
18.4: B 淋巴细胞和抗体
- Page ID
- 200182

学习目标
- 描述 B 细胞的产生和成熟过程
- 比较 B 细胞受体和 T 细胞受体的结构
- 比较 B 细胞的 T 依赖性和非依赖性 T 的激活
- 比较初级抗体和二级抗体反应
体液免疫是指由B淋巴细胞或B细胞分泌的抗体介导的适应性免疫防御机制。 本节将重点介绍 B 细胞,讨论它们的产生和成熟、受体和激活机制。
B 细胞的产生和成熟
与T细胞一样,B细胞由骨髓中的多能造血干细胞(HSC)形成,沿着淋巴样干细胞和淋巴细胞的途径行驶(见图 17.3.1)。 但是,与 T 细胞不同,注定成为 B 细胞的淋巴细胞不会离开骨髓进入胸腺进行成熟。 相反,最终的B细胞继续在骨髓中成熟。
B 细胞成熟的第一步是评估其抗原结合受体的功能。 这是通过对具有正常功能受体的B细胞进行阳性选择来实现的。 然后使用阴性选择机制来消除自我反应的B细胞并最大限度地降低自身免疫的风险。 否定选择自反应 B 细胞可能涉及通过细胞凋亡、编辑或修饰受体使其不再具有自反应或诱导 B 细胞中的能量来消除。 通过骨髓选择的未成熟B细胞然后进入脾脏进入成熟的最后阶段。 在那里,它们变成了天真的成熟B细胞,即尚未被激活的成熟B细胞。
练习\(\PageIndex{1}\)
将 B 细胞的成熟与 T 细胞的成熟进行比较。
B 细胞受体
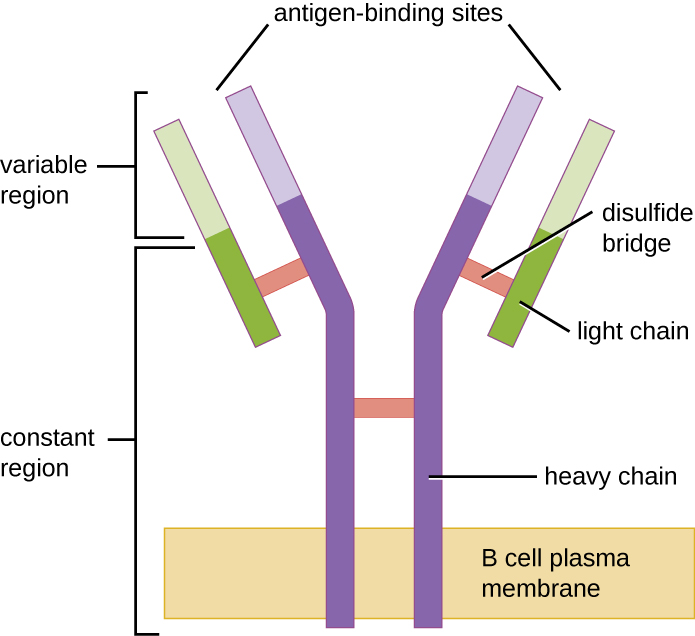
像 T 细胞一样,B 细胞拥有具有不同特异性的抗原特异性受体。 尽管它们依靠T细胞来获得最佳功能,但B细胞可以在没有T细胞帮助的情况下被激活。 幼稚成熟 B 细胞的 B 细胞受体 (BCR) 是 IgD 和 IgM 的膜结合单体形式。 它们有两条相同的重链和两条相同的轻链,通过二硫键连接成基本的 “Y” 形(图\(\PageIndex{1}\))。 Y形分子的树干是两条重链的恒定区域,横跨B细胞膜。 暴露于B细胞外部的两个抗原结合位点参与特定病原表位的结合,从而启动激活过程。 据估计,每个幼稚的成熟B细胞在其膜上有超过100,000个BCR,并且每个BCR具有相同的表位结合特异性。
为了做好对各种微生物表位反应的准备,B 细胞,如 T 细胞,使用数百个基因片段的遗传重排来提供必要的受体特异性多样性。 BCR 重链的可变区域由 V、D 和 J 段组成,类似于 TCR 的 β 链。 BCR 轻链的可变区域由 V 和 J 段组成,类似于 TCR 的 α 链。 所有可能的V-J-D(重链)和V-J(轻链)组合的基因重排为BCR和激活后分泌的抗体提供了数百万个独特的抗原结合位点。
