
3.3: Características únicas das células procarióticas
- Page ID
- 181816

Objetivos de
- Explicar as características distintivas das células procarióticas
- Descreva morfologias celulares comuns e arranjos celulares típicos das células procarióticas e explique como as células mantêm sua morfologia
- Descreva as estruturas internas e externas das células procarióticas em termos de sua estrutura física, estrutura química e função
- Compare as características distintivas das células bacterianas e arqueais
A teoria celular afirma que a célula é a unidade fundamental da vida. No entanto, as células variam significativamente em tamanho, forma, estrutura e função. No nível mais simples de construção, todas as células possuem alguns componentes fundamentais. Isso inclui citoplasma (uma substância gelatinosa composta por água e produtos químicos dissolvidos necessários para o crescimento), que está contido em uma membrana plasmática (também chamada de membrana celular ou membrana citoplasmática); um ou mais cromossomos, que contêm os projetos genéticos da célula; e ribossomos, organelas usadas para a produção de proteínas.
Além desses componentes básicos, as células podem variar muito entre os organismos e até mesmo dentro do mesmo organismo multicelular. As duas maiores categorias de células - células procarióticas e células eucarióticas - são definidas por grandes diferenças em várias estruturas celulares. As células procarióticas não possuem um núcleo cercado por uma membrana nuclear complexa e geralmente têm um único cromossomo circular localizado em um nucleóide. As células eucarióticas têm um núcleo cercado por uma membrana nuclear complexa que contém vários cromossomos em forma de bastonete. 1
Todas as células vegetais e animais são eucarióticas. Alguns microrganismos são compostos por células procarióticas, enquanto outros são compostos por células eucarióticas. Os microrganismos procarióticos são classificados nos domínios Archaea e Bactérias, enquanto os eucarióticos são classificados no domínio Eukarya.
As estruturas dentro de uma célula são análogas aos órgãos dentro do corpo humano, com estruturas únicas adequadas a funções específicas. Algumas das estruturas encontradas nas células procarióticas são semelhantes às encontradas em algumas células eucarióticas; outras são exclusivas dos procariontes. Embora haja algumas exceções, as células eucarióticas tendem a ser maiores do que as células procarióticas. O tamanho comparativamente maior das células eucarióticas determina a necessidade de compartimentar vários processos químicos em diferentes áreas da célula, usando organelas complexas ligadas à membrana. Em contraste, as células procarióticas geralmente não possuem organelas ligadas à membrana; no entanto, elas geralmente contêm inclusões que compartimentam seu citoplasma. A figura\(\PageIndex{1}\) ilustra estruturas tipicamente associadas a células procarióticas. Essas estruturas são descritas com mais detalhes na próxima seção.
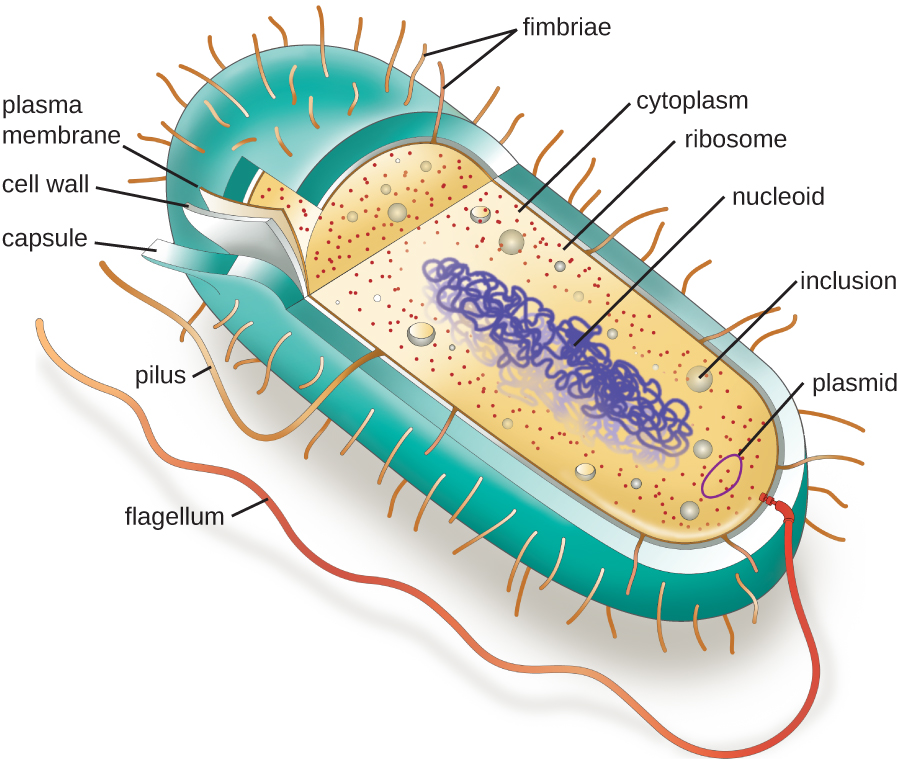
Morfologias e arranjos celulares comuns
