
3.4: Características únicas das células eucarióticas
- Page ID
- 181806

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Objetivos de
- Explicar as características distintivas das células eucarióticas
- Descreva as estruturas internas e externas das células procarióticas em termos de sua estrutura física, estrutura química e função
- Identifique e descreva estruturas e organelas exclusivas das células eucarióticas
- Compare e contraste estruturas similares encontradas em células procarióticas e eucarióticas
Organismos eucarióticos incluem protozoários, algas, fungos, plantas e animais. Algumas células eucarióticas são microrganismos unicelulares independentes, enquanto outras fazem parte de organismos multicelulares. As células dos organismos eucarióticos têm várias características distintivas. Acima de tudo, as células eucarióticas são definidas pela presença de um núcleo cercado por uma membrana nuclear complexa. Além disso, as células eucarióticas são caracterizadas pela presença de organelas ligadas à membrana no citoplasma. Organelas como mitocôndrias, retículo endoplasmático (ER), aparelho de Golgi, lisossomos e peroxissomos são mantidas no lugar pelo citoesqueleto, uma rede interna que suporta o transporte de componentes intracelulares e ajuda a manter a forma celular (Figura\(\PageIndex{1}\)). O genoma das células eucarióticas é empacotado em vários cromossomos em forma de bastonete, em oposição ao cromossomo único de formato circular que caracteriza a maioria das células procarióticas. A tabela\(\PageIndex{1}\) compara as características das estruturas celulares eucarióticas com as de bactérias e arquéias.

| Estrutura celular | Procariontes | Eucariotos | |
|---|---|---|---|
| Bactérias | Archaea | ||
| Tamanho | ~0,5—1 μM | ~0,5—1 μM | ~5—20 μM |
| Relação entre a área de superfície e o volume | Alto | Alto | Baixo |
| Núcleo | Não | Não | sim |
| Características do genoma |
|
|
|
| Divisão celular | Fissão binária | Fissão binária | Mitose, meiose |
| Composição lipídica da membrana |
|
|
|
| Composição da parede celular |
|
|
|
| Estruturas de motilidade | Flagelos espirais rígidos compostos por flagelina | Flagelos espirais rígidos compostos por flagelinos arqueais | Flagelos e cílios flexíveis compostos por microtúbulos |
| Organelas ligadas à membrana | Não | Não | sim |
| Sistema endomembranar | Não | Não | Sim (ER, Golgi, lisossomos) |
| Ribossomos | ANOS 70 | ANOS 70 |
|
Morfologias celulares
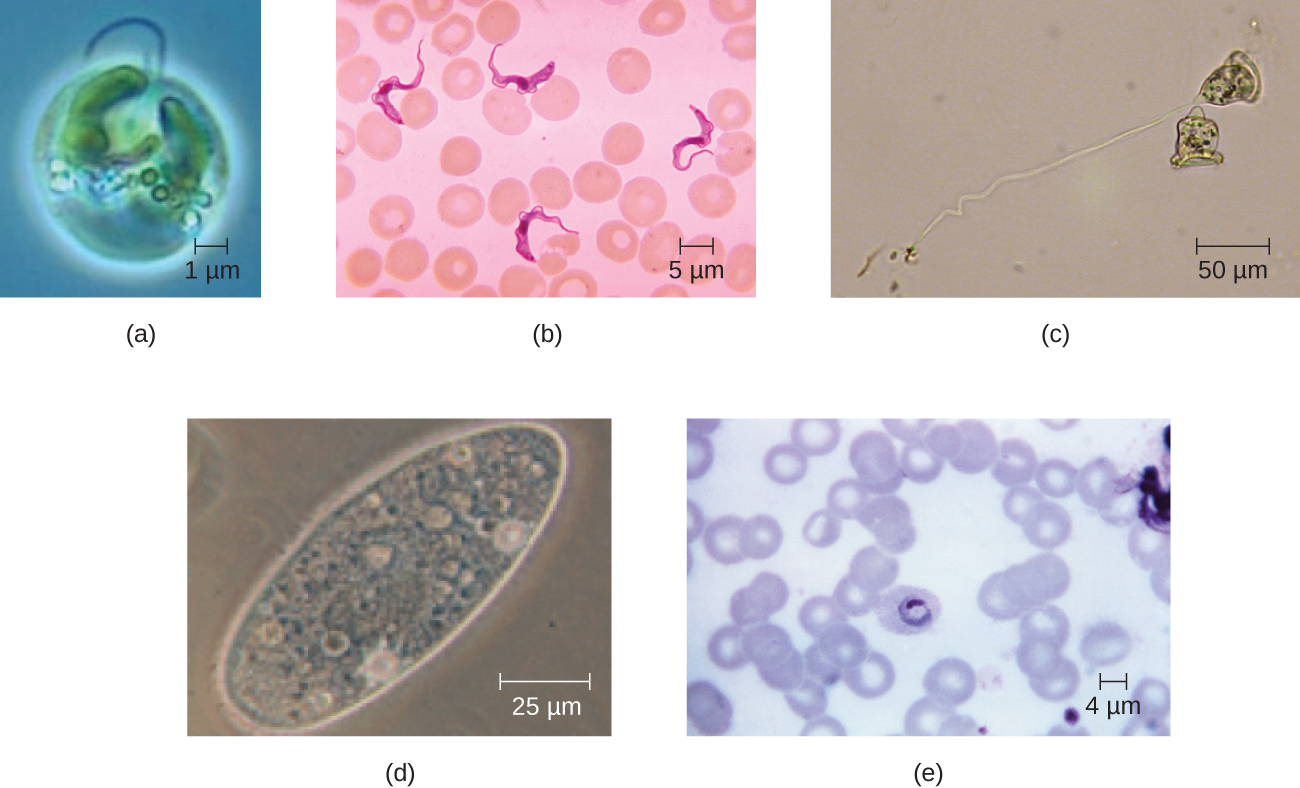
As células eucarióticas apresentam uma grande variedade de morfologias celulares diferentes. As formas possíveis incluem esferóide, ovóide, cuboidal, cilíndrica, plana, lenticular, fusiforme, discoidal, crescente, estrelada em anel e poligonal (Figura\(\PageIndex{2}\)). Algumas células eucarióticas têm formato irregular e outras são capazes de mudar de forma. A forma de um tipo específico de célula eucariótica pode ser influenciada por fatores como sua função primária, a organização de seu citoesqueleto, a viscosidade de seu citoplasma, a rigidez de sua membrana celular ou parede celular (se houver) e a pressão física exercida sobre ela pelo ambiente circundante e/ou células adjacentes.
Exercício\(\PageIndex{1}\)
Identifique duas diferenças entre células eucarióticas e procarióticas.
Núcleo
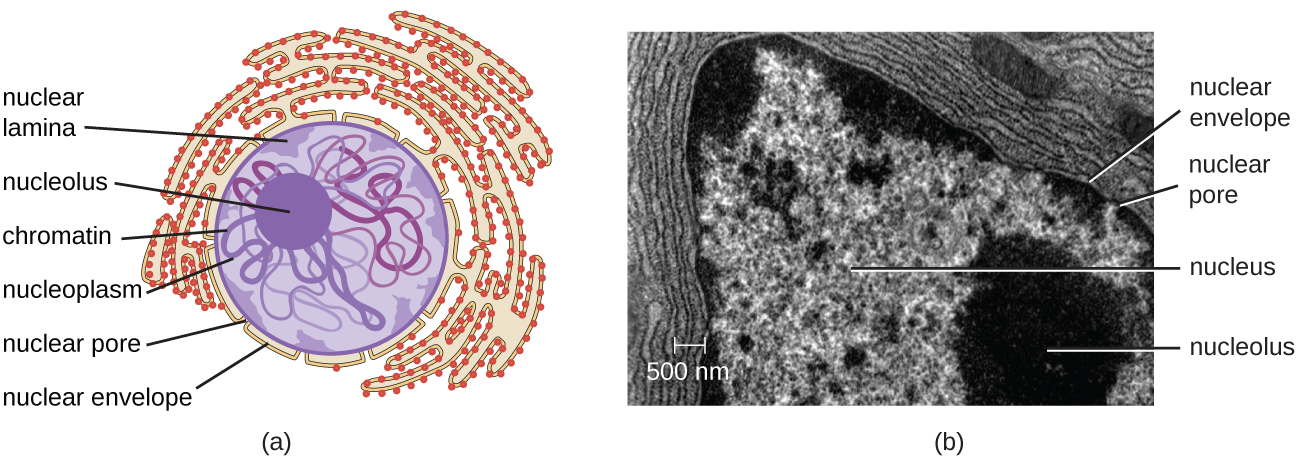
Ao contrário das células procarióticas, nas quais o DNA está frouxamente contido na região nucleóide, as células eucarióticas possuem um núcleo, que é cercado por uma membrana nuclear complexa que abriga o genoma do DNA (Figura\(\PageIndex{3}\)). Ao conter o DNA da célula, o núcleo finalmente controla todas as atividades da célula e também desempenha um papel essencial na reprodução e na hereditariedade. As células eucarióticas normalmente têm seu DNA organizado em vários cromossomos lineares. O DNA dentro do núcleo é altamente organizado e condensado para caber dentro do núcleo, o que é obtido envolvendo o DNA em torno de proteínas chamadas histonas.
