
8.2: Catabolismo de carboidratos
- Page ID
- 181487

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de
- Descreva por que a glicólise não depende de oxigênio
- Definir e descrever o rendimento líquido de moléculas de três carbonos, ATP e NADH da glicólise
- Explique como as moléculas de piruvato de três carbonos são convertidas em grupos acetil de dois carbonos que podem ser canalizados para o ciclo de Krebs.
- Defina e descreva o rendimento líquido de CO 2, GTP/ATP, FADH 2 e NADH do ciclo de Krebs
- Explique como as moléculas intermediárias de carbono do ciclo de Krebs podem ser usadas em uma célula
Existem extensas vias enzimáticas para decompor os carboidratos e capturar energia nas ligações ATP. Além disso, muitas vias catabólicas produzem moléculas intermediárias que também são usadas como blocos de construção para o anabolismo. Compreender esses processos é importante por vários motivos. Primeiro, como os principais processos metabólicos envolvidos são comuns a uma ampla variedade de organismos quimioheterotróficos, podemos aprender muito sobre o metabolismo humano estudando o metabolismo em bactérias mais facilmente manipuláveis, como a E. coli. Segundo, como os patógenos animais e humanos também são quimioheterotróficos, aprender sobre os detalhes do metabolismo dessas bactérias, incluindo possíveis diferenças entre as vias bacterianas e humanas, é útil para o diagnóstico de patógenos, bem como para a descoberta de terapias antimicrobianas direcionadas patógenos específicos. Por último, aprender especificamente sobre as vias envolvidas no metabolismo quimioheterotrófico também serve como base para comparar outras estratégias metabólicas mais incomuns usadas pelos micróbios. Embora a fonte química dos elétrons que iniciam a transferência de elétrons seja diferente entre quimioheterógrafos e quimioautotróficos, muitos processos similares são usados em ambos os tipos de organismos.
O exemplo típico usado para introduzir conceitos de metabolismo aos estudantes é o catabolismo de carboidratos. Para quimioheterotróficos, nossos exemplos de metabolismo começam com o catabolismo de polissacarídeos, como glicogênio, amido ou celulose. Enzimas como a amilase, que decompõe o glicogênio ou o amido, e as celulases, que decompõem a celulose, podem causar a hidrólise das ligações glicosídicas entre os monômeros de glicose nesses polímeros, liberando glicose para um maior catabolismo.
Glicólise
Para bactérias, eucariotos e a maioria das arquéias, a glicólise é a via mais comum para o catabolismo da glicose; ela produz energia, portadores de elétrons reduzidos e moléculas precursoras para o metabolismo celular. Todo organismo vivo realiza alguma forma de glicólise, sugerindo que esse mecanismo é um antigo processo metabólico universal. O processo em si não usa oxigênio; no entanto, a glicólise pode ser associada a processos metabólicos adicionais que são aeróbicos ou anaeróbicos. A glicólise ocorre no citoplasma das células procarióticas e eucarióticas. Começa com uma única molécula de glicose de seis carbonos e termina com duas moléculas de um açúcar de três carbonos chamado piruvato. O piruvato pode ser decomposto ainda mais após a glicólise para aproveitar mais energia por meio da respiração aeróbica ou anaeróbica, mas muitos organismos, incluindo muitos micróbios, podem ser incapazes de respirar; para esses organismos, a glicólise pode ser sua única fonte de geração de ATP.