Células individuais de um organismo procariótico específico são normalmente semelhantes em forma ou morfologia celular. Embora milhares de organismos procarióticos tenham sido identificados, apenas algumas morfologias celulares são comumente vistas microscopicamente. A figura\(\PageIndex{2}\) nomeia e ilustra as morfologias celulares comumente encontradas em células procarióticas. Além da forma celular, células procarióticas da mesma espécie podem se agrupar em certos arranjos distintos, dependendo do plano de divisão celular. Alguns arranjos comuns são mostrados na Figura\(\PageIndex{3}\).
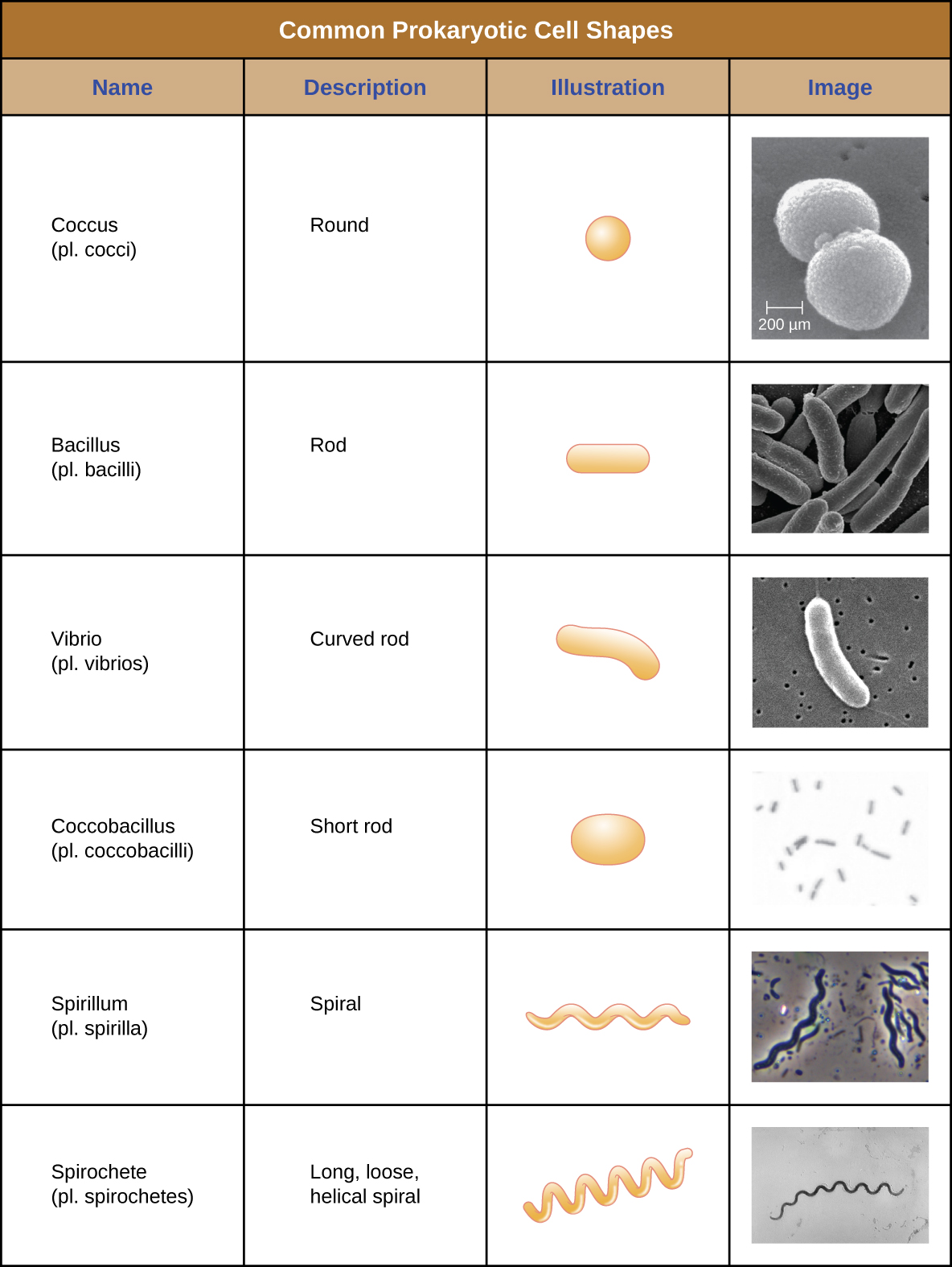
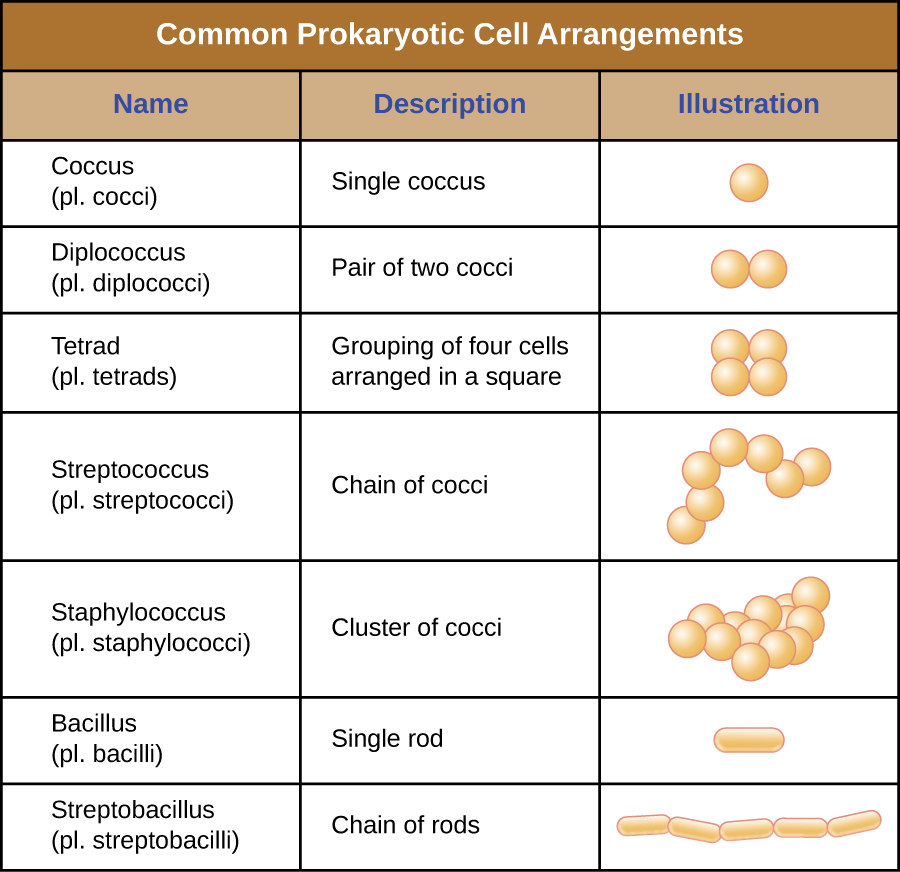
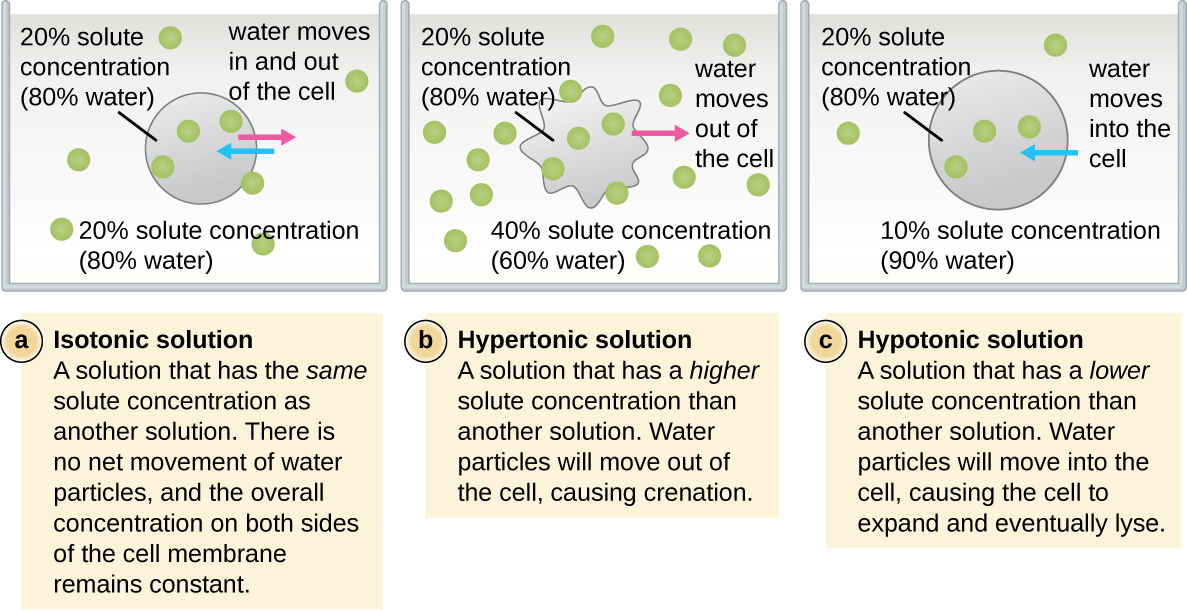
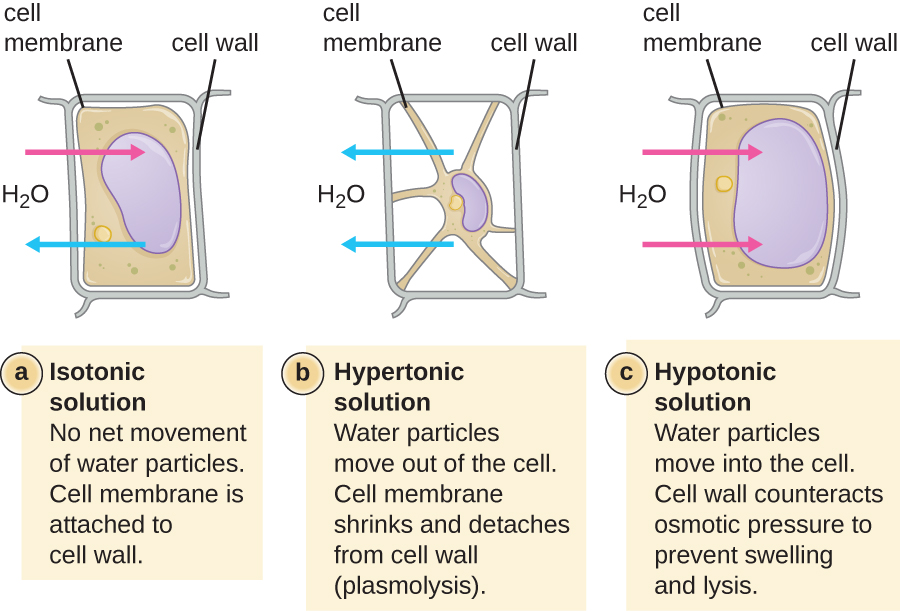
Na maioria das células procarióticas, a morfologia é mantida pela parede celular em combinação com elementos do citoesqueleto. A parede celular é uma estrutura encontrada na maioria dos procariontes e em alguns eucariotos; ela envolve a membrana celular, protegendo a célula das mudanças na pressão osmótica (Figura\(\PageIndex{4}\)). A pressão osmótica ocorre devido a diferenças na concentração de solutos em lados opostos de uma membrana semipermeável. A água é capaz de passar por uma membrana semipermeável, mas os solutos (moléculas dissolvidas como sais, açúcares e outros compostos) não conseguem. Quando a concentração de solutos é maior em um lado da membrana, a água se difunde pela membrana do lado com a menor concentração (mais água) para o lado com a maior concentração (menos água) até que as concentrações em ambos os lados se tornem iguais. Essa difusão da água é chamada de osmose e pode causar pressão osmótica extrema em uma célula quando seu ambiente externo muda.
O ambiente externo de uma célula pode ser descrito como um meio isotônico, hipertônico ou hipotônico. Em um meio isotônico, as concentrações de soluto dentro e fora da célula são aproximadamente iguais, portanto, não há movimento líquido de água pela membrana celular. Em um meio hipertônico, a concentração de soluto fora da célula excede aquela dentro da célula, então a água se difunde para fora da célula e para o meio externo. Em um meio hipotônico, a concentração de soluto dentro da célula excede aquela fora da célula, então a água se moverá por osmose para dentro da célula. Isso faz com que a célula inche e potencialmente se libere ou rompa.
O grau em que uma determinada célula é capaz de suportar mudanças na pressão osmótica é chamado de tonicidade. As células que têm uma parede celular são mais capazes de suportar mudanças sutis na pressão osmótica e manter sua forma. Em ambientes hipertônicos, as células que não possuem parede celular podem ficar desidratadas, causando creação ou enrugamento da célula; a membrana plasmática se contrai e parece recortada ou entalhada (Figura\(\PageIndex{4}\)). Por outro lado, as células que possuem uma parede celular sofrem plasmólise em vez de crenação. Na plasmólise, a membrana plasmática se contrai e se desprende da parede celular, e há uma diminuição no volume interno, mas a parede celular permanece intacta, permitindo que a célula mantenha alguma forma e integridade por um período de tempo (Figura\(\PageIndex{5}\)). Da mesma forma, células que não possuem parede celular são mais propensas à lise em ambientes hipotônicos. A presença de uma parede celular permite que a célula mantenha sua forma e integridade por mais tempo antes da lisagem (Figura\(\PageIndex{5}\)).