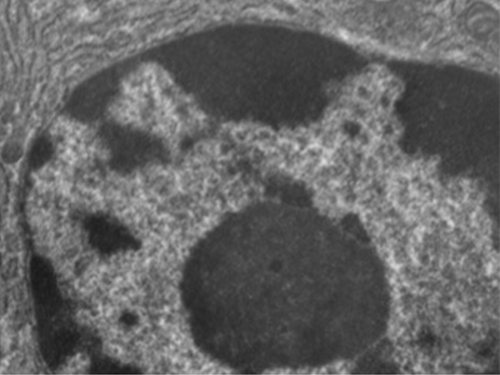
Embora a maioria das células eucarióticas tenha apenas um núcleo, existem exceções. Por exemplo, os protozoários do gênero Paramecium normalmente têm dois núcleos completos: um núcleo pequeno que é usado para reprodução (micronúcleo) e um núcleo grande que direciona o metabolismo celular (macronúcleo). Além disso, alguns fungos formam transitoriamente células com dois núcleos, chamadas células heterocarióticas, durante a reprodução sexual. Células cujos núcleos se dividem, mas cujo citoplasma não se divide, são chamadas de coenócitos.
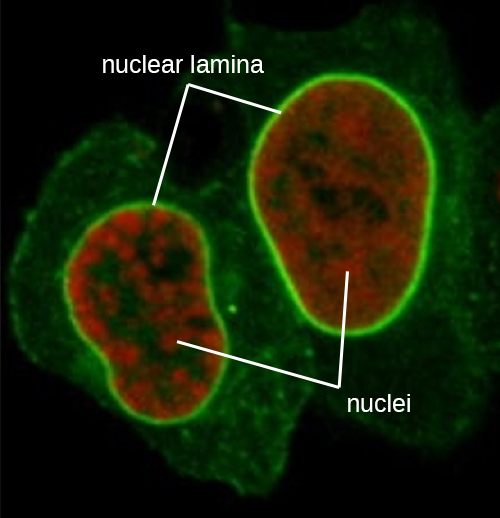
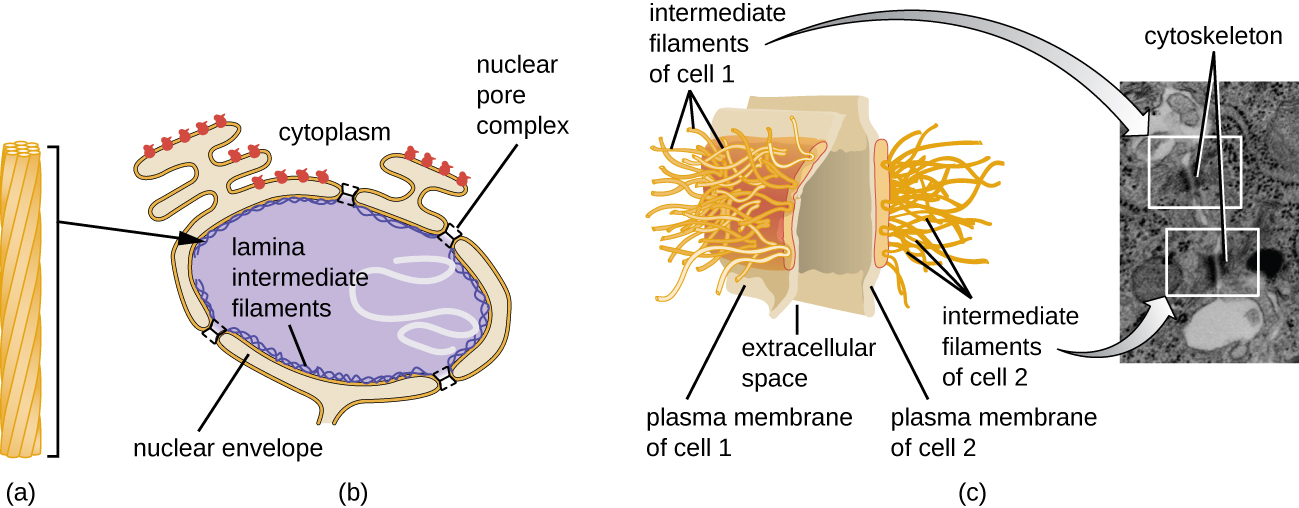
O núcleo é ligado por uma membrana nuclear complexa, frequentemente chamada de envelope nuclear, que consiste em duas bicamadas lipídicas distintas que são contíguas uma à outra (Figura\(\PageIndex{4}\)). Apesar dessas conexões entre as membranas interna e externa, cada membrana contém lipídios e proteínas exclusivos em suas superfícies interna e externa. O envelope nuclear contém poros nucleares, que são grandes complexos proteicos em forma de roseta que controlam o movimento de materiais para dentro e para fora do núcleo. A forma geral do núcleo é determinada pela lâmina nuclear, uma malha de filamentos intermediários encontrada logo dentro das membranas do envelope nuclear. Fora do núcleo, filamentos intermediários adicionais formam uma malha mais solta e servem para ancorar o núcleo em posição dentro da célula.
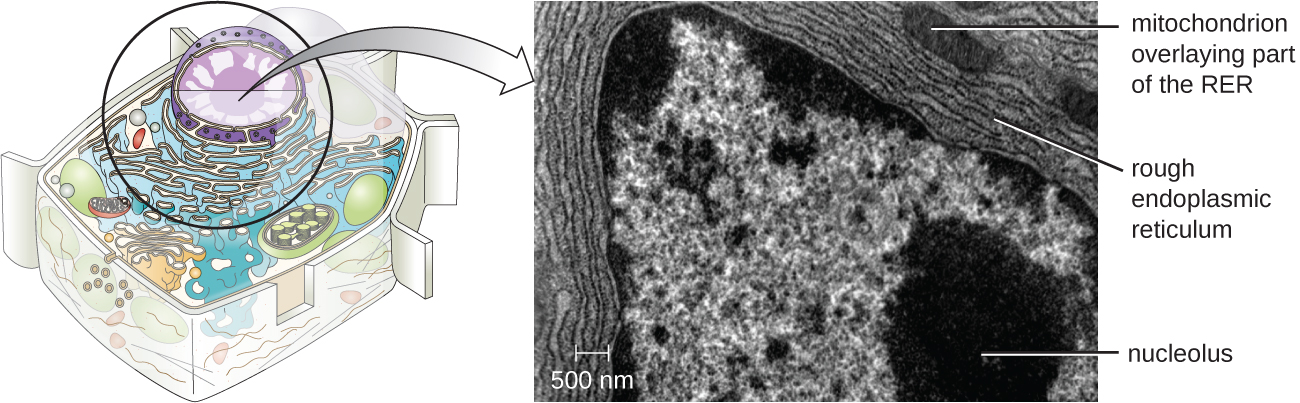
Nucléolo
O nucléolo é uma região densa dentro do núcleo onde ocorre a biossíntese do RNA ribossômico (rRNA). Além disso, o nucléolo também é o local onde começa a montagem dos ribossomos. Os complexos pré-ribossômicos são montados a partir de rRNA e proteínas no nucléolo; eles são então transportados para o citoplasma, onde a montagem do ribossomo é concluída (Figura\(\PageIndex{5}\)).
Ribossomos
Os ribossomos encontrados em organelas eucarióticas, como mitocôndrias ou cloroplastos, têm ribossomos 70S - o mesmo tamanho dos ribossomos procarióticos. No entanto, os ribossomos não associados a organelas em células eucarióticas são ribossomos 80S, compostos por uma pequena subunidade 40S e uma subunidade 60S grande. Em termos de tamanho e composição, isso os torna distintos dos ribossomos das células procarióticas.
Os dois tipos de ribossomos eucarióticos não associados a organelas são definidos por sua localização na célula: ribossomos livres e ribossomos ligados à membrana. Ribossomos livres são encontrados no citoplasma e servem para sintetizar proteínas solúveis em água; ribossomos ligados à membrana são encontrados ligados ao retículo endoplasmático rugoso e produzem proteínas para inserção na membrana celular ou proteínas destinadas à exportação da célula.
As diferenças entre ribossomos eucarióticos e procarióticos são clinicamente relevantes porque certos antibióticos são projetados para atingir um ou outro. Por exemplo, a cicloheximida tem como alvo a ação eucariótica, enquanto o cloranfenicol tem como alvo os ribossomos procarióticos. 1 Como as células humanas são eucarióticas, elas geralmente não são prejudicadas por antibióticos que destroem os ribossomos procarióticos das bactérias. No entanto, às vezes podem ocorrer efeitos colaterais negativos porque as mitocôndrias nas células humanas contêm ribossomos procarióticos.
Sistema Endomembranar
O sistema endomembranoso, exclusivo das células eucarióticas, é uma série de túbulos membranosos, sacos e discos achatados que sintetizam muitos componentes celulares e movem materiais dentro da célula (Figura\(\PageIndex{6}\)). Por causa de seu maior tamanho celular, as células eucarióticas precisam desse sistema para transportar materiais que não podem ser dispersos apenas por difusão. O sistema endomembranar compreende várias organelas e conexões entre elas, incluindo o retículo endoplasmático, o aparelho de Golgi, os lisossomos e as vesículas.