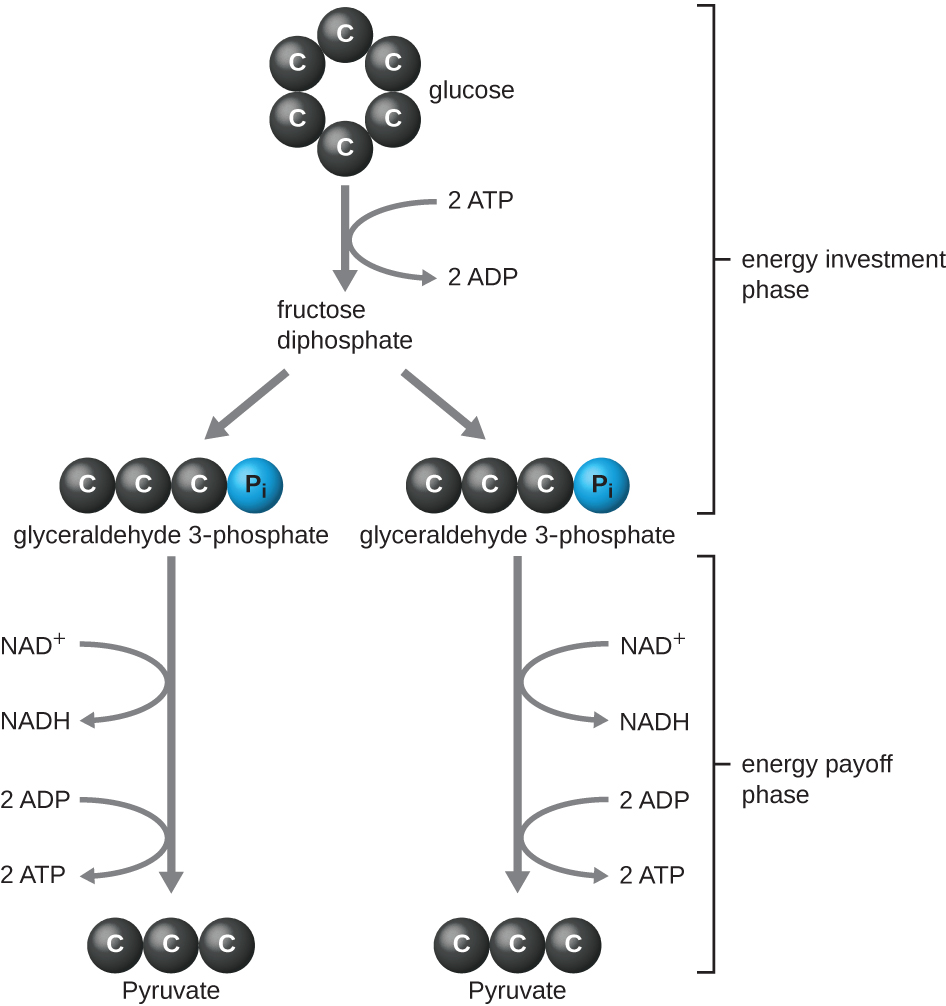
O tipo de glicólise encontrado em animais e que é mais comum em micróbios é a via Embden-Meyerhof-Parnas (EMP), em homenagem a Gustav Embden (1874-1933), Otto Meyerhof (1884-1951) e Jakub Parnas (1884-1949). A glicólise usando a via EMP consiste em duas fases distintas (Figura\(\PageIndex{1}\)). A primeira parte do caminho, chamada de fase de investimento em energia, usa energia de duas moléculas de ATP para modificar uma molécula de glicose para que a molécula de açúcar de seis carbonos possa ser dividida uniformemente em duas moléculas fosforiladas de três carbonos chamadas gliceraldeído 3-fosfato (G3P). A segunda parte da via, chamada de fase de pagamento de energia, extrai energia oxidando o G3P em piruvato, produzindo quatro moléculas de ATP e reduzindo duas moléculas de NAD+ a duas moléculas de NADH, usando elétrons que se originaram da glicose. (Uma discussão e ilustração da via EMP completa com estruturas químicas e nomes de enzimas aparecem no Apêndice C.)