Exercício\(\PageIndex{1}\)
- Explique a diferença entre a morfologia e o arranjo celular.
- Quais as vantagens que as paredes celulares oferecem às células procarióticas?
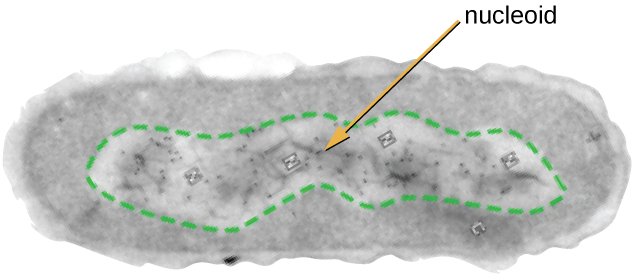
O Nucleóide
Toda a vida celular tem um genoma de DNA organizado em um ou mais cromossomos. Os cromossomos procarióticos são tipicamente circulares, haplóides (não pareados) e não estão ligados por uma membrana nuclear complexa. O DNA procariótico e as proteínas associadas ao DNA estão concentrados na região nucleóide da célula (Figura\(\PageIndex{6}\)). Em geral, o DNA procariótico interage com proteínas associadas a nucleóides (NAPs) que auxiliam na organização e empacotamento do cromossomo. Nas bactérias, os NAPs funcionam de forma semelhante às histonas, que são as proteínas organizadoras do DNA encontradas nas células eucarióticas. Nas arquéias, o nucleóide é organizado por NAPs ou proteínas organizadoras de DNA semelhantes a histonas.
Plasmídeos
As células procarióticas também podem conter DNA extracromossômico ou DNA que não faz parte do cromossomo. Esse DNA extracromossômico é encontrado em plasmídeos, que são moléculas de DNA pequenas, circulares e de fita dupla. Células que têm plasmídeos geralmente têm centenas delas dentro de uma única célula. Os plasmídeos são mais comumente encontrados em bactérias; no entanto, plasmídeos foram encontrados em arquéias e organismos eucarióticos. Os plasmídeos geralmente carregam genes que conferem características vantajosas, como resistência a antibióticos; portanto, eles são importantes para a sobrevivência do organismo. Discutiremos os plasmídeos com mais detalhes em Mecanismos de Genética Microbiana.
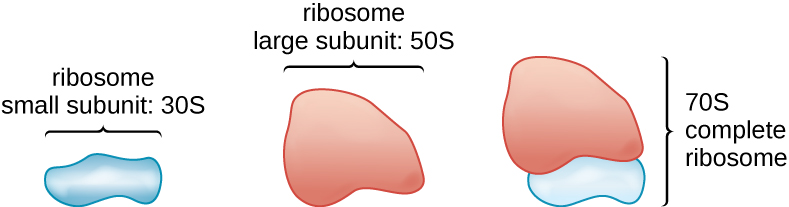
Ribossomos
Toda a vida celular sintetiza proteínas, e organismos em todos os três domínios da vida possuem ribossomos, estruturas responsáveis pela síntese protéica. No entanto, os ribossomos em cada um dos três domínios são estruturalmente diferentes. Os próprios ribossomos são construídos a partir de proteínas, junto com o RNA ribossômico (rRNA). Os ribossomos procarióticos são encontrados no citoplasma. Eles são chamados de ribossomos 70S porque têm um tamanho de 70S (Figura\(\PageIndex{7}\)), enquanto os ribossomos citoplasmáticos eucarióticos têm um tamanho de 80S. (O S significa unidade Svedberg, uma medida de sedimentação em uma ultracentrífuga, que é baseada no tamanho, forma e qualidades de superfície da estrutura que está sendo analisada). Embora tenham o mesmo tamanho, os ribossomos bacterianos e arqueais têm proteínas e moléculas de rRNA diferentes, e as versões arqueais são mais semelhantes às eucarióticas do que às encontradas nas bactérias.
Inclusões
Como organismos unicelulares vivem em ambientes instáveis, algumas células procarióticas têm a capacidade de armazenar nutrientes em excesso nas estruturas citoplasmáticas chamadas inclusões. Armazenar nutrientes em uma forma polimerizada é vantajoso porque reduz o acúmulo de pressão osmótica que ocorre quando uma célula acumula solutos. Vários tipos de inclusões armazenam glicogênio e amidos, que contêm carbono que as células podem acessar para obter energia. Os grânulos de volutina, também chamados de grânulos metacromáticos por suas características de coloração, são inclusões que armazenam fosfato inorgânico polimerizado que podem ser usados no metabolismo e auxiliar na formação de biofilmes. Os micróbios conhecidos por conter grânulos de volutina incluem a archaea Methanosarcina, a bactéria Corynebacterium diphtheriae e a alga eucariótica unicelular Chlamydomonas. Grânulos de enxofre, outro tipo de inclusão, são encontrados em bactérias de enxofre do gênero Thiobacillus; esses grânulos armazenam enxofre elementar, que as bactérias usam para o metabolismo.
Ocasionalmente, certos tipos de inclusões são cercados por uma monocamada fosfolipídica incorporada com proteína. O polihidroxibutirato (PHB), que pode ser produzido por espécies de Bacillus e Pseudomonas, é um exemplo de inclusão que apresenta esse tipo de estrutura monocamada. Industrialmente, o PHB também tem sido usado como fonte de polímeros biodegradáveis para bioplásticos. Vários tipos diferentes de inclusões são mostrados na Figura\(\PageIndex{8}\).

Algumas células procarióticas têm outros tipos de inclusões que servem a outros propósitos além do armazenamento de nutrientes. Por exemplo, algumas células procarióticas produzem vacúolos gasosos, acumulações de pequenas vesículas de gás revestidas de proteínas. Esses vacúolos gasosos permitem que as células procarióticas que os sintetizam alterem sua flutuabilidade para que possam ajustar sua localização na coluna de água. Bactérias magnetotáticas, como Magnetospirillum magnetotacticum, contêm magnetossomos, que são inclusões de óxido de ferro magnético ou sulfeto de ferro cercadas por uma camada lipídica. Isso permite que as células se alinhem ao longo de um campo magnético, auxiliando em seu movimento (Figura\(\PageIndex{8}\)). Cianobactérias como Anabaena cylindrica e bactérias como Halothiobacillus neapolitanus produzem inclusões de carboxisomas. Os carboxisomas são compostos por camadas externas de milhares de subunidades proteicas. Seu interior é preenchido com ribulose-1,5-bisfosfato carboxilase/oxigenase (RuBisCO) e anidrase carbônica. Ambos os compostos são usados para o metabolismo do carbono. Algumas células procarióticas também possuem carboxisomas que sequestram enzimas funcionalmente relacionadas em um único local. Essas estruturas são consideradas proto-organelas porque compartimentam compostos ou reações químicas importantes, assim como muitas organelas eucarióticas.
Endósporos
As células bacterianas são geralmente observadas como células vegetativas, mas alguns gêneros de bactérias têm a capacidade de formar endósporos, estruturas que essencialmente protegem o genoma bacteriano em um estado latente quando as condições ambientais são desfavoráveis. Os endósporos (não devem ser confundidos com os esporos reprodutivos formados por fungos) permitem que algumas células bacterianas sobrevivam por longos períodos sem comida ou água, bem como exposição a produtos químicos, temperaturas extremas e até radiação. A tabela\(\PageIndex{1}\) compara as características das células vegetativas e dos endosporos.
| Células vegetativas | Endósporos |
|---|---|
| Sensível a temperaturas e radiações extremas | Resistente a temperaturas e radiações extremas |
| Gram-positivo | Não absorva a coloração de Gram, apenas manchas especiais de endosporos (consulte Coloração de amostras microscópicas) |
| Conteúdo normal de água e atividade enzimática | Desidratado; sem atividade metabólica |
| Capaz de crescimento e metabolismo ativos | Dormente; sem crescimento ou atividade metabólica |
O processo pelo qual as células vegetativas se transformam em endosporos é chamado de esporulação e geralmente começa quando os nutrientes se esgotam ou as condições ambientais se tornam desfavoráveis (Figura\(\PageIndex{9}\)). O processo começa com a formação de um septo na célula bacteriana vegetativa. O septo divide a célula de forma assimétrica, separando um anteesporo de DNA da célula-mãe. O prepúcio, que formará o núcleo do endosporo, é essencialmente uma cópia dos cromossomos da célula e é separado da célula-mãe por uma segunda membrana. Um córtex se forma gradualmente ao redor do anteporo, depositando camadas de cálcio e ácido dipicolínico entre as membranas. Uma camada de esporos de proteína então se forma ao redor do córtex enquanto o DNA da célula-mãe se desintegra. A maturação adicional do endosporo ocorre com a formação de um exosporio externo. O endosporo é liberado após a desintegração da célula-mãe, completando a esporulação.