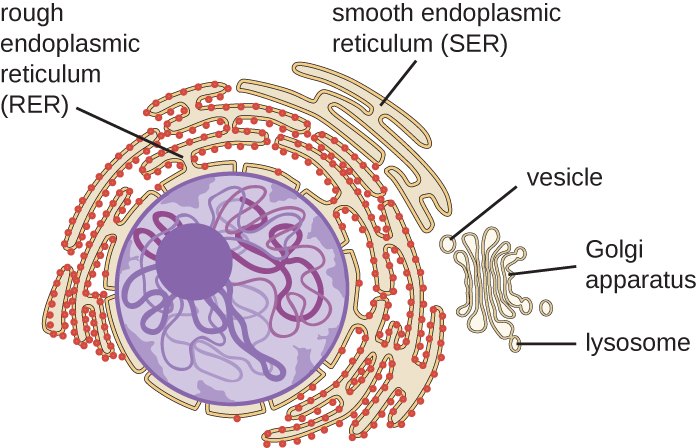
Retículo endoplasmático
O retículo endoplasmático (ER) é uma matriz interconectada de túbulos e cisternas (sacos achatados) com uma única bicamada lipídica (Figura\(\PageIndex{7}\)). Os espaços dentro das cisternas são chamados de lúmen do pronto-socorro. Existem dois tipos de ER, retículo endoplasmático rugoso (RER) e retículo endoplasmático liso (SER). Esses dois tipos diferentes de ER são locais para a síntese de tipos distintamente diferentes de moléculas. O RER é cravejado de ribossomos ligados ao lado citoplasmático da membrana. Esses ribossomos produzem proteínas destinadas à membrana plasmática (Figura\(\PageIndex{}\)). Após a síntese, essas proteínas são inseridas na membrana do RER. Pequenos sacos do RER contendo essas proteínas recém-sintetizadas então brotam como vesículas transportadoras e se movem para o aparelho de Golgi para posterior processamento, diretamente para a membrana plasmática, para a membrana de outra organela ou para fora da célula. As vesículas de transporte são esferas membranosas de lipídios, bicamadas e membranosas com interiores ocos que transportam moléculas. O SER não tem ribossomos e, portanto, parece “suave”. Está envolvido na biossíntese de lipídios, metabolismo de carboidratos e desintoxicação de compostos tóxicos dentro da célula.
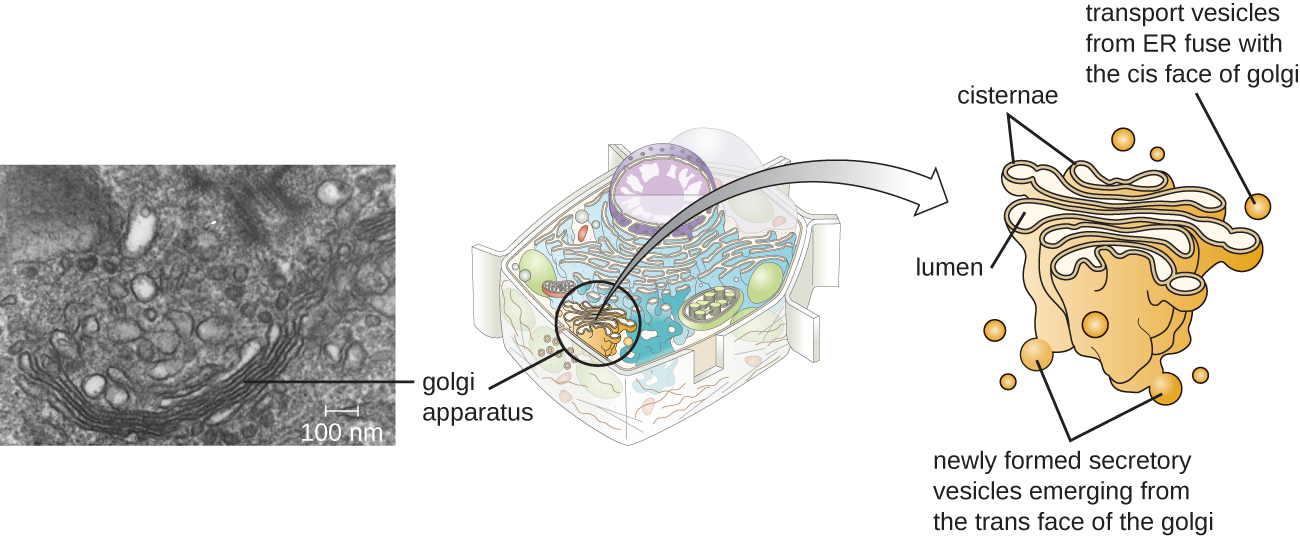
Aparelho de Golgi
O aparelho de Golgi foi descoberto dentro do sistema endomembranar em 1898 pelo cientista italiano Camillo Golgi (1843-1926), que desenvolveu uma nova técnica de coloração que mostrava estruturas de membrana empilhadas dentro das células do Plasmodium, o agente causador da malária. O aparelho de Golgi é composto por uma série de discos membranosos chamados dictiossomos, cada um com uma única bicamada lipídica, que são empilhados juntos (Figura\(\PageIndex{8}\)).
As enzimas do aparelho de Golgi modificam lipídios e proteínas transportados do pronto-socorro para o Golgi, frequentemente adicionando componentes de carboidratos a eles, produzindo glicolipídios, glicoproteínas ou proteoglicanos. Os glicolipídios e glicoproteínas são frequentemente inseridos na membrana plasmática e são importantes para o reconhecimento do sinal por outras células ou partículas infecciosas. Diferentes tipos de células podem ser distinguidos uns dos outros pela estrutura e disposição dos glicolipídios e glicoproteínas contidos em suas membranas plasmáticas. Esses glicolipídios e glicoproteínas geralmente também servem como receptores da superfície celular.
Transporte as vesículas que saem do fusível ER com um aparelho de Golgi em sua face receptora ou cis. As proteínas são processadas dentro do aparelho de Golgi e, em seguida, vesículas de transporte adicionais contendo as proteínas e lipídios modificados se soltam do aparelho de Golgi em sua face de saída, ou trans. Essas vesículas de saída se movem e se fundem com a membrana plasmática ou com a membrana de outras organelas.
A exocitose é o processo pelo qual as vesículas secretoras (sacos membranosos esféricos) liberam seu conteúdo para o exterior da célula (Figura\(\PageIndex{8}\)). Todas as células têm vias secretoras constitutivas nas quais as vesículas secretoras transportam proteínas solúveis que são liberadas da célula continuamente (constitutivamente). Certas células especializadas também têm vias secretoras reguladas, que são usadas para armazenar proteínas solúveis em vesículas secretoras. A secreção regulada envolve substâncias que só são liberadas em resposta a determinados eventos ou sinais. Por exemplo, certas células do sistema imunológico humano (por exemplo, mastócitos) secretam histamina em resposta à presença de objetos estranhos ou patógenos no corpo. A histamina é um composto que aciona vários mecanismos usados pelo sistema imunológico para eliminar patógenos.
Lisossomos
Na década de 1960, o cientista belga Christian de Duve (1917—2013) descobriu os lisossomos, organelas ligadas à membrana do sistema endomembranar que contêm enzimas digestivas. Certos tipos de células eucarióticas usam lisossomos para decompor várias partículas, como alimentos, organelas danificadas ou detritos celulares, microrganismos ou complexos imunes. A compartimentalização das enzimas digestivas dentro do lisossomo permite que a célula digerir a matéria com eficiência sem prejudicar os componentes citoplasmáticos da célula.
Exercício\(\PageIndex{2}\)
Nomeie os componentes do sistema endomembranar e descreva a função de cada componente.
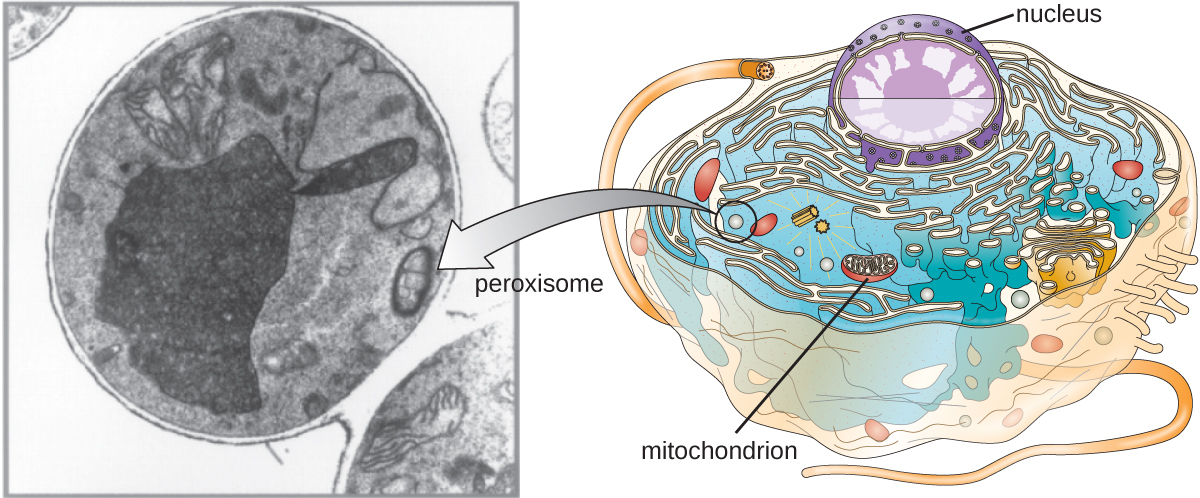
Peroxissomas
Christian de Duve também é creditado com a descoberta de peroxissomos, organelas ligadas à membrana que não fazem parte do sistema endomembranar (Figura\(\PageIndex{9}\)). Os peroxissomas se formam de forma independente no citoplasma a partir da síntese de proteínas de peroxina por ribossomos livres e da incorporação dessas proteínas de peroxina nos peroxissomos existentes. Os peroxissomas em crescimento são então divididos por um processo semelhante à fissão binária.