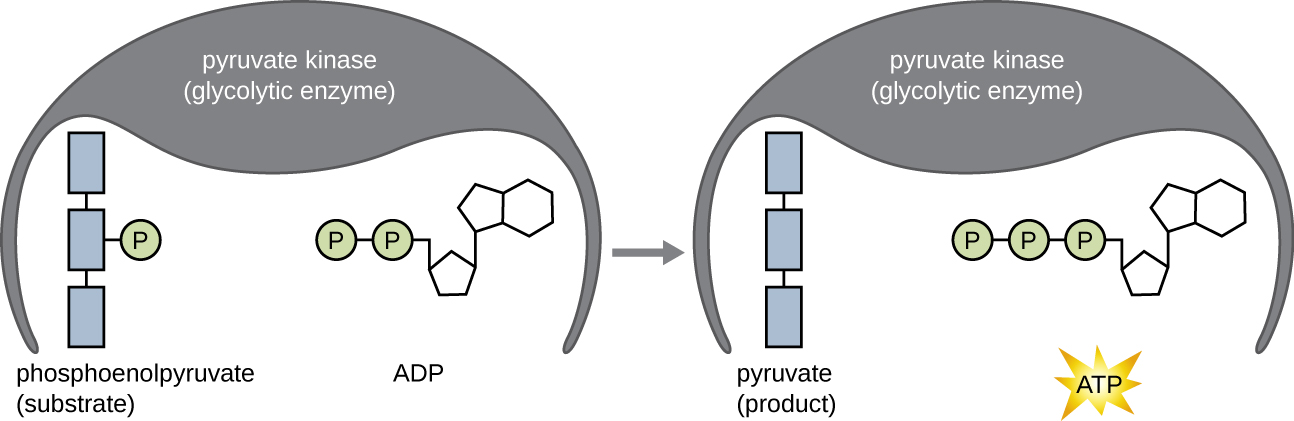
As moléculas de ATP produzidas durante a fase de retorno de energia da glicólise são formadas pela fosforilação em nível de substrato (Figura\(\PageIndex{1}\)), um dos dois mecanismos de produção de ATP. Na fosforilação em nível de substrato, um grupo fosfato é removido de uma molécula orgânica e transferido diretamente para uma molécula de ADP disponível, produzindo ATP. Durante a glicólise, grupos de fosfato de alta energia das moléculas intermediárias são adicionados ao ADP para produzir ATP.
No geral, nesse processo de glicólise, o ganho líquido da quebra de uma única molécula de glicose é:
- duas moléculas de ATP
- duas moléculas de NADH e
- duas moléculas de piruvato.
Outras vias glicolíticas
Quando nos referimos à glicólise, salvo indicação em contrário, estamos nos referindo à via EMP usada por animais e muitas bactérias. No entanto, alguns procariontes usam vias glicolíticas alternativas. Uma alternativa importante é a via Entner-Doudoroff (ED), em homenagem a seus descobridores Nathan Entner e Michael Doudoroff (1911—1975). Embora algumas bactérias, incluindo o patógeno oportunista gram-negativo Pseudomonas aeruginosa, contenham apenas a via ED para a glicólise, outras bactérias, como a E. coli, têm a capacidade de usar a via ED ou a via EMP.
Um terceiro tipo de via glicolítica que ocorre em todas as células, que é bem diferente das duas vias anteriores, é a via da pentose fosfato (PPP), também chamada de via do fosfogluconato ou derivação da hexose monofosfato. As evidências sugerem que o PPP pode ser a via glicolítica universal mais antiga. Os intermediários do PPP são usados para a biossíntese de nucleotídeos e aminoácidos. Portanto, essa via glicolítica pode ser favorecida quando a célula precisa de síntese de ácido nucléico e/ou proteína, respectivamente. Uma discussão e ilustração da via de ED completa e do PPP com estruturas químicas e nomes de enzimas aparecem no Apêndice C.
Exercício\(\PageIndex{1}\)
Quando um organismo pode usar a via de ED ou o PPP para glicólise?