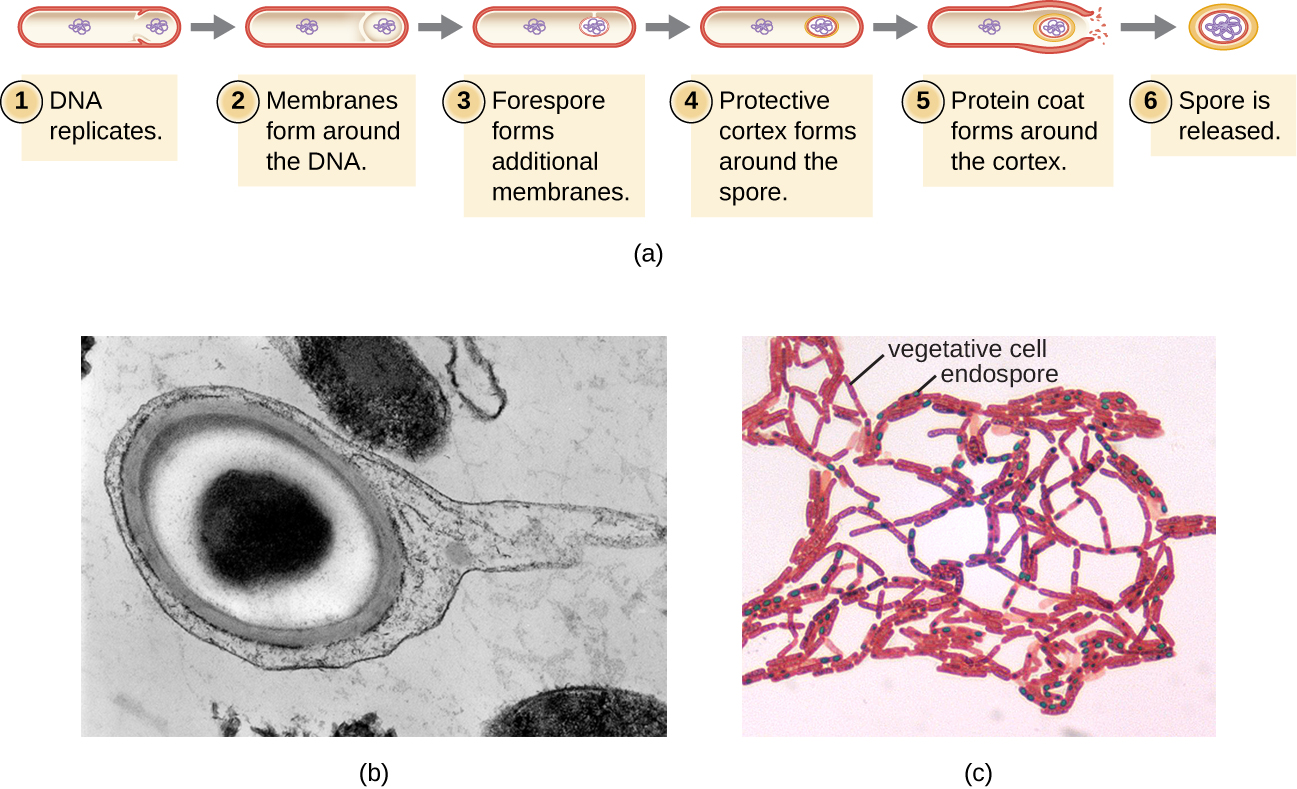
Foi demonstrado que os endósporos de certas espécies persistem em estado de dormência por longos períodos de tempo, até milhares de anos. 2 No entanto, quando as condições de vida melhoram, os endosporos germinam, reentrando no estado vegetativo. Após a germinação, a célula se torna metabolicamente ativa novamente e é capaz de realizar todas as suas funções normais, incluindo crescimento e divisão celular.
Nem todas as bactérias têm a capacidade de formar endosporos; no entanto, existem várias bactérias gram-positivas formadoras de endosporos clinicamente significativas dos gêneros Bacillus e Clostridium. Isso inclui B. anthracis, o agente causador do antraz, que produz endosporos capazes de sobreviver por muitas décadas 3; C. tetani (causa tétano); C. difficile (causa colite pseudomembranosa); C. perfringens (causa gangrena gasosa); e C. botulinum (causa botulismo). Patógenos como esses são particularmente difíceis de combater porque seus endosporos são muito difíceis de matar. Métodos especiais de esterilização para bactérias formadoras de endosporos são discutidos em Controle do Crescimento Microbiano.
Exercício\(\PageIndex{2}\)
- O que é uma inclusão?
- Qual é a função de um endosporo?
Membrana de
As estruturas que envolvem o citoplasma e as estruturas internas da célula são conhecidas coletivamente como envelope celular. Nas células procarióticas, as estruturas do envelope celular variam dependendo do tipo de célula e do organismo. A maioria (mas não todas) das células procarióticas tem uma parede celular, mas a composição dessa parede celular varia. Todas as células (procarióticas e eucarióticas) têm uma membrana plasmática (também chamada de membrana citoplasmática ou membrana celular) que exibe permeabilidade seletiva, permitindo que algumas moléculas entrem ou saiam da célula enquanto restringem a passagem de outras.
A estrutura da membrana plasmática é frequentemente descrita em termos do modelo de mosaico de fluidos, que se refere à capacidade dos componentes da membrana de se moverem fluidamente dentro do plano da membrana, bem como à composição em forma de mosaico dos componentes, que incluem uma gama diversificada de componentes lipídicos e proteicos ( Figura\(\PageIndex{10}\)). A estrutura da membrana plasmática da maioria dos tipos de células bacterianas e eucarióticas é uma bicamada composta principalmente por fosfolipídios formados por ligações ésteres e proteínas. Esses fosfolipídios e proteínas têm a capacidade de se mover lateralmente dentro do plano das membranas, bem como entre as duas camadas de fosfolipídios.

As membranas arqueais são fundamentalmente diferentes das membranas bacterianas e eucarióticas em alguns aspectos significativos. Primeiro, os fosfolipídios da membrana arqueal são formados com ligações etéreas, em contraste com as ligações ésteres encontradas nas membranas celulares bacterianas ou eucarióticas. Em segundo lugar, os fosfolipídios arqueais têm cadeias ramificadas, enquanto os das células bacterianas e eucarióticas são de cadeia reta. Finalmente, embora algumas membranas arqueais possam ser formadas por bicamadas, como as encontradas em bactérias e eucariotos, outras membranas plasmáticas arqueais são monocamadas lipídicas.
As proteínas na superfície da célula são importantes para uma variedade de funções, incluindo a comunicação entre células e a detecção de condições ambientais e fatores de virulência patogênicos. As proteínas de membrana e os fosfolipídios podem ter carboidratos (açúcares) associados a eles e são chamados de glicoproteínas ou glicolipídios, respectivamente. Esses complexos de glicoproteínas e glicolipídios se estendem para fora da superfície da célula, permitindo que a célula interaja com o ambiente externo (Figura\(\PageIndex{10}\)). As glicoproteínas e os glicolipídios na membrana plasmática podem variar consideravelmente na composição química entre arquéias, bactérias e eucariotos, permitindo que os cientistas os usem para caracterizar espécies únicas.
As membranas plasmáticas de diferentes tipos de células também contêm fosfolipídios exclusivos, que contêm ácidos graxos. Conforme descrito em Usando a bioquímica para identificar microrganismos, os perfis de análise de ácidos graxos derivados de fosfolipídios (PLFA) podem ser usados para identificar tipos únicos de células com base nas diferenças nos ácidos graxos. Arqueias, bactérias e eucariotos têm um perfil exclusivo de PFLA.
Mecanismos de transporte de
Uma das funções mais importantes da membrana plasmática é controlar o transporte de moléculas para dentro e para fora da célula. As condições internas devem ser mantidas dentro de um determinado intervalo, apesar de quaisquer mudanças no ambiente externo. O transporte de substâncias pela membrana plasmática permite que as células o façam.