Os peroxissomos foram nomeados pela primeira vez por sua capacidade de produzir peróxido de hidrogênio, uma molécula altamente reativa que ajuda a quebrar moléculas como ácido úrico, aminoácidos e ácidos graxos. Os peroxissomas também possuem a enzima catalase, que pode degradar o peróxido de hidrogênio. Junto com o SER, os peroxissomas também desempenham um papel na biossíntese de lipídios. Como os lisossomos, a compartimentalização dessas moléculas degradativas dentro de uma organela ajuda a proteger o conteúdo citoplasmático de danos indesejados.
Os peroxissomas de certos organismos são especializados para atender às suas necessidades funcionais específicas. Por exemplo, os glioxissomas são peroxissomos modificados de leveduras e células vegetais que desempenham várias funções metabólicas, incluindo a produção de moléculas de açúcar. Da mesma forma, os glicossomos são peroxissomas modificados produzidos por certos tripanossomas, os protozoários patogênicos que causam a doença de Chagas e a doença do sono africana.
Citoesqueleto
As células eucarióticas têm um citoesqueleto interno feito de microfilamentos, filamentos intermediários e microtúbulos. Essa matriz de fibras e tubos fornece suporte estrutural, bem como uma rede pela qual os materiais podem ser transportados dentro da célula e na qual as organelas podem ser ancoradas (Figura\(\PageIndex{10}\)). Por exemplo, o processo de exocitose envolve o movimento de uma vesícula através da rede citoesquelética até a membrana plasmática, onde ela pode liberar seu conteúdo.
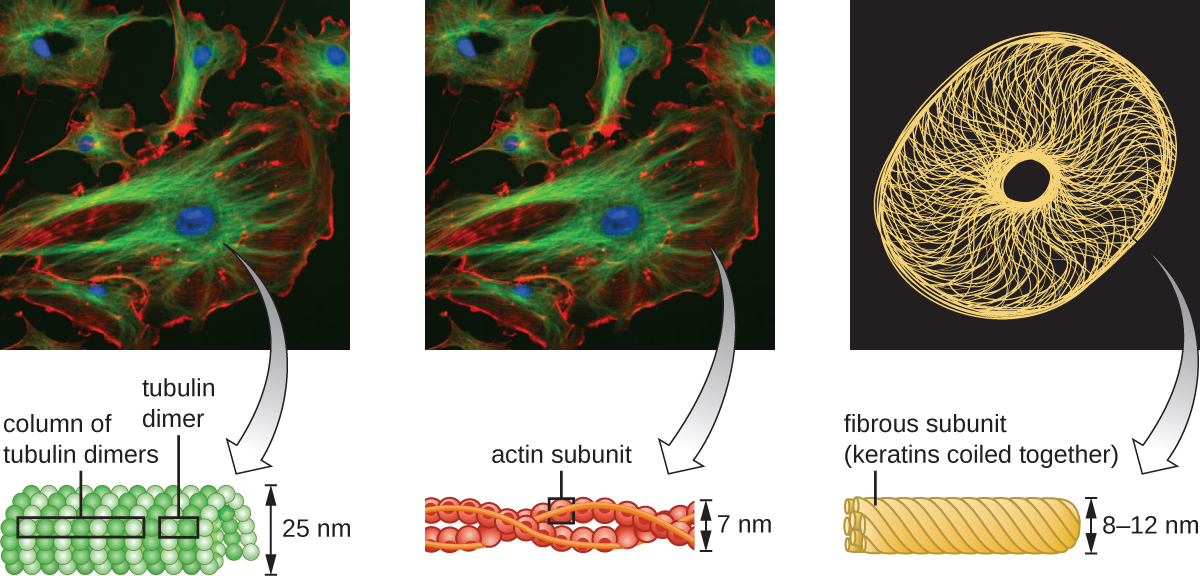
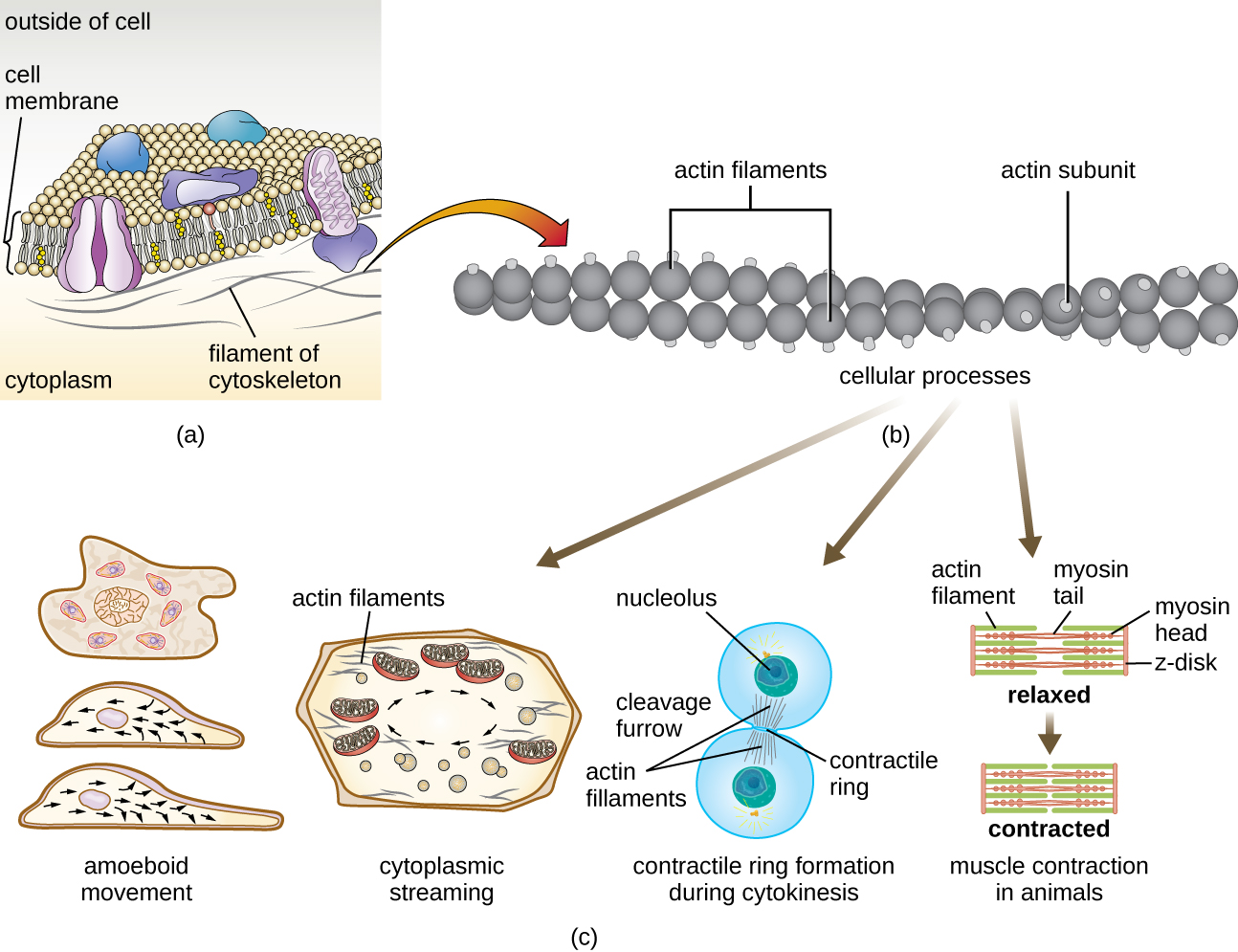
Os microfilamentos são compostos por dois fios entrelaçados de actina, cada um composto por monômeros de actina formando cabos filamentosos de 6 nm de diâmetro 2 (Figura\(\PageIndex{11}\)). Os filamentos de actina trabalham em conjunto com proteínas motoras, como a miosina, para efetuar a contração muscular em animais ou o movimento ameboide de alguns micróbios eucarióticos. Em organismos ameboides, a actina pode ser encontrada em duas formas: uma forma de gel mais rígida e polimerizada e uma forma solúvel mais fluida e não polimerizada. A actina na forma de gel cria estabilidade no ectoplasma, a área gelatinosa do citoplasma logo dentro da membrana plasmática dos protozoários ameboides.
