (“copiar e colar”). Como os transposons podem se mover dentro de uma molécula de DNA, de uma molécula de DNA para outra, ou mesmo de uma célula para outra, eles têm a capacidade de introduzir diversidade genética. O movimento dentro da mesma molécula de DNA pode alterar o fenótipo ao inativar ou ativar um gene.
Os transposons podem carregar consigo genes adicionais, movendo esses genes de um local para outro com eles. Por exemplo, os transposons bacterianos podem realocar genes de resistência a antibióticos, movendo-os dos cromossomos para os plasmídeos. Esse mecanismo demonstrou ser responsável pela colocalização de vários genes de resistência a antibióticos em um único plasmídeo R em cepas de Shigella que causam disenteria bacteriana. Esse plasmídeo R pode então ser facilmente transferido entre uma população bacteriana por meio do processo de conjugação.
Exercício\(\PageIndex{6}\)
Quais são as duas maneiras pelas quais um transposon pode afetar o fenótipo de uma célula para a qual ele se move?
| Prazo | Definição |
|---|---|
| Conjugação | Transferência de DNA por contato direto usando um pilus de conjugação |
| Transdução | Mecanismo de transferência horizontal de genes em bactérias nas quais os genes são transferidos por meio de infecção viral |
| Transformação | Mecanismo de transferência horizontal de genes em que o DNA ambiental nu é absorvido por uma célula bacteriana |
| Transposição | Processo pelo qual o DNA se retira de forma independente de um local em uma molécula de DNA e se integra em outro lugar |
Foco clínico: Parte 3
Apesar do tratamento contínuo com antibióticos, a infecção de Mark continuou a progredir rapidamente. A região infectada continuou a se expandir e ele teve que ser colocado em um ventilador para ajudá-lo a respirar. O médico de Mark ordenou a remoção cirúrgica do tecido infectado. Após uma cirurgia inicial, a ferida de Mark foi monitorada diariamente para garantir que a infecção não retornasse, mas continuasse a se espalhar.
Depois de duas rodadas adicionais de cirurgia, a infecção finalmente pareceu estar contida. Alguns dias depois, Mark foi retirado do ventilador e conseguiu respirar sozinho. No entanto, ele havia perdido uma grande quantidade de pele e tecidos moles na parte inferior da perna.
Exercício\(\PageIndex{7}\)
- Por que a remoção do tecido infectado interrompe a infecção?
- Quais são algumas complicações prováveis desse método de tratamento?
Conceitos principais e resumo
- A transferência horizontal de genes é uma forma importante para organismos que se reproduzem assexuadamente, como procariontes, adquirirem novas características.
- Existem três mecanismos de transferência horizontal de genes normalmente usados pelas bactérias: transformação, transdução e conjugação.
- A transformação permite que as células competentes absorvam o DNA nu, liberado de outras células quando morrem, para o citoplasma, onde ele pode se recombinar com o genoma do hospedeiro.
- Na transdução generalizada, qualquer pedaço de DNA cromossômico pode ser transferido pelo empacotamento acidental do cromossomo hospedeiro degradado em uma cabeça de fago. Na transdução especializada, somente o DNA cromossômico adjacente ao local de integração de um fago lisogênico pode ser transferido como resultado da excisão imprecisa do profago.
- A conjugação é mediada pelo plasmídeo F, que codifica um pilus de conjugação que coloca uma célula F + contendo plasmídeo F em contato com uma célula F.
- A rara integração do plasmídeo F no cromossomo bacteriano, gerando uma célula Hfr, permite a transferência do DNA cromossômico do doador para o receptor. Além disso, a excisão imprecisa do plasmídeo F do cromossomo pode gerar um plasmídeo F' que pode ser transferido para um receptor por conjugação.
- A transferência de conjugação de plasmídeos R é um mecanismo importante para a disseminação da resistência a antibióticos nas comunidades bacterianas.
- Transposons são moléculas de DNA com repetições invertidas em suas extremidades que também codificam a enzima transposase, permitindo sua movimentação de um local no DNA para outro. Embora encontrados em procariontes e eucariotos, os transposons são clinicamente relevantes em patógenos bacterianos para a movimentação de fatores de virulência, incluindo genes de resistência a antibióticos.