BCR 和 TCR 之间的一个重要区别是它们与抗原表位相互作用的方式。 虽然 TCR 只能与 MHC I 或 MHC II 的抗原结合裂缝内存在的抗原表位相互作用,而 BCR 不要求与 MHC 呈现抗原;它们可以与游离抗原的表位或完整病原体表面显示的抗原表位相互作用。 另一个重要的区别是 TCR 只能识别蛋白质表位,而 BCR 可以识别与不同分子类别(例如蛋白质、多糖、脂多糖)相关的表位。
B 细胞的激活是通过不同的机制发生的,具体取决于抗原的分子类别。 蛋白质抗原激活 B 细胞需要 B 细胞起到 APC 的作用,将具有 MHC II 的蛋白质表位呈现给辅助 T 细胞。 由于蛋白质抗原依赖于 T 细胞来激活 B 细胞,因此被归类为 T 依赖性抗原。 相比之下,多糖、脂多糖和其他非蛋白抗原被认为是与T无关的抗原,因为它们可以在不进行抗原处理和呈现给T细胞的情况下激活B细胞。
练习\(\PageIndex{2}\)
- 什么类型的分子充当 BCR?
- TCR 和 BCR 在抗原识别方面有什么区别?
- 哪些分子类别是 T 依赖性抗原,哪些是 T 依赖性抗原?
不依赖于 T 细胞的 B 细胞激活
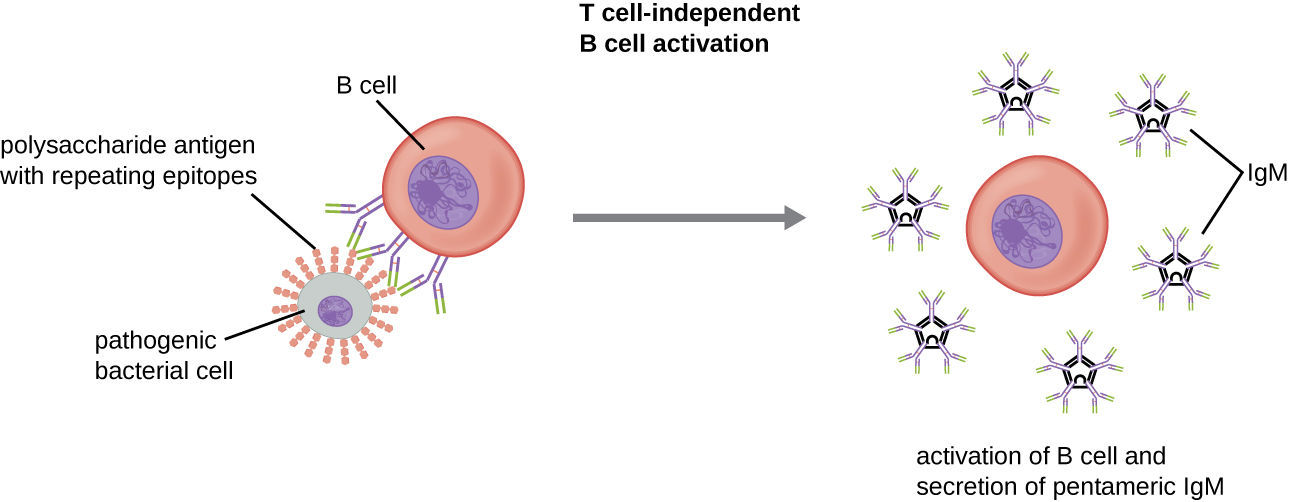
在没有辅助性 T 细胞合作的情况下激活 B 细胞被称为 T 细胞依赖性激活,发生在 BCR 与 T 非依赖性抗原相互作用时。 不依赖于 T 的抗原(例如多糖胶囊、脂多糖)在其结构中具有重复的表位单元,这种重复允许多个 BCR 交联,从而提供第一个激活信号(图\(\PageIndex{2}\))。 由于 T 细胞不参与,第二个信号必须来自其他来源,例如 toll 样受体与 PAMP 的相互作用或与补体系统因子的相互作用。
一旦B细胞被激活,它就会进行克隆增殖,子细胞分化为浆细胞。 浆细胞是分泌大量抗体的抗体工厂。 分化后,表面 BCR 消失,浆细胞分泌出与 BCR 具有相同抗原特异性的五聚体 IgM 分子(图\(\PageIndex{2}\))。
与 T 细胞无关的反应是短暂的,不会产生记忆 B 细胞。 因此,它不会对随后暴露于 T 非依赖性抗原产生次要反应。
练习\(\PageIndex{3}\)
- 独立于 T 细胞的激活 B 细胞需要哪两个信号?