Extensões temporárias da membrana citoplasmática chamadas pseudópodes (que significa “pés falsos”) são produzidas através do fluxo direto de filamentos de actina solúveis para os pseudópodes, seguido pela ciclagem gel-sol dos filamentos de actina, resultando na motilidade celular. Uma vez que o citoplasma se estende para fora, formando um pseudopódio, o citoplasma restante flui para se juntar à borda dianteira, criando assim uma locomoção para frente. Além do movimento ameboide, os microfilamentos também estão envolvidos em uma variedade de outros processos nas células eucarióticas, incluindo fluxo citoplasmático (movimento ou circulação do citoplasma dentro da célula), formação de sulco de clivagem durante a divisão celular e movimento muscular em animais (Figura\(\PageIndex{11}\)). Essas funções são o resultado da natureza dinâmica dos microfilamentos, que podem polimerizar e despolimerizar com relativa facilidade em resposta aos sinais celulares, e de suas interações com motores moleculares em diferentes tipos de células eucarióticas.
Filamentos intermediários (Figura\(\PageIndex{12}\)) são um grupo diverso de filamentos citoesqueléticos que atuam como cabos dentro da célula. Eles são denominados “intermediários” porque seu diâmetro de 10 nm é mais espesso que o da actina, mas mais fino que o dos microtúbulos. 3 Eles são compostos por vários fios de subunidades polimerizadas que, por sua vez, são constituídos por uma grande variedade de monômeros. Filamentos intermediários tendem a ser mais permanentes na célula e manter a posição do núcleo. Eles também formam a lâmina nuclear (revestimento ou camada) logo dentro do envelope nuclear. Além disso, os filamentos intermediários desempenham um papel na ancoragem das células nos tecidos animais. A proteína de filamento intermediário desmina é encontrada nos desmossomos, as estruturas proteicas que unem as células musculares e as ajudam a resistir às forças físicas externas. A proteína de filamento intermediário queratina é uma proteína estrutural encontrada no cabelo, pele e unhas.
Os microtúbulos (Figura\(\PageIndex{13}\)) são um terceiro tipo de fibra citoesquelética composta por dímeros de tubulina (α tubulina e β tubulina). Eles formam tubos ocos de 23 nm de diâmetro que são usados como vigas dentro do citoesqueleto. 4 Como os microfilamentos, os microtúbulos são dinâmicos e têm a capacidade de montar e desmontar rapidamente. Os microtúbulos também funcionam com proteínas motoras (como dineína e cinesina) para mover organelas e vesículas dentro do citoplasma. Além disso, os microtúbulos são os principais componentes dos flagelos e cílios eucarióticos, compondo tanto o filamento quanto os componentes basais do corpo (Figura\(\PageIndex{20}\)).
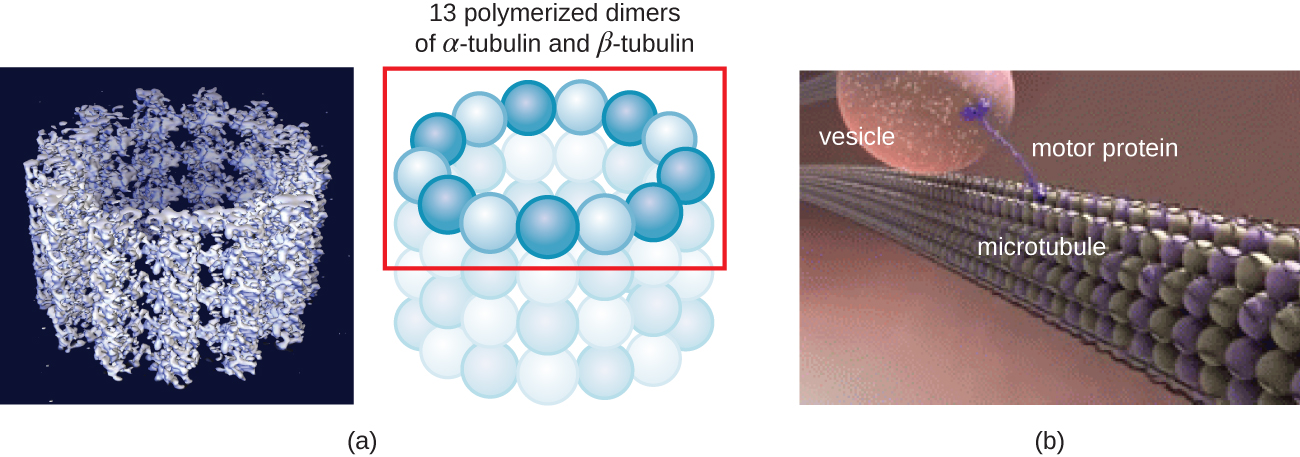
Além disso, os microtúbulos estão envolvidos na divisão celular, formando o fuso mitótico que serve para separar os cromossomos durante a mitose e a meiose. O fuso mitótico é produzido por dois centrossomos, que são essencialmente centros organizadores de microtúbulos, em extremidades opostas da célula. Cada centrossoma é composto por um par de centríolos posicionados em ângulo reto um com o outro, e cada centríolo é uma matriz de nove microtúbulos paralelos dispostos em trigêmeos (Figura\(\PageIndex{14}\)).
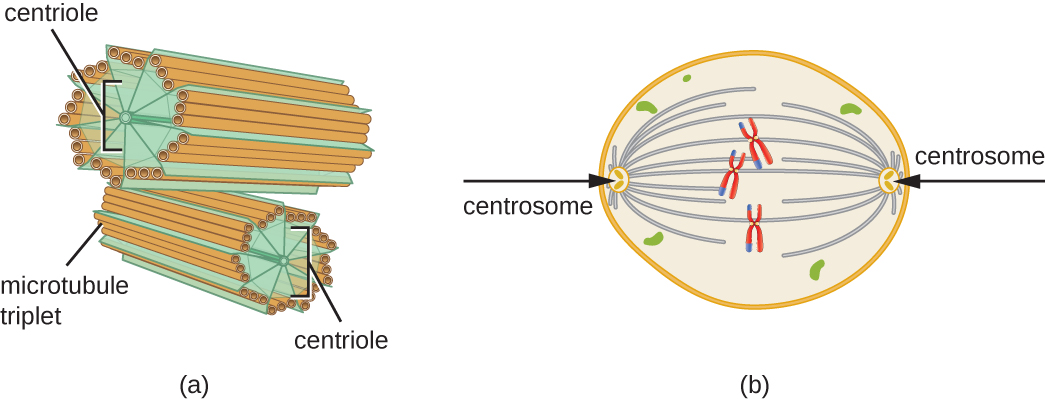
Exercício\(\PageIndex{3}\)
Compare e contraste os três tipos de estruturas do citoesqueleto descritos nesta seção.
Mitocôndrias
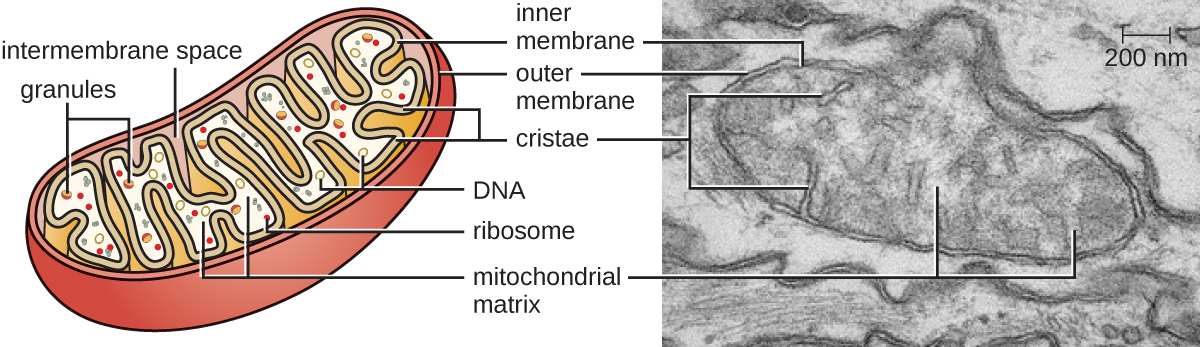
As organelas grandes e complexas nas quais a respiração celular aeróbica ocorre nas células eucarióticas são chamadas de mitocôndrias (Figura\(\PageIndex{15}\)). O termo “mitocôndria” foi cunhado pela primeira vez pelo microbiologista alemão Carl Benda em 1898 e foi posteriormente associado ao processo de respiração por Otto Warburg em 1913. Cientistas durante a década de 1960 descobriram que as mitocôndrias têm seu próprio genoma e ribossomos 70S. Verificou-se que o genoma mitocondrial era bacteriano, quando foi sequenciado em 1976. Em última análise, essas descobertas apoiaram a teoria endossimbiótica proposta por Lynn Margulis, que afirma que as mitocôndrias surgiram originalmente por meio de um evento endossimbiótico no qual uma bactéria capaz de respiração celular aeróbica foi absorvida pela fagocitose em uma célula hospedeira e permaneceu como uma bactéria intracelular viável. componente.