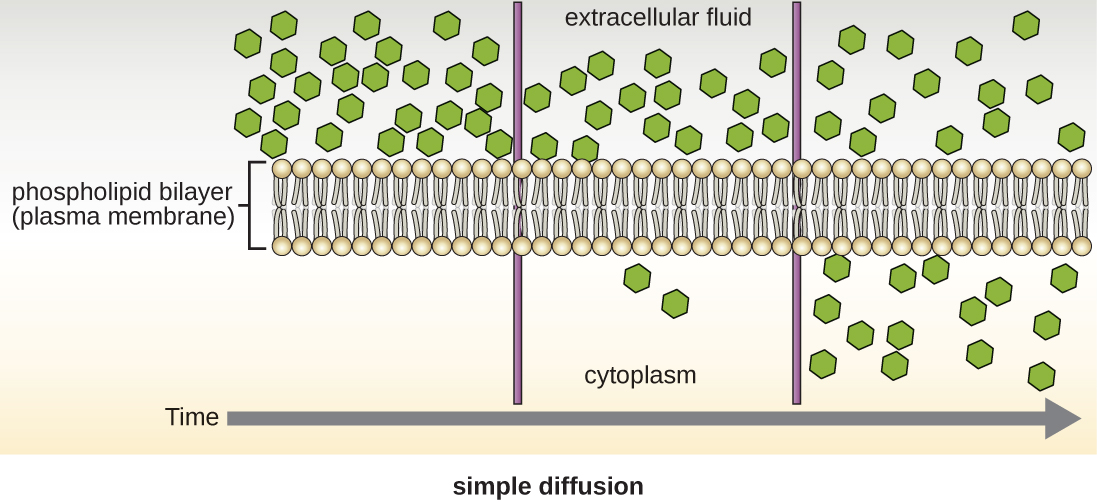
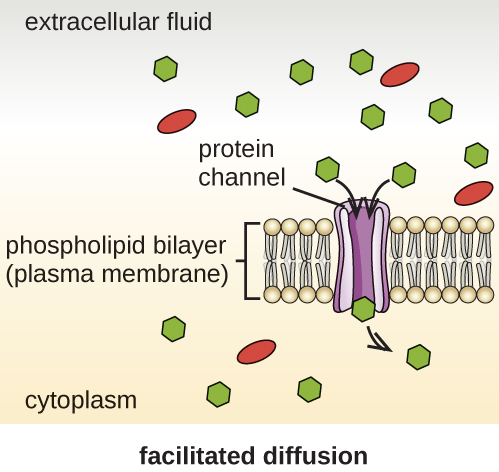
As células usam vários modos de transporte através da membrana plasmática. Por exemplo, moléculas que se movem de uma concentração mais alta para uma menor com o gradiente de concentração são transportadas por difusão simples, também conhecida como transporte passivo (Figura\(\PageIndex{11}\)). Algumas moléculas pequenas, como o dióxido de carbono, podem atravessar a bicamada da membrana diretamente por simples difusão. No entanto, moléculas carregadas, assim como moléculas grandes, precisam da ajuda de transportadores ou canais na membrana. Essas estruturas transportam moléculas pela membrana, um processo conhecido como difusão facilitada (Figura\(\PageIndex{12}\)).
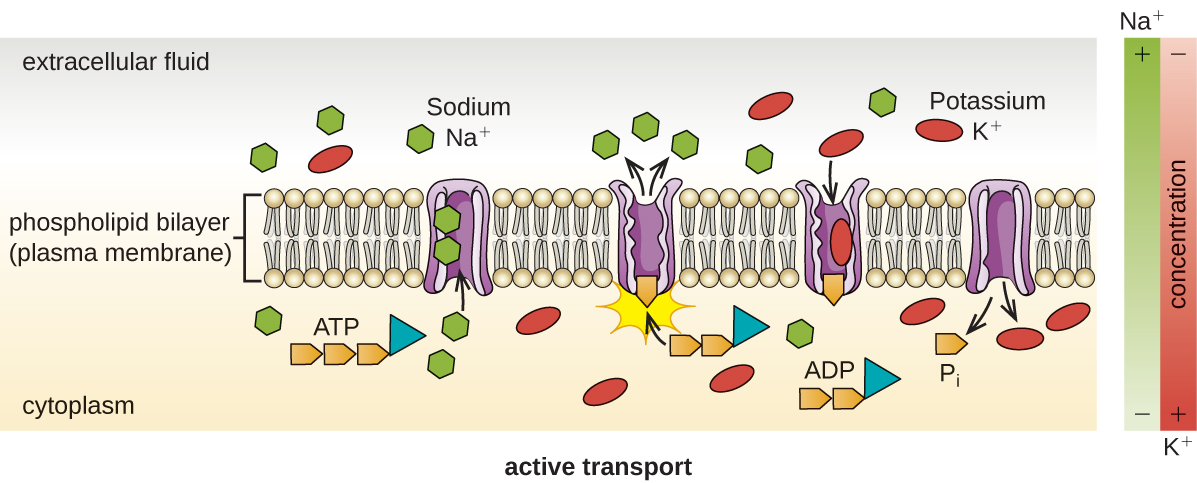
O transporte ativo ocorre quando as células movem moléculas através de sua membrana contra gradientes de concentração (Figura\(\PageIndex{13}\)). Uma grande diferença entre transporte passivo e ativo é que o transporte ativo requer trifosfato de adenosina (ATP) ou outras formas de energia para mover as moléculas “morro acima”. Portanto, as estruturas de transporte ativas são freqüentemente chamadas de “bombas”.
A translocação de grupos também transporta substâncias para as células bacterianas. Nesse caso, à medida que uma molécula se move para dentro de uma célula contra seu gradiente de concentração, ela é quimicamente modificada para não requerer transporte contra um gradiente de concentração desfavorável. Um exemplo comum disso é o sistema de fosfotransferase bacteriana, uma série de transportadores que fosforila (ou seja, adiciona íons fosfato à) glicose ou outros açúcares ao entrar nas células. Como a fosforilação dos açúcares é necessária durante os estágios iniciais do metabolismo do açúcar, o sistema fosfotransferase é considerado um sistema de energia neutra.
Estruturas de membrana fotossint
Algumas células procarióticas, nomeadamente cianobactérias e bactérias fotossintéticas, têm estruturas de membrana que lhes permitem realizar a fotossíntese. Essas estruturas consistem em um desdobramento da membrana plasmática que envolve pigmentos fotossintéticos, como clorofilas verdes e bacterioclorofilas. Nas cianobactérias, essas estruturas de membrana são chamadas de tilacóides; nas bactérias fotossintéticas, elas são chamadas de cromatóforos, lamelas ou clorossomos.
Parede celular
A função principal da parede celular é proteger a célula de condições adversas no ambiente externo. Quando presentes, existem semelhanças e diferenças notáveis entre as paredes celulares de arquéias, bactérias e eucariotos.
O principal componente das paredes celulares bacterianas é chamado peptidoglicano (ou mureína); só é encontrado em bactérias. Estruturalmente, o peptidoglicano se assemelha a uma camada de malha ou tecido (Figura\(\PageIndex{14}\)). Cada camada é composta por longas cadeias de moléculas alternadas de N-acetilglucosamina (NAG) e ácido N-acetilmurâmico (NAM). A estrutura das cadeias longas tem uma resistência à tração bidimensional significativa devido à formação de pontes peptídicas que conectam NAG e NAM dentro de cada camada de peptidoglicano. Nas bactérias gram-negativas, as cadeias de tetrapeptídeos que se estendem de cada unidade NAM são diretamente reticuladas, enquanto nas bactérias gram-positivas, essas cadeias de tetrapeptídeos são ligadas por pontes cruzadas de pentaglicina. As subunidades do peptidoglicano são produzidas dentro da célula bacteriana e depois exportadas e montadas em camadas, dando forma à célula.
Como o peptidoglicano é exclusivo das bactérias, muitos antibióticos são projetados para interferir na síntese do peptidoglicano, enfraquecendo a parede celular e tornando as células bacterianas mais suscetíveis aos efeitos da pressão osmótica (consulte Mecanismos de medicamentos antibacterianos). Além disso, certas células do sistema imunológico humano são capazes de “reconhecer” patógenos bacterianos detectando peptidoglicano na superfície de uma célula bacteriana; essas células então engolfam e destroem a célula bacteriana, usando enzimas como a lisozima, que decompõe e digere o peptidoglicano em suas paredes celulares (veja Reconhecimento de Patógenos e Fagocitose).
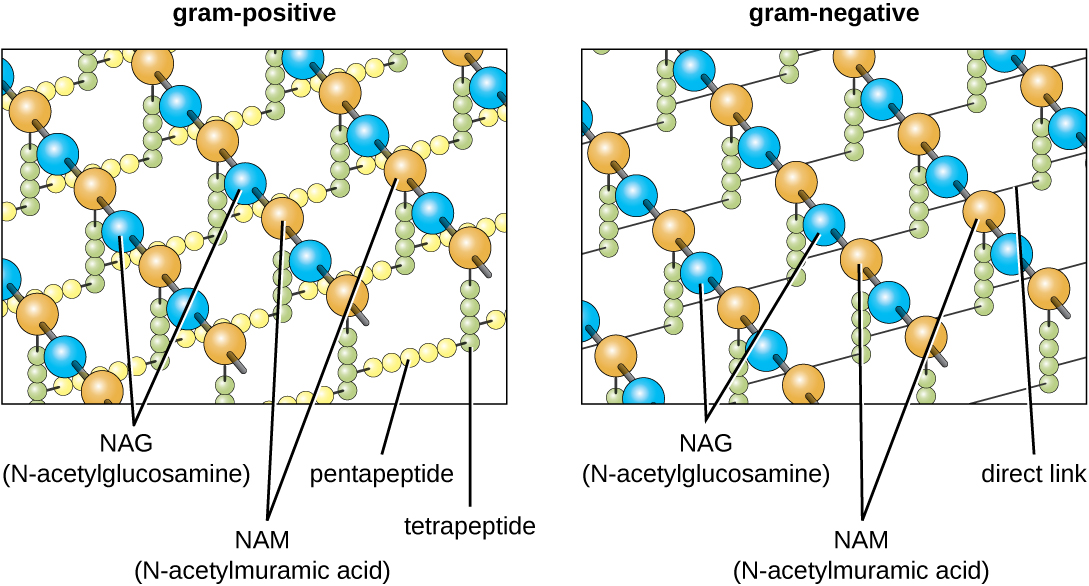
O protocolo de coloração de Gram (consulte Coloração de amostras microscópicas) é usado para diferenciar dois tipos comuns de estruturas da parede celular (Figura\(\PageIndex{15}\)). As células Gram-positivas têm uma parede celular que consiste em muitas camadas de peptidoglicano totalizando 30—100 nm de espessura. Essas camadas de peptidoglicano são comumente incorporadas com ácidos teicóicos (TAs), cadeias de carboidratos que se estendem através e além da camada de peptidoglicano. 4 Acredita-se que o TA estabilize o peptidoglicano aumentando sua rigidez. A TA também desempenha um papel na capacidade de bactérias gram-positivas patogênicas, como o Streptococcus, de se ligarem a certas proteínas na superfície das células hospedeiras, aumentando sua capacidade de causar infecções. Além do peptidoglicano e dos TAs, as bactérias da família Mycobacteriaceae têm uma camada externa de ácidos micólicos cerosos em sua parede celular; conforme descrito em Espécimes Microscópicos de Coloração, essas bactérias são chamadas de ácido-resistentes, uma vez que manchas ácido-resistentes devem ser usadas para penetrar no micólico camada ácida para fins de microscopia (Figura\(\PageIndex{16}\)).