- 浆细胞的功能是什么?
T 细胞依赖性激活 B 细胞
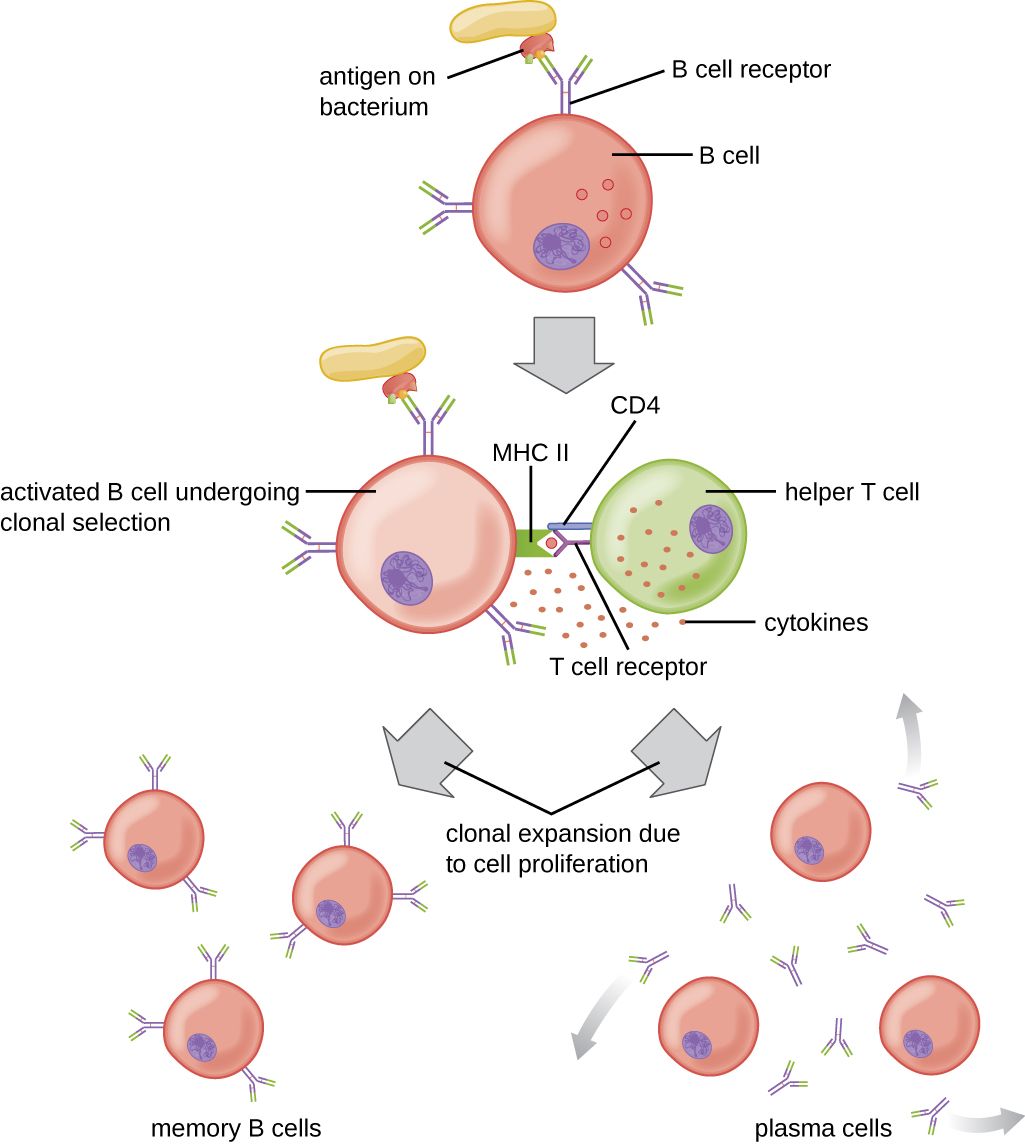
依赖于 T 细胞的 B 细胞激活比不依赖于 T 细胞的激活更为复杂,但由此产生的免疫反应更强,会产生记忆力。 T 细胞依赖性激活既可以是对游离蛋白抗原的反应,也可以是对与完整病原体相关的蛋白质抗原的反应。 天真成熟 B 细胞上的 BCR 与游离蛋白抗原之间的相互作用刺激了抗原的内化,而与与完整病原体相关的抗原的相互作用则在内化之前启动从病原体中提取抗原。 一旦内化在 B 细胞内,蛋白质抗原就会被处理并呈现 MHC II。 然后,对同一抗原具有特异性的辅助T细胞识别所呈现的抗原。 辅助性 T 细胞的 TCR 识别外来抗原,而 T 细胞的 CD4 分子与 B 细胞上的 MHC II 相互作用。 对同一抗原具有特异性的 B 细胞和辅助性 T 细胞之间的协调被称为关联识别。
一旦通过关联识别激活,T H 2 细胞就会产生并分泌细胞因子,激活 B 细胞并导致增殖为克隆子细胞。 经过几轮增殖后,T H 2 细胞提供的额外细胞因子刺激活的 B 细胞克隆分化为记忆 B 细胞,记忆 B 细胞将在随后暴露于相同蛋白质表位时迅速做出反应,而浆细胞则会失去膜 BCR 和最初分泌五聚体 IgM(图\(\PageIndex{3}\))。
初次分泌 IgM 后,T H 2 细胞分泌的细胞因子刺激浆细胞从 IgM 的产生转变为 IgG、IgA 或 IgE 的产生。 这种过程称为类别切换或同型切换,允许从同一个活化的 B 细胞中克隆的浆细胞产生具有相同表位特异性的各种抗体类别。 类别切换是通过对编码恒定区域的基因片段进行遗传重排来实现的,恒定区域决定了抗体的类别。 可变区域没有改变,因此新的抗体类别保留了原始抗原表位的特异性。