Cada mitocôndria tem duas membranas lipídicas. A membrana externa é um remanescente das estruturas da membrana da célula hospedeira original. A membrana interna foi derivada da membrana plasmática bacteriana. A cadeia de transporte de elétrons para respiração aeróbica usa proteínas integrais embutidas na membrana interna. A matriz mitocondrial, correspondente à localização do citoplasma da bactéria original, é a localização atual de muitas enzimas metabólicas. Ele também contém DNA mitocondrial e ribossomos 70S. As invaginações da membrana interna, chamadas cristas, evoluíram para aumentar a área superficial para a localização das reações bioquímicas. Os padrões de dobramento das cristas diferem entre vários tipos de células eucarióticas e são usados para distinguir diferentes organismos eucarióticos uns dos outros.
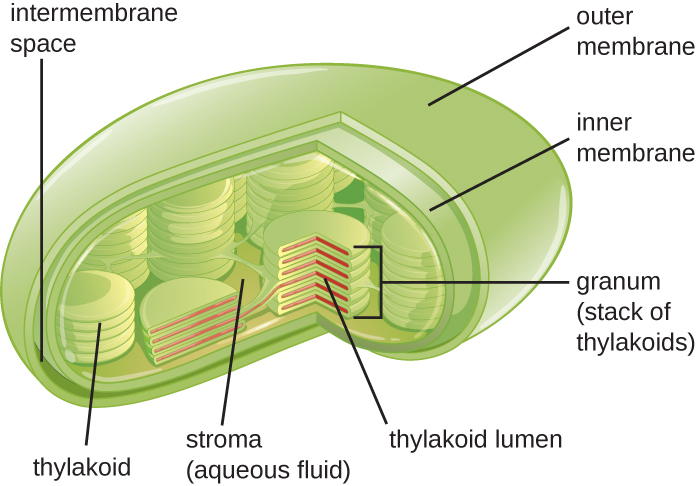
Cloroplastos
As células vegetais e as células de algas contêm cloroplastos, as organelas nas quais ocorre a fotossíntese (Figura\(\PageIndex{16}\)). Todos os cloroplastos têm pelo menos três sistemas de membrana: a membrana externa, a membrana interna e o sistema de membrana tilacóide. Dentro das membranas externa e interna está o estroma do cloroplasto, um fluido gelatinoso que compõe grande parte do volume do cloroplasto e no qual o sistema tilacóide flutua. O sistema tilacóide é uma coleção altamente dinâmica de sacos de membrana dobrados. É onde se encontra a clorofila do pigmento fotossintético verde e ocorrem as reações de luz da fotossíntese. Na maioria dos cloroplastos vegetais, os tilacóides estão dispostos em pilhas chamadas grana (singular: granum), enquanto em alguns cloroplastos de algas, os tilacóides flutuam livremente.
Outras organelas semelhantes às mitocôndrias surgiram em outros tipos de eucariotos, mas suas funções são diferentes. Os hidrogenossomos são encontrados em alguns eucariotos anaeróbicos e servem como o local da produção anaeróbica de hidrogênio. Os hidrogenossomas normalmente não têm seu próprio DNA e ribossomos. Os cinetoplastos são uma variação da mitocôndria encontrada em alguns patógenos eucarióticos. Nesses organismos, cada célula tem uma mitocôndria única, longa e ramificada, na qual o DNA do cinetoplasto, organizado como vários pedaços circulares de DNA, é encontrado concentrado em um polo da célula.
Organelas relacionadas à mitocôndria em parasitas protozoários
Muitos protozoários, incluindo vários parasitas protozoários que causam infecções em humanos, podem ser identificados por sua aparência incomum. As características distintivas podem incluir morfologias celulares complexas, a presença de organelas únicas ou a ausência de organelas comuns. Os protozoários parasitas Giardia lamblia e Trichomonas vaginalis são dois exemplos.
G. lamblia, causa frequente de diarreia em humanos e muitos outros animais, é um parasita anaeróbico que possui dois núcleos e vários flagelos. Seu aparelho de Golgi e seu retículo endoplasmático são bastante reduzidos e carecem completamente de mitocôndrias. No entanto, tem organelas conhecidas como mitossomos, organelas ligadas à membrana dupla que parecem ser mitocôndrias severamente reduzidas. Isso levou os cientistas a acreditar que os ancestrais de G. lamblia já possuíram mitocôndrias que evoluíram para se tornarem mitossomos. O T. vaginalis, que causa a vaginite por infecção sexualmente transmissível, é outro protozoário parasita que não possui mitocôndrias convencionais. Em vez disso, possui hidrogenossomos, organelas mitocondriais ligadas à membrana dupla que produzem hidrogênio molecular usado no metabolismo celular. Os cientistas acreditam que os hidrogenossomos, assim como os mitossomos, também evoluíram das mitocôndrias. 5
Membrana de
A membrana plasmática das células eucarióticas tem estrutura semelhante à membrana plasmática procariótica, pois é composta principalmente por fosfolipídios formando uma bicamada com proteínas periféricas e integrais incorporadas (Figura\(\PageIndex{17}\)). Esses componentes da membrana se movem dentro do plano da membrana de acordo com o modelo de mosaico de fluidos. No entanto, diferentemente da membrana procariótica, as membranas eucarióticas contêm esteróis, incluindo colesterol, que alteram a fluidez da membrana. Além disso, muitas células eucarióticas contêm alguns lipídios especializados, incluindo esfingolípidos, que acredita-se que desempenhem um papel na manutenção da estabilidade da membrana, bem como estejam envolvidos nas vias de transdução de sinal e na comunicação entre células.

Mecanismos de transporte de
Os processos de difusão simples, difusão facilitada e transporte ativo são usados em células eucarióticas e procarióticas. No entanto, as células eucarióticas também têm a capacidade única de realizar vários tipos de endocitose, a absorção de matéria por meio da invaginação da membrana plasmática e a formação de vacúole/vesícula (Figura\(\PageIndex{18}\)). Um tipo de endocitose que envolve o engolfamento de partículas grandes por meio da invaginação da membrana é chamada de fagocitose, que significa “comer células”. Na fagocitose, partículas (ou outras células) são colocadas em uma bolsa dentro da membrana, que então se solta da membrana para formar um vacúolo que envolve completamente a partícula. Outro tipo de endocitose é chamado de pinocitose, que significa “beber células”. Na pinocitose, materiais e líquidos pequenos e dissolvidos são levados para a célula por meio de pequenas vesículas. Os fungos saprofíticos, por exemplo, obtêm seus nutrientes da matéria morta e em decomposição em grande parte por meio da pinocitose.
A endocitose mediada por receptores é um tipo de endocitose que é iniciada por moléculas específicas chamadas ligantes quando elas se ligam aos receptores da superfície celular na membrana. A endocitose mediada por receptores é o mecanismo que os hormônios derivados de peptídeos e aminas usam para entrar nas células e também é usada por vários vírus e bactérias para entrar nas células hospedeiras.
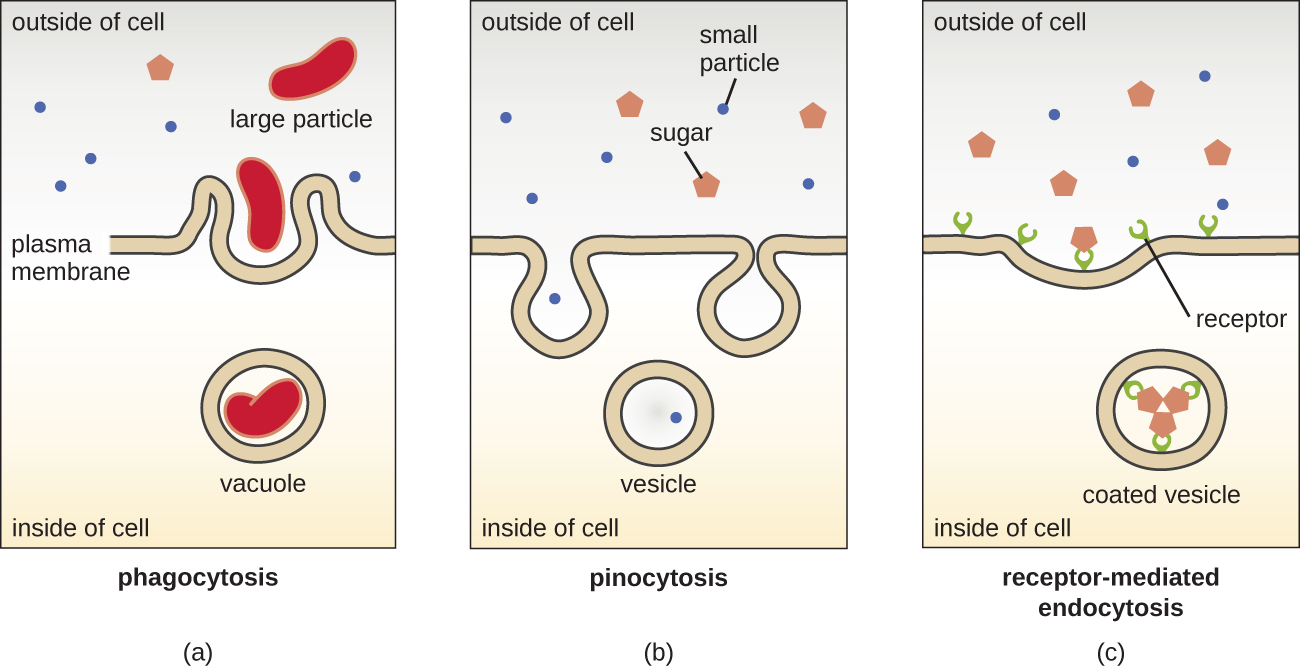
O processo pelo qual as vesículas secretoras liberam seu conteúdo para o exterior da célula é chamado de exocitose. As vesículas se movem em direção à membrana plasmática e depois se fundem com a membrana, ejetando seu conteúdo para fora da célula. A exocitose é usada pelas células para remover resíduos e também pode ser usada para liberar sinais químicos que podem ser captados por outras células.