Reação de transição, coenzima A e o ciclo de Krebs
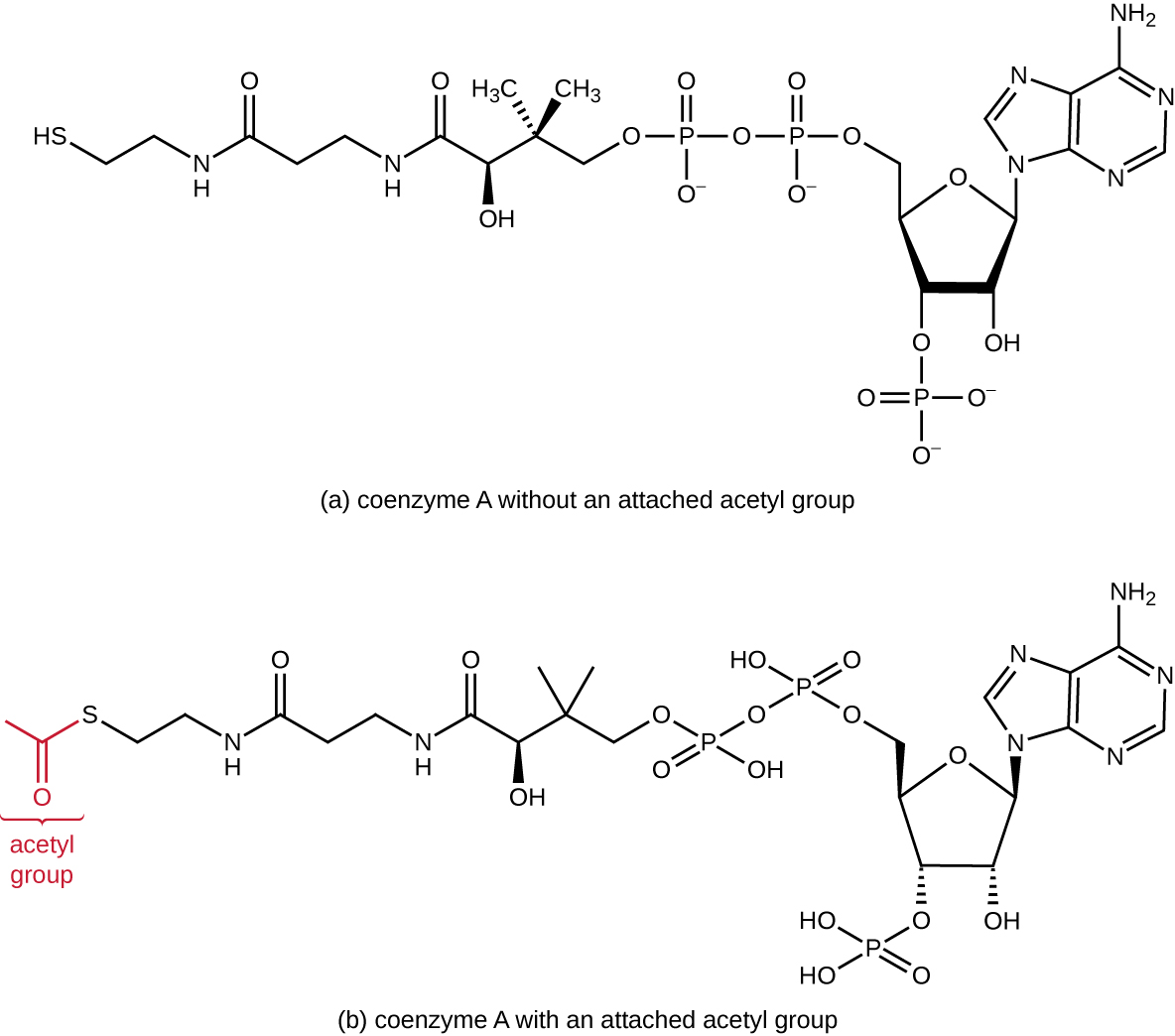
A glicólise produz piruvato, que pode ser ainda mais oxidado para capturar mais energia. Para que o piruvato entre na próxima via oxidativa, ele deve primeiro ser descarboxilado pelo complexo enzimático piruvato desidrogenase em um grupo acetil de dois carbonos na reação de transição, também chamada de reação ponte (ver Apêndice C e Figura\(\PageIndex{3}\)). Na reação de transição, os elétrons também são transferidos para o NAD + para formar o NADH. Para prosseguir para a próxima fase desse processo metabólico, o relativamente pequeno acetil de dois carbonos deve ser ligado a um composto transportador muito grande chamado coenzima A (CoA). A reação de transição ocorre na matriz mitocondrial dos eucariotos; nos procariontes, ocorre no citoplasma porque os procariontes não possuem organelas fechadas pela membrana.