练习\(\PageIndex{4}\)
- 依赖 T 细胞激活 B 细胞需要哪些步骤?
- 什么是抗体类别转换?为什么它很重要?
主要和次要回应
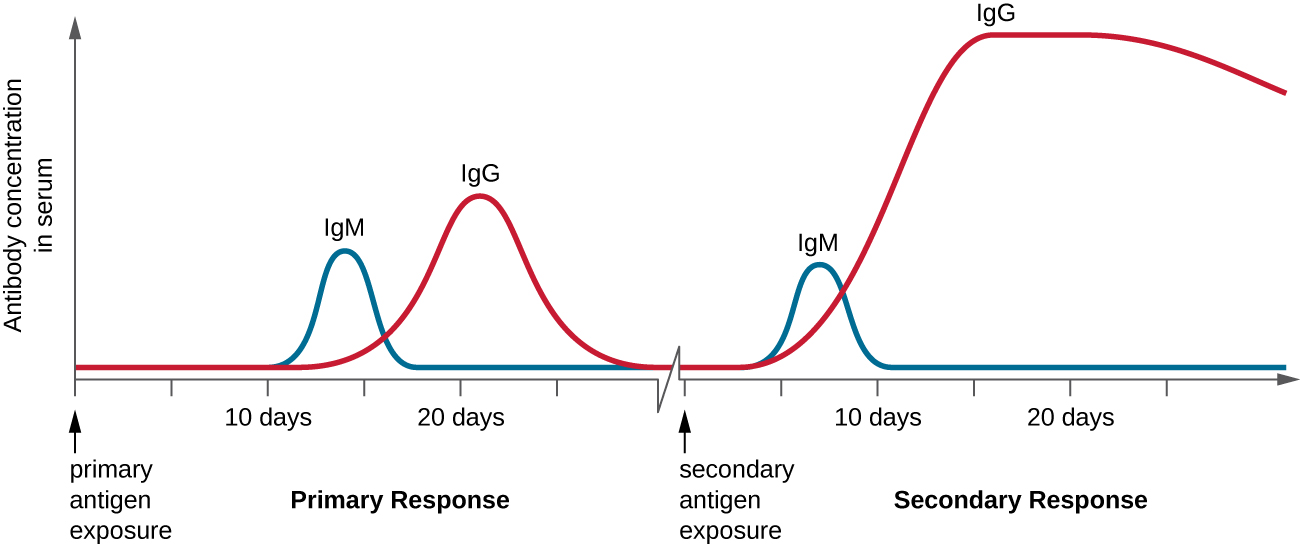
B 细胞依赖性激活 T 细胞在与适应性免疫相关的原发和次要反应中都起着重要作用。 首次接触蛋白质抗原时,会出现依赖于 T 细胞的一级抗体反应。 初始反应的初始阶段是大约10天的滞后期或潜伏期,在此期间血清中无法检测到抗体。 这个滞后期是初级反应的所有步骤所需的时间,包括抗原与BCR的天真成熟B细胞结合、抗原处理和表现、辅助T细胞激活、B细胞激活和克隆增殖。 滞后期结束的特征是血清中 IgM 水平升高,因为 T H 2 细胞刺激 B 细胞分化为浆细胞。 IgM 水平在初级抗原暴露后大约 14 天达到峰值;大约在同一时间,T H 2 刺激抗体类别转换,血清中的 IgM 水平开始下降。 同时,IgG 水平会增加,直到在初级反应大约三周后达到峰值(图\(\PageIndex{4}\))。
在初级反应期间,一些克隆的B细胞被分化为存储器B细胞,编程为对随后的暴露做出反应。 这种次要响应的发生速度比主要响应更快、更有力。 滞后期缩短到只有几天,IgG的产量明显高于初级反应中观察到的产量(图\(\PageIndex{4}\))。 此外,二次反应期间产生的抗体更有效,与靶向表位结合的亲和力更高。 继发反应期间产生的浆细胞比初级反应期间产生的浆细胞寿命更长,因此特异性抗体水平在更长的时间内保持较高的水平。