Parede celular
Além de uma membrana plasmática, algumas células eucarióticas têm uma parede celular. Células de fungos, algas, plantas e até mesmo alguns protistas têm paredes celulares. Dependendo do tipo de célula eucariótica, as paredes celulares podem ser feitas de uma ampla variedade de materiais, incluindo celulose (fungos e plantas); sílica biogênica, carbonato de cálcio, ágar e carragenina (protistas e algas); ou quitina (fungos). Em geral, todas as paredes celulares fornecem estabilidade estrutural para a célula e proteção contra estresses ambientais, como dessecação, mudanças na pressão osmótica e lesões traumáticas. 6
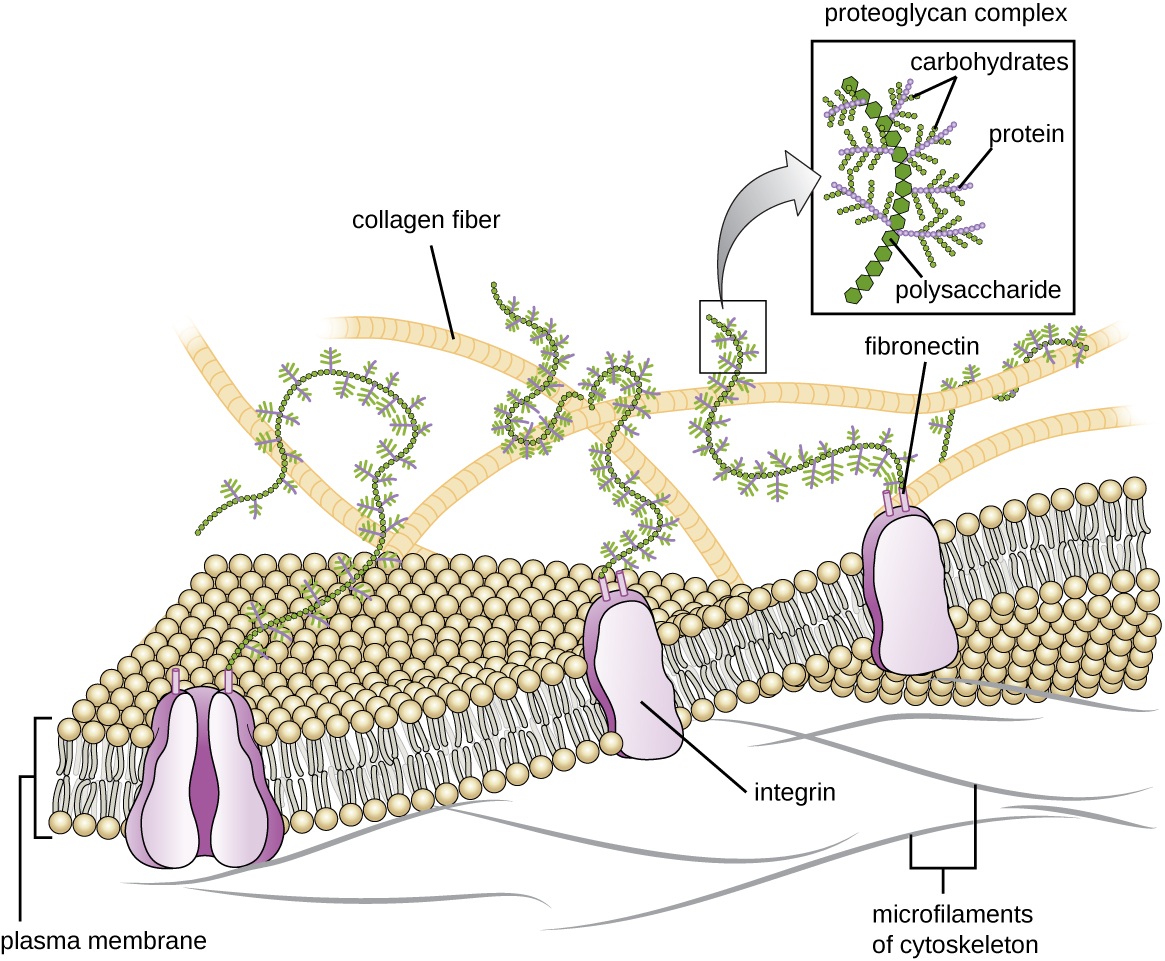
Matriz extracelular
Células de animais e alguns protozoários não têm paredes celulares para ajudar a manter a forma e proporcionar estabilidade estrutural. Em vez disso, esses tipos de células eucarióticas produzem uma matriz extracelular para essa finalidade. Eles secretam uma massa pegajosa de carboidratos e proteínas nos espaços entre as células adjacentes (Figura\(\PageIndex{19}\)). Alguns componentes proteicos se reúnem em uma membrana basal à qual os componentes restantes da matriz extracelular aderem. Os proteoglicanos normalmente formam a massa volumosa da matriz extracelular, enquanto as proteínas fibrosas, como o colágeno, fornecem força. Tanto os proteoglicanos quanto o colágeno estão ligados às proteínas da fibronectina, que, por sua vez, estão ligadas às proteínas da integrina. Essas proteínas integrinas interagem com proteínas transmembranares nas membranas plasmáticas de células eucarióticas que não possuem paredes celulares.
Nas células animais, a matriz extracelular permite que as células dentro dos tecidos resistam a tensões externas e transmitam sinais de fora da célula para dentro. A quantidade de matriz extracelular é bastante extensa em vários tipos de tecidos conjuntivos, e variações na matriz extracelular podem dar a diferentes tipos de tecidos suas propriedades distintas. Além disso, a matriz extracelular da célula hospedeira costuma ser o local onde os patógenos microbianos se ligam para estabelecer a infecção. Por exemplo, Streptococcus pyogenes, a bactéria que causa faringite estreptocócica e várias outras infecções, liga-se à fibronectina na matriz extracelular das células que revestem a orofaringe (região superior da garganta).