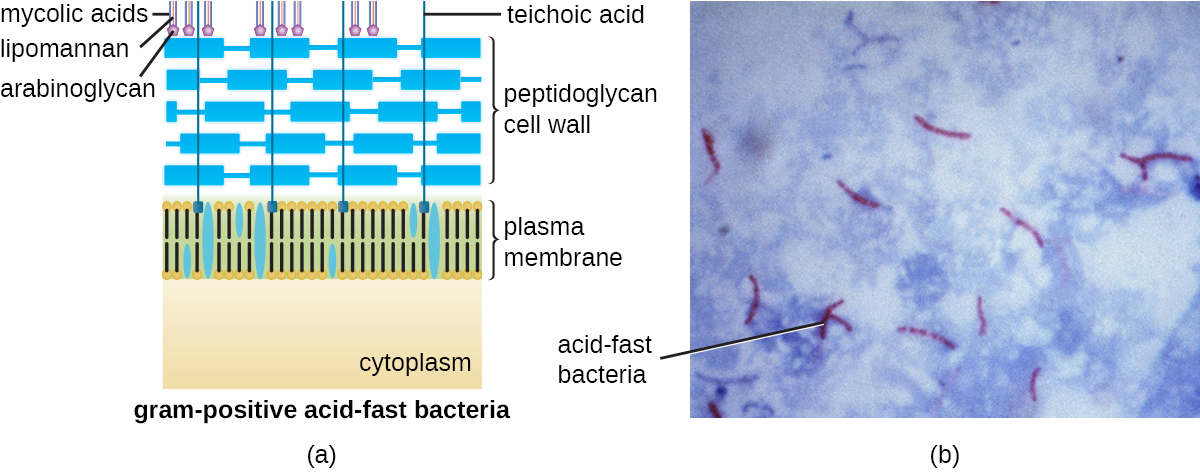
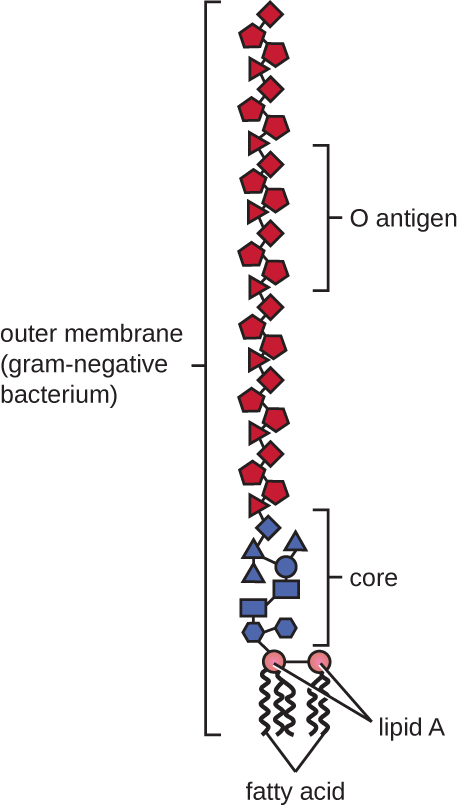
As células gram-negativas têm uma camada muito mais fina de peptidoglicano (não mais do que cerca de 4 nm de espessura 6) do que as células gram-positivas, e a estrutura geral de seu envelope celular é mais complexa. Nas células gram-negativas, uma matriz gelatinosa ocupa o espaço periplasmático entre a parede celular e a membrana plasmática, e há uma segunda bicamada lipídica chamada membrana externa, que é externa à camada de peptidoglicano (Figura\(\PageIndex{15}\)). Essa membrana externa está ligada ao peptidoglicano pela lipoproteína mureína. O folheto externo da membrana externa contém a molécula lipopolissacarídeo (LPS), que funciona como uma endotoxina em infecções envolvendo bactérias gram-negativas, contribuindo para sintomas como febre, hemorragia e choque séptico. Cada molécula de LPS é composta por lipídio A, um polissacarídeo central e uma cadeia lateral O que é composta por moléculas semelhantes ao açúcar que compõem a face externa do LPS (Figura\(\PageIndex{17}\)). A composição da cadeia lateral O varia entre diferentes espécies e cepas de bactérias. Partes da cadeia lateral O chamadas antígenos podem ser detectadas usando testes sorológicos ou imunológicos para identificar cepas patogênicas específicas, como a Escherichia coli O157:H7, uma cepa mortal de bactéria que causa diarreia com sangue e insuficiência renal.
A estrutura da parede celular arqueal difere da das bactérias de várias maneiras significativas. Primeiro, as paredes celulares arqueais não contêm peptidoglicano; em vez disso, elas contêm um polímero similar chamado pseudopeptidoglicano (pseudomureína) no qual o NAM é substituído por uma subunidade diferente. Outras arquéias podem ter uma camada de glicoproteínas ou polissacarídeos que serve como parede celular em vez de pseudopeptidoglicano. Por último, como é o caso de algumas espécies bacterianas, existem algumas arquéias que parecem não ter paredes celulares por completo.
Glicocálicos e camadas S
Embora a maioria das células procarióticas tenha paredes celulares, algumas podem ter estruturas adicionais de envelope celular externas à parede celular, como glicocálicos e camadas S. Um glicocálix é uma camada de açúcar, da qual existem dois tipos importantes: cápsulas e camadas de lodo. Uma cápsula é uma camada organizada localizada fora da parede celular e geralmente composta por polissacarídeos ou proteínas (Figura\(\PageIndex{18}\)). Uma camada de lodo é uma camada menos organizada que só está frouxamente presa à parede celular e pode ser lavada com mais facilidade. As camadas de limo podem ser compostas por polissacarídeos, glicoproteínas ou glicolipídios.
Os glicocálicos permitem que as células adiram às superfícies, auxiliando na formação de biofilmes (colônias de micróbios que se formam em camadas nas superfícies). Na natureza, a maioria dos micróbios vive em comunidades mistas dentro dos biofilmes, em parte porque o biofilme lhes oferece algum nível de proteção. Os biofilmes geralmente retêm água como uma esponja, evitando a dessecação. Eles também protegem as células da predação e impedem a ação de antibióticos e desinfetantes. Todas essas propriedades são vantajosas para os micróbios que vivem em um biofilme, mas apresentam desafios em um ambiente clínico, onde o objetivo geralmente é eliminar os micróbios.

A capacidade de produzir uma cápsula pode contribuir para a patogenicidade de um micróbio (capacidade de causar doenças) porque a cápsula pode dificultar a absorção e a morte de células fagocíticas (como os glóbulos brancos). Streptococcus pneumoniae, por exemplo, produz uma cápsula que é bem conhecida por auxiliar na patogenicidade dessa bactéria. Conforme explicado em Coloração de amostras microscópicas, as cápsulas são difíceis de colorir para microscopia; técnicas de coloração negativa são normalmente usadas.
Uma camada S é outro tipo de estrutura do envelope celular; ela é composta por uma mistura de proteínas estruturais e glicoproteínas. Nas bactérias, as camadas S são encontradas fora da parede celular, mas em algumas arquéias, a camada S serve como parede celular. A função exata das camadas S não é totalmente compreendida e elas são difíceis de estudar; mas as evidências disponíveis sugerem que elas podem desempenhar uma variedade de funções em diferentes células procarióticas, como ajudar a célula a suportar a pressão osmótica e, para certos patógenos, interagir com o sistema imunológico do hospedeiro sistema.
Foco clínico: Parte 3
Depois de diagnosticar Barbara com pneumonia, o PA prescreve a ela uma receita de amoxicilina, um tipo de derivado da penicilina comumente prescrito. Mais de uma semana depois, apesar de fazer o curso completo conforme as instruções, Barbara ainda se sente fraca e não está totalmente recuperada, embora ainda seja capaz de realizar suas atividades diárias. Ela retorna ao centro de saúde para uma consulta de acompanhamento.
Muitos tipos de bactérias, fungos e vírus podem causar pneumonia. A amoxicilina tem como alvo o peptidoglicano das paredes celulares bacterianas. Como a amoxicilina não resolveu os sintomas de Barbara, o AP conclui que o agente causador provavelmente não tem peptidoglicano, o que significa que o patógeno pode ser um vírus, um fungo ou uma bactéria sem peptidoglicano. Outra possibilidade é que o patógeno seja uma bactéria contendo peptidoglicano, mas tenha desenvolvido resistência à amoxicilina.