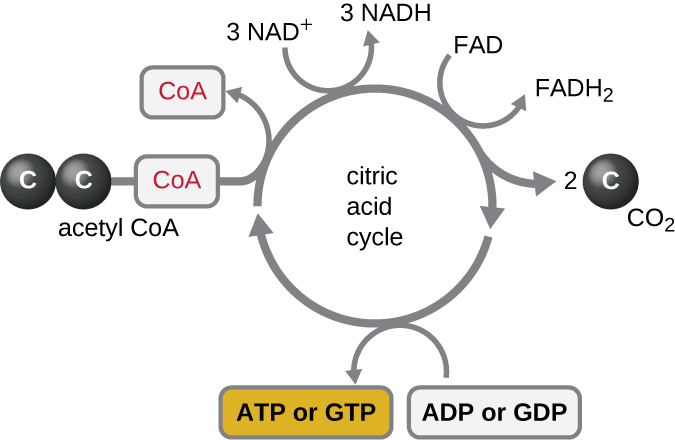
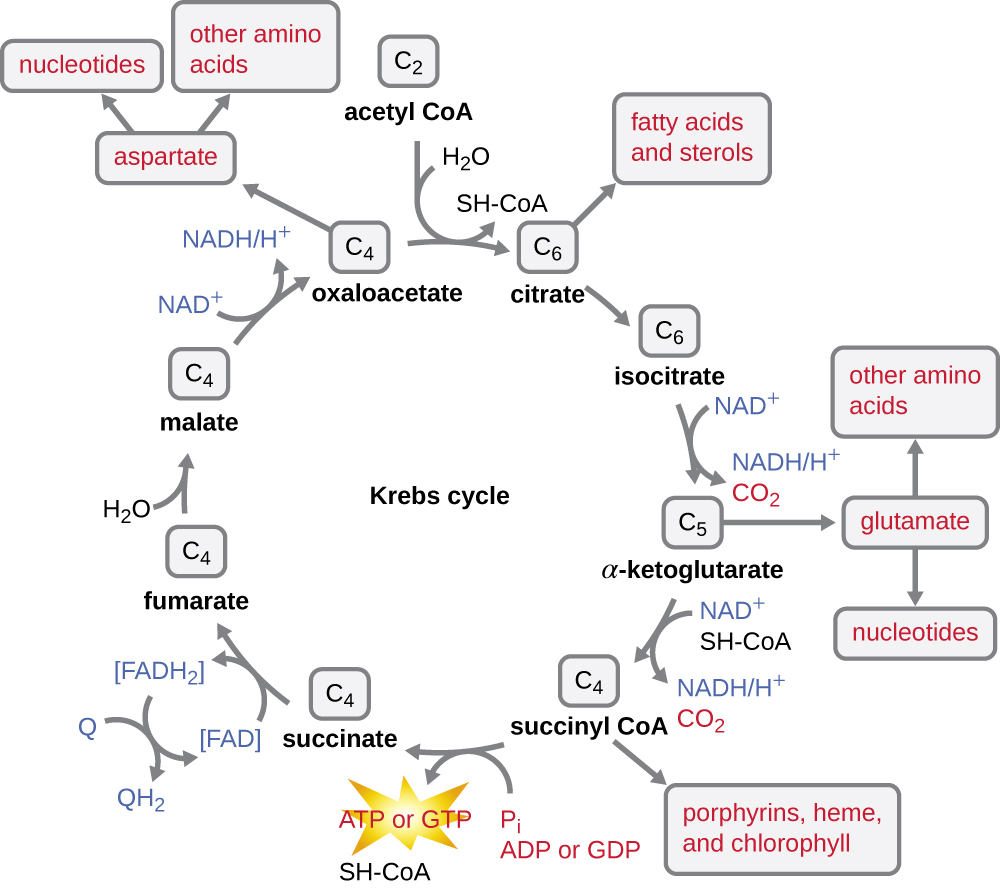
O ciclo de Krebs transfere os elétrons restantes do grupo acetil produzido durante a reação de transição para as moléculas transportadoras de elétrons, reduzindo-os assim. O ciclo de Krebs também ocorre no citoplasma dos procariontes junto com a glicólise e a reação de transição, mas ocorre na matriz mitocondrial das células eucarióticas, onde a reação de transição também ocorre. O ciclo de Krebs recebeu o nome de seu descobridor, o cientista britânico Hans Adolf Krebs (1900-1981) e também é chamado de ciclo do ácido cítrico, ou ciclo do ácido tricarboxílico (TCA) porque o ácido cítrico tem três grupos carboxila em sua estrutura. Ao contrário da glicólise, o ciclo de Krebs é um ciclo fechado: a última parte da via regenera o composto usado na primeira etapa (Figura\(\PageIndex{4}\)). As oito etapas do ciclo são uma série de reações químicas que capturam o grupo acetil de dois carbonos (o transportador de CoA não entra no ciclo de Krebs) da reação de transição, que é adicionada a um intermediário de quatro carbonos no ciclo de Krebs, produzindo o ácido cítrico intermediário de seis carbonos (dando nome alternativo para este ciclo). Quando uma volta do ciclo retorna ao ponto inicial do intermediário de quatro carbonos, o ciclo produz duas moléculas de CO 2, uma molécula de ATP (ou equivalente, como trifosfato de guanosina [GTP]) produzida por fosforilação em nível de substrato e três moléculas de NADH e uma de FADH 2 . (Uma discussão e uma ilustração detalhada do ciclo completo de Krebs aparecem no Apêndice C.)