Flagella e Cilia
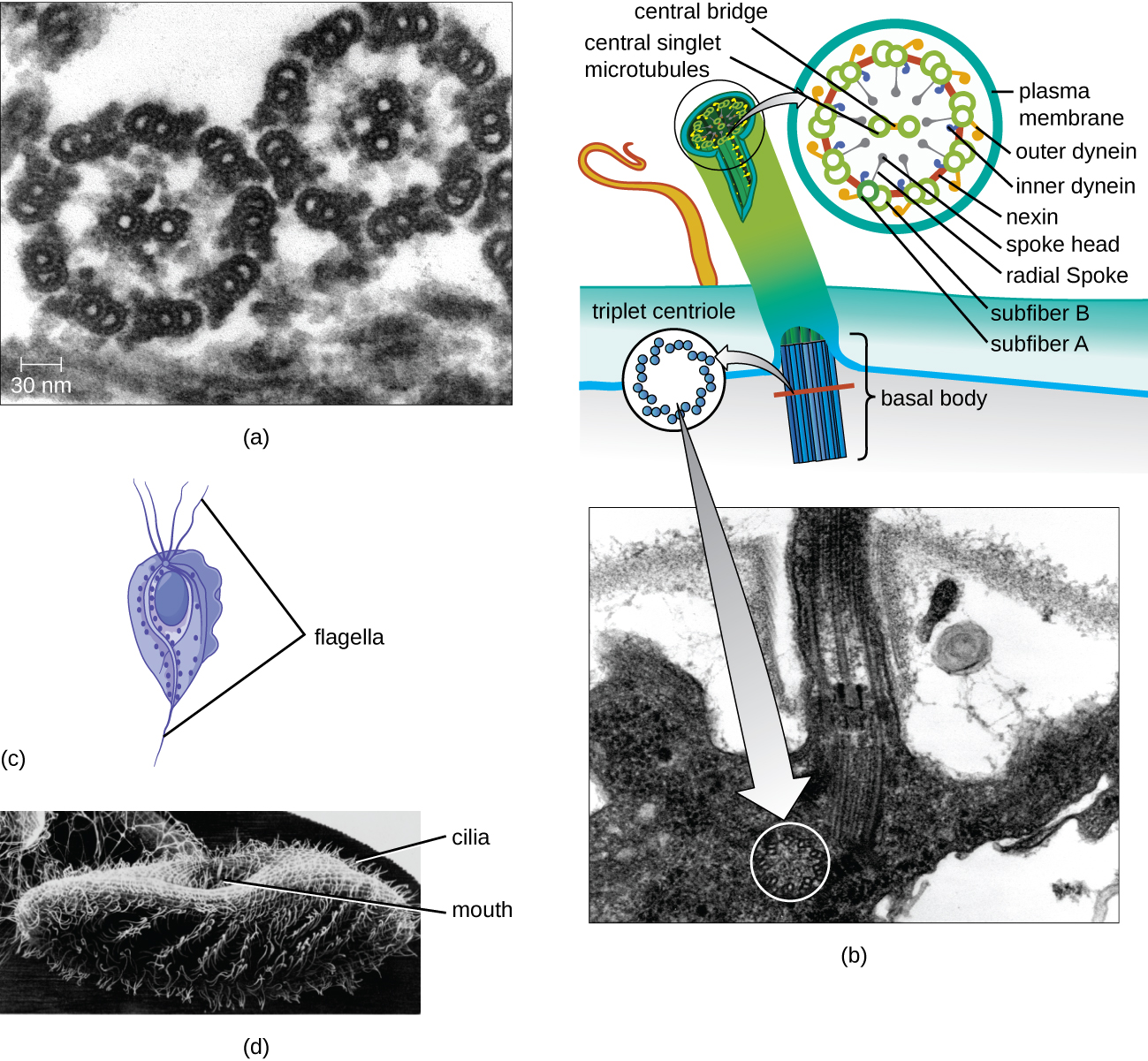
Algumas células eucarióticas usam flagelos para locomoção; no entanto, os flagelos eucarióticos são estruturalmente distintos daqueles encontrados nas células procarióticas. Enquanto o flagelo procariótico é uma estrutura rígida e rotativa, um flagelo eucariótico é mais parecido com um chicote flexível composto por nove pares paralelos de microtúbulos ao redor de um par central de microtúbulos. Esse arranjo é chamado de matriz 9+2 (Figura\(\PageIndex{20}\)). Os microtúbulos paralelos usam proteínas motoras da dineína para se moverem em relação umas às outras, fazendo com que o flagelo se dobre.
Os cílios (singular: cílio) são uma estrutura externa similar encontrada em algumas células eucarióticas. Exclusivos dos eucariotos, os cílios são mais curtos que os flagelos e geralmente cobrem toda a superfície de uma célula; no entanto, eles são estruturalmente semelhantes aos flagelos (uma matriz 9+2 de microtúbulos) e usam o mesmo mecanismo de movimento. Uma estrutura chamada corpo basal é encontrada na base de cada cílio e flagelo. O corpo basal, que liga o cílio ou flagelo à célula, é composto por uma série de microtúbulos trigêmeos semelhantes aos de um centríolo, mas embutidos na membrana plasmática. Por causa de seu comprimento mais curto, os cílios usam um movimento rápido, flexível e ondulado. Além da motilidade, os cílios podem ter outras funções, como varrer partículas passadas ou para dentro das células. Por exemplo, protozoários ciliados usam a varredura dos cílios para mover partículas de alimentos para o aparelho bucal, e as células ciliadas do trato respiratório dos mamíferos batem em sincronia para varrer muco e detritos para cima e para fora dos pulmões (Figura\(\PageIndex{20}\)).
Exercício\(\PageIndex{4}\)
- Explique como o envelope celular das células eucarióticas se compara ao das células procarióticas.
- Explique a diferença entre flagelos eucarióticos e procarióticos.
Foco clínico: Resolução
Como a amoxicilina não resolveu o caso de pneumonia de Barbara, o AP prescreve outro antibiótico, a azitromicina, que tem como alvo os ribossomos bacterianos em vez do peptidoglicano. Depois de tomar a azitromicina conforme as instruções, os sintomas de Barbara desaparecem e ela finalmente começa a se sentir como ela mesma novamente. Presumindo que não houve resistência medicamentosa à amoxicilina e, dada a eficácia da azitromicina, o agente causador da pneumonia de Barbara é provavelmente o Mycoplasma pneumoniae. Embora essa bactéria seja uma célula procariótica, ela não é inibida pela amoxicilina porque não tem parede celular e, portanto, não produz peptidoglicano.
Conceitos principais e resumo
- As células eucarióticas são definidas pela presença de um núcleo contendo o genoma do DNA e ligado por uma membrana nuclear (ou envelope nuclear) composta por duas bicamadas lipídicas que regulam o transporte de materiais para dentro e para fora do núcleo através dos poros nucleares.
- As morfologias das células eucarióticas variam muito e podem ser mantidas por várias estruturas, incluindo o citoesqueleto, a membrana celular e/ou a parede celular.
- O nucléolo, localizado no núcleo das células eucarióticas, é o local da síntese ribossômica e os primeiros estágios da montagem do ribossomo.
- As células eucarióticas contêm ribossomos 80S no retículo endoplasmático rugoso (ribossomos ligados à membrana) e no citoplasma (ribossomos livres). Eles contêm ribossomos dos anos 70 em mitocôndrias e cloroplastos.
- As células eucarióticas desenvolveram um sistema de endomembrana, contendo organelas ligadas à membrana envolvidas no transporte. Isso inclui vesículas, o retículo endoplasmático e o aparelho de Golgi.
- O retículo endoplasmático liso desempenha um papel na biossíntese de lipídios, no metabolismo de carboidratos e na desintoxicação de compostos tóxicos. O retículo endoplasmático rugoso contém ribossomos 80S ligados à membrana que sintetizam proteínas destinadas à membrana celular
- O aparelho de Golgi processa proteínas e lipídios, normalmente por meio da adição de moléculas de açúcar, produzindo glicoproteínas ou glicolipídios, componentes da membrana plasmática que são usados na comunicação célula a célula.
- Os lisossomos contêm enzimas digestivas que decompõem pequenas partículas ingeridas pela endocitose, partículas grandes ou células ingeridas pela fagocitose e componentes intracelulares danificados.
- O citoesqueleto, composto por microfilamentos, filamentos intermediários e microtúbulos, fornece suporte estrutural em células eucarióticas e serve como uma rede para o transporte de materiais intracelulares.
- Os centrossomos são centros organizadores de microtúbulos importantes na formação do fuso mitótico na mitose.
- As mitocôndrias são o local da respiração celular. Eles têm duas membranas: uma externa e uma interna com cristas. A matriz mitocondrial, dentro da membrana interna, contém o DNA mitocondrial, os ribossomos 70S e as enzimas metabólicas.
- A membrana plasmática das células eucarióticas é estruturalmente semelhante à encontrada nas células procarióticas, e os componentes da membrana se movem de acordo com o modelo de mosaico de fluidos. No entanto, as membranas eucarióticas contêm esteróis, que alteram a fluidez da membrana, bem como glicoproteínas e glicolipídios, que ajudam a célula a reconhecer outras células e partículas infecciosas.
- Além do transporte ativo e do transporte passivo, as membranas celulares eucarióticas podem levar material para a célula por meio de endocitose ou expulsar matéria da célula por exocitose.
- Células de fungos, algas, plantas e alguns protistas têm uma parede celular, enquanto as células de animais e alguns protozoários têm uma matriz extracelular pegajosa que fornece suporte estrutural e medeia a sinalização celular.
- Os flagelos eucarióticos são estruturalmente distintos dos flagelos procarióticos, mas têm um propósito semelhante (locomoção). Os cílios são estruturalmente semelhantes aos flagelos eucarióticos, mas mais curtos; eles podem ser usados para locomoção, alimentação ou movimentação de partículas extracelulares.
Notas de pé
- 1 A.E. Barnhill, MT Brewer, S.A. Carlson. “Efeitos adversos dos antimicrobianos por meio da inibição previsível ou idiossincrática dos componentes mitocondriais do hospedeiro.” Agentes antimicrobianos e quimioterapia 56 no. 8 (2012) :4046—4051.
- 2 Fuchs E, Cleveland D.W. “Um andaime estrutural de filamentos intermediários na saúde e na doença.” Ciência 279 nº 5350 (1998) :514—519.
- 3 E. Fuchs, D.W. Cleveland. “Um andaime estrutural de filamentos intermediários na saúde e na doença.” Ciência 279 nº 5350 (1998) :514—519.
- 4 E. Fuchs, D.W. Cleveland. “Um andaime estrutural de filamentos intermediários na saúde e na doença.” Ciência 279 nº 5350 (1998) :514—519.
- 5 N. Yarlett, J. H. P. Hackstein. “Hidrogenossomas: uma organela, várias origens.” BioScience 55 nº 8 (2005) :657—658.
- 6 Sr. Dudzick. “Protistas”. OpenStax CNX. 27 de novembro de 2013. http://cnx.org/contents/f7048bb6-e46...ef291cf7049c@1