Exercício\(\PageIndex{3}\)
- Como a AP pode identificar definitivamente a causa da pneumonia de Barbara?
- Que forma de tratamento a PA deve prescrever, uma vez que a amoxicilina foi ineficaz?
Apêndices filamentosos
Muitas células bacterianas têm apêndices proteicos embutidos em seus envelopes celulares que se estendem para fora, permitindo a interação com o meio ambiente. Esses apêndices podem se fixar em outras superfícies, transferir DNA ou fornecer movimento. Os apêndices filamentosos incluem fímbrias, pili e flagelos.
Fimbriae e Pili
As fímbrias e os pili são estruturalmente semelhantes e, como a diferenciação entre os dois é problemática, esses termos são frequentemente usados de forma intercambiável. 7 8 O termo fímbrias geralmente se refere a proteínas semelhantes a cerdas curtas que se projetam da superfície celular às centenas. As fímbrias permitem que uma célula se fixe em superfícies e outras células. Para bactérias patogênicas, a adesão às células hospedeiras é importante para colonização, infectividade e virulência. A aderência às superfícies também é importante na formação de biofilme.
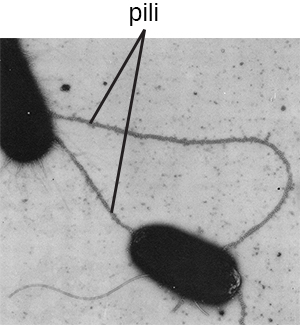
O termo pili (singular: pilus) geralmente se refere a apêndices proteicos mais longos e menos numerosos que ajudam na fixação às superfícies (Figura\(\PageIndex{19}\)). Um tipo específico de pilus, chamado de F pilus ou pilus sexual, é importante na transferência de DNA entre células bacterianas, que ocorre entre membros da mesma geração quando duas células transferem ou trocam fisicamente partes de seus respectivos genomas (veja Como os procariontes assexuais alcançam a genética Diversidade).
Antes que a estrutura e a função dos vários componentes do envelope celular bacteriano fossem bem compreendidas, os cientistas já estavam usando as características do envelope celular para classificar as bactérias. Em 1933, Rebecca Lancefield propôs um método para sorotipar várias cepas β-hemolíticas de espécies de Streptococcus usando um ensaio de aglutinação, uma técnica que usa a aglomeração de bactérias para detectar antígenos específicos da superfície celular. Ao fazer isso, Lancefield descobriu que um grupo de S. pyogenes, encontrado no Grupo A, estava associado a uma variedade de doenças humanas. Ela determinou que várias cepas de estreptococos do Grupo A poderiam ser distinguidas umas das outras com base nas variações em proteínas específicas da superfície celular que ela chamou de proteínas M.
Hoje, mais de 80 cepas diferentes de estreptococos do Grupo A foram identificadas com base nas proteínas M. Várias cepas de estreptococos do Grupo A estão associadas a uma ampla variedade de infecções humanas, incluindo faringite estreptocócica (faringite estreptocócica), impétigo, síndrome do choque tóxico, escarlatina, febre reumática e fascite necrosante. A proteína M é um importante fator de virulência para o estreptococo do grupo A, ajudando essas cepas a fugirem do sistema imunológico. Alterações nas proteínas M parecem alterar a infectividade de uma cepa específica do estreptococo do Grupo A.
Flagelos
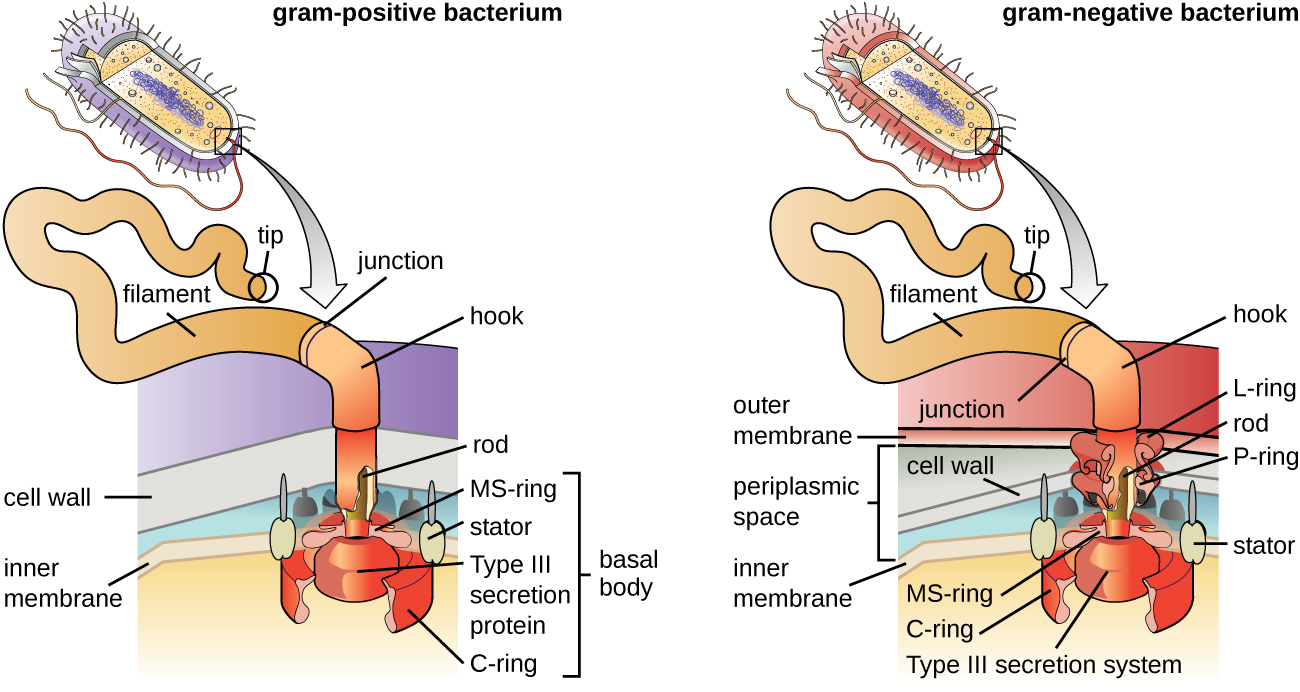
Os flagelos são estruturas usadas pelas células para se mover em ambientes aquosos. Os flagelos bacterianos agem como hélices. Eles são filamentos espirais rígidos compostos por subunidades da proteína flagelina que se estendem para fora da célula e giram em solução. O corpo basal é o motor do flagelo e está embutido na membrana plasmática (Figura\(\PageIndex{20}\)). Uma região de gancho conecta o corpo basal ao filamento. As bactérias gram-positivas e gram-negativas têm diferentes configurações corporais basais devido às diferenças na estrutura da parede celular.
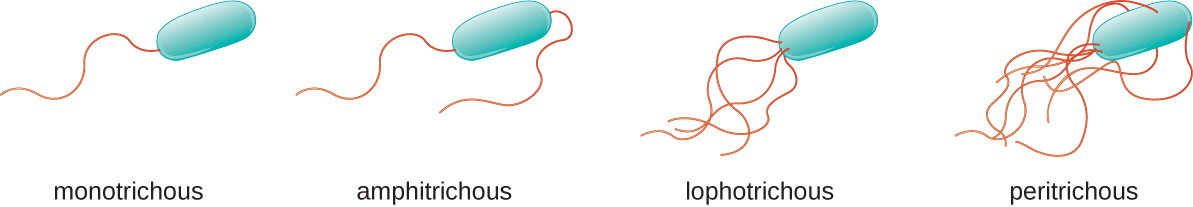
Diferentes tipos de bactérias móveis exibem diferentes arranjos de flagelos (Figura\(\PageIndex{21}\)). Diz-se que uma bactéria com um flagelo singular, normalmente localizada em uma extremidade da célula (polar), tem um flagelo monotricoso. Um exemplo de patógeno bacteriano monotricosamente flagelado é a Vibrio cholerae, a bactéria gram-negativa que causa a cólera. As células com flagelos anfitricos têm um flagelo ou tufos de flagelos em cada extremidade. Um exemplo é o Spirillum minor, a causa da febre espirilar (asiática) por picada de rato ou sodoku. As células com flagelos lofotricosos têm um tufo em uma extremidade da célula. O bacilo gram-negativo Pseudomonas aeruginosa, um patógeno oportunista conhecido por causar muitas infecções, incluindo infecções por “orelha de nadador” e queimaduras, tem flagelos lofotricosos. Os flagelos que cobrem toda a superfície de uma célula bacteriana são chamados de flagelos peritricos. A bactéria gram-negativa E. coli mostra um arranjo peritriquoso de flagelos.