练习\(\PageIndex{5}\)
- 在一级抗体反应的滞后期会发生哪些事件?
- 为什么在二级抗体反应期间抗体水平保持较高的时间?
关键概念和摘要
- B 淋巴细胞或 B 细胞产生与体液免疫有关的抗体。 B 细胞在骨髓中产生,成熟的初始阶段发生在骨髓中,然后传播到脾脏,进入最后的成熟步骤,形成天真的成熟 B 细胞。
- B 细胞受体 (BCR) 是 IgD 和 IgM 的膜结合单体形式,可将特定的抗原表位与其 Fab 抗原结合区结合。 抗原结合特异性的多样性是由V、D和J片段的遗传重排产生的,类似于用于TCR多样性的机制。
- 蛋白质抗原被称为 T 依赖性抗原,因为它们只能在辅助性 T 细胞的合作下激活 B 细胞。 其他分子类别不需要 T 细胞合作,被称为 T 无关抗原。
- B 细胞的非@@ 依赖性 T 细胞的激活涉及重复的非蛋白抗原表位对 BCR 进行交联。 它的特点是浆细胞产生 IgM,不产生记忆 B 细胞。
- B 细胞依赖性 T 细胞的激活包括处理蛋白质抗原并将其呈现给辅助 T 细胞,通过活化的 T H 2 细胞分泌的细胞因子激活 B 细胞,以及由于 Cl ass 产生不同类别抗体的浆细胞切换。 还会产生@@ 记忆 B 细胞。
- 二次暴露于 T 依赖性抗原会导致记忆 B 细胞引发的二级抗体反应。 二次反应发展得更快,产生更高、更持续的抗体水平,对特异性抗原具有更高的亲和力。