Embora muitos organismos usem o ciclo de Krebs conforme descrito como parte do metabolismo da glicose, vários dos compostos intermediários do ciclo de Krebs podem ser usados na síntese de uma ampla variedade de moléculas celulares importantes, incluindo aminoácidos, clorofilas, ácidos graxos e nucleotídeos; portanto, o ciclo é tanto anabólico quanto catabólico (Figura\(\PageIndex{5}\)).
Conceitos principais e resumo
- A glicólise é o primeiro passo na quebra da glicose, resultando na formação de ATP, que é produzido pela fosforilação em nível de substrato; NADH; e duas moléculas de piruvato. A glicólise não usa oxigênio e não é dependente de oxigênio.
- Após a glicólise, um piruvato de três carbonos é descarboxilado para formar um grupo acetil de dois carbonos, juntamente com a formação de NADH. O grupo acetil está ligado a um grande composto transportador chamado coenzima A.
- Após a etapa de transição, a coenzima A transporta o acetil de dois carbonos para o ciclo de Krebs, onde os dois carbonos entram no ciclo. Por turno do ciclo, um grupo acetil derivado da glicólise é ainda mais oxidado, produzindo três moléculas de NADH, uma FADH 2 e um ATP por fosforilação em nível de substrato e liberando duas moléculas de CO 2.
- O ciclo de Krebs pode ser usado para outros fins. Muitos dos intermediários são usados para sintetizar moléculas celulares importantes, incluindo aminoácidos, clorofilas, ácidos graxos e nucleotídeos.