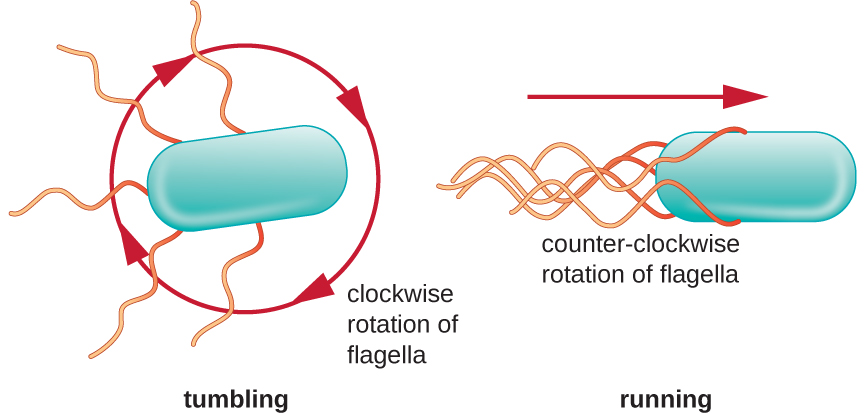
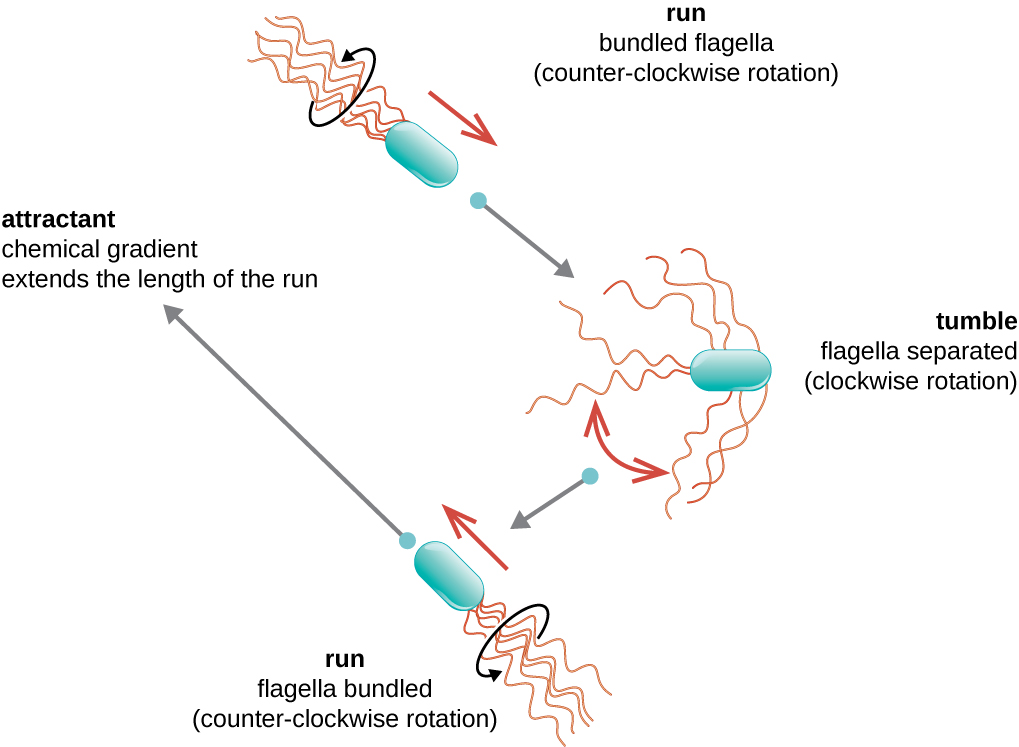
O movimento direcional depende da configuração dos flagelos. As bactérias podem se mover em resposta a uma variedade de sinais ambientais, incluindo luz (fototaxia), campos magnéticos (magnetotaxia) usando magnetossomos e, mais comumente, gradientes químicos (quimiotaxia). O movimento intencional em direção a um atrativo químico, como uma fonte de alimento, ou longe de um repelente, como um produto químico venenoso, é obtido aumentando a duração das corridas e diminuindo a duração das quedas. Ao correr, os flagelos giram no sentido anti-horário, permitindo que a célula bacteriana avance. Em uma bactéria peritricosa, os flagelos são todos agrupados de uma forma muito simplificada (Figura\(\PageIndex{22}\)), permitindo um movimento eficiente. Ao cair, os flagelos são espalhados enquanto giram no sentido horário, criando um movimento em loop e impedindo movimentos significativos para frente, mas reorientando a célula em direção ao atrativo. Quando existe um atrativo, ainda ocorrem corridas e tombos; no entanto, a duração das corridas é maior, enquanto o comprimento dos tombos é reduzido, permitindo um movimento geral em direção à maior concentração do atrativo. Quando não existe gradiente químico, os comprimentos das corridas e tombos são mais iguais e o movimento geral é mais aleatório (Figura\(\PageIndex{23}\)).
Exercício\(\PageIndex{4}\)
- O que é a camada de peptidoglicano e como ela difere entre bactérias gram-positivas e gram-negativas?
- Compare e contraste flagelos monotricos, anfitricos, lofotricosos e períricos.
Resumo
- As células procarióticas diferem das células eucarióticas porque seu material genético está contido em um nucleóide e não em um núcleo ligado à membrana. Além disso, as células procarióticas geralmente não possuem organelas ligadas à membrana.
- Células procarióticas da mesma espécie geralmente compartilham uma morfologia celular e um arranjo celular semelhantes.
- A maioria das células procarióticas tem uma parede celular que ajuda o organismo a manter a morfologia celular e o protege contra mudanças na pressão osmótica.
- Fora do nucleóide, as células procarióticas podem conter DNA extracromossômico em plasmídeos.
- Os ribossomos procarióticos encontrados no citoplasma têm um tamanho de 70S.
- Algumas células procarióticas têm inclusões que armazenam nutrientes ou produtos químicos para outros usos.
- Algumas células procarióticas são capazes de formar endosporos por meio da esporulação para sobreviver em um estado dormente quando as condições são desfavoráveis. Os endosporos podem germinar, transformando-se novamente em células vegetativas quando as condições melhoram.
- Nas células procarióticas, o envelope celular inclui uma membrana plasmática e geralmente uma parede celular.
- As membranas bacterianas são compostas por fosfolipídios com proteínas integrais ou periféricas. Os componentes de ácidos graxos desses fosfolipídios estão ligados aos ésteres e são frequentemente usados para identificar tipos específicos de bactérias. As proteínas desempenham uma variedade de funções, incluindo transporte, comunicação célula a célula e detecção de condições ambientais. As membranas arqueais são distintas por serem compostas por ácidos graxos que estão ligados ao éter aos fosfolipídios.
- Algumas moléculas podem se mover pela membrana bacteriana por simples difusão, mas a maioria das moléculas grandes deve ser transportada ativamente pelas estruturas da membrana usando energia celular.
- As paredes celulares procarióticas podem ser compostas por peptidoglicano (bactéria) ou pseudopeptidoglicano (archaea).
- As células bacterianas gram-positivas são caracterizadas por uma espessa camada de peptidoglicanos, enquanto as células bacterianas gram-negativas são caracterizadas por uma fina camada de peptidoglicano cercada por uma membrana externa.
- Algumas células procarióticas produzem revestimentos de glicocálix, como cápsulas e camadas de lodo, que ajudam na fixação às superfícies e/ou na evasão do sistema imunológico do hospedeiro.
- Algumas células procarióticas têm fímbrias ou pili, apêndices filamentosos que ajudam na fixação às superfícies. Os pili também são usados na transferência de material genético entre as células.
- Algumas células procarióticas usam um ou mais flagelos para se mover pela água. As bactérias peritricosas, que têm numerosos flagelos, usam corridas e tombos para se mover propositalmente na direção de um atrativo químico.
Notas de pé
- 1 Y.-H.M. Chan, W.F. Marshall. “Propriedades de escala do tamanho da célula e da organela”. Organogênese 6 no. 2 (2010) :88—96.
- 2 F. Rothfuss, M. Bender, R. Conrad. “Sobrevivência e atividade de bactérias em um sedimento lacustre profundo e envelhecido (Lago Constança).” Ecologia microbiana 33 no. 1 (1997) :69—77.
- 3 R. Sinclair et al. “Persistência de agentes selecionados da categoria A no ambiente”. Microbiologia Aplicada e Ambiental 74 no. 3 (2008) :555—563.
- 4 T.J. Silhavy, D. Kahne, S. Walker. “O envelope celular bacteriano”. Perspectivas de Cold Spring Harbor em Biologia 2 no. 5 (2010) :a000414.
- 5 B. Zuber et al. “Camada granular no espaço periplasmático de bactérias gram-positivas e estruturas finas dos septos de Enterococcus gallinarum e Streptococcus gordonii revelada por microscopia crio-eletrônica de seções vítreas.” Jornal de Bacteriologia 188 nº 18 (2006) :6652—6660
- 6 L. Gana, S. Chena, G. J. Jensena. “Organização molecular do peptidoglicano gram-negativo”. Anais da Academia Nacional de Ciências dos Estados Unidos da América 105 no. 48 (2008) :18953—18957.
- 7 J.A. Garnetta et al. “Informações estruturais sobre a biogênese e a formação de biofilme pelo pilus comum de Escherichia coli.” Anais da Academia Nacional de Ciências dos Estados Unidos da América 109 no. 10 (2012) :3950—3955.
- 8. T. Profit, E.N. Baker. “Pili em bactérias gram-negativas e gram-positivas — estrutura, montagem e seu papel na doença.” Ciências da Vida Celular e Molecular 66 (2009) :613.