
18.3: T 淋巴细胞
- Page ID
- 200166

学习目标
- 描述 T 细胞成熟和胸腺选择的过程
- 解释导致 T 细胞受体多样性的遗传事件
- 比较和对比 T 细胞在激活和功能方面的各种类别和亚型
- 解释超抗原影响不受控制的 T 细胞激活的机制
正如《特异性适应性免疫概述》中所解释的那样,参与体液免疫的抗体通常在病原体和毒素附着和侵入宿主细胞之前将其结合。 因此,体液免疫主要涉及对抗细胞外空间中的病原体。 但是,已经进入宿主细胞的病原体在很大程度上受到保护,免受体液抗体介导的防御的影响。 另一方面,细胞免疫通过T淋巴细胞或T细胞的作用靶向和消除细胞内病原体(图\(\PageIndex{1}\))。 T 细胞在协调整体适应性免疫反应(体液和细胞)以及先天免疫的细胞防御方面也起着更为重要的作用。

T 细胞的产生和成熟
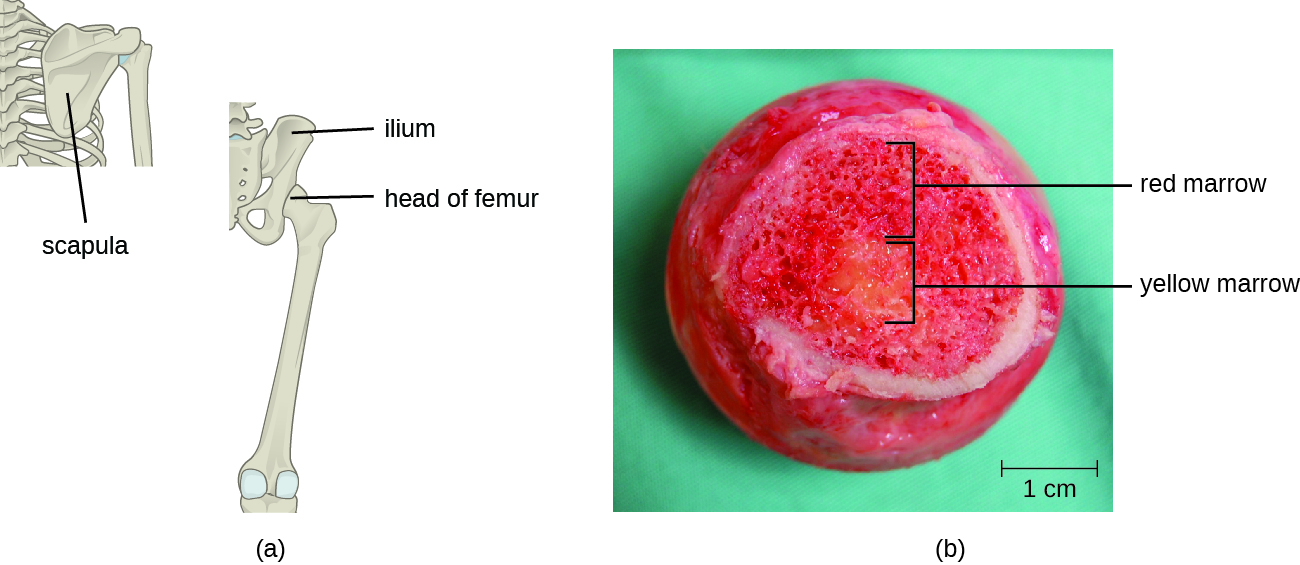
与所有其他参与先天免疫和适应性免疫的白细胞一样,T细胞是由骨髓中的多能造血干细胞(HSC)形成的(见图17.3.1)。 但是,与具有先天免疫力的白细胞不同,最终的T细胞首先分化为淋巴样干细胞,然后变成小的、未成熟的淋巴细胞,有时也称为淋巴细胞。 分化的第一步发生在骨髓的红骨髓中(图\(\PageIndex{2}\)),之后未成熟的 T 淋巴细胞进入血液并进入胸腺进行最后的成熟步骤(图\(\PageIndex{3}\))。 一旦进入胸腺,未成熟的 T 淋巴细胞就被称为胸腺细胞。
胸腺细胞在胸腺中的成熟可以分为三个关键步骤,即正向选择和负向选择,统称为胸腺选择。 胸腺选择的第一步发生在胸腺皮层,涉及APC激活所需的功能性T细胞受体(TCR)的开发。 通过诱导细胞凋亡(程序性控制细胞死亡),通过阴性选择去除具有缺陷的胸腺细胞。 胸腺选择的第二步也发生在皮层中,涉及积极选择能够与MHC分子适当相互作用的胸腺细胞。 能够与 MHC 分子适当相互作用的胸腺细胞会受到正向刺激,使它们进一步进入成熟过程,而相互作用不当的胸腺细胞不会受到刺激,会被细胞凋亡消除。 胸腺选择的第三步也是最后一步发生在皮层和延髓中,涉及阴性选择,通过细胞凋亡去除自我反应的胸腺细胞,即那些对自身抗原有反应的胸腺细胞。 最后一步有时被称为中枢耐受,因为它会阻止自我反应的T细胞进入血液,并可能导致自身免疫性疾病,这种疾病发生在免疫系统攻击健康的 “自我” 细胞时。
尽管具有中枢耐受性,但一些自反应 T 细胞通常会逃离胸腺进入外周血液。 因此,需要第二道称为外周耐受的防线来预防自身免疫性疾病。 外周耐受性涉及调节性 T 细胞对自反应 T 细胞的过敏和抑制机制。 无能是指对抗原刺激无反应的状态。 对于逃离胸腺的自反应 T 细胞,缺乏激活所需的基本共刺激信号会导致过敏并阻止自身免疫激活。 调节性 T 细胞通过抑制自反应 T 细胞的激活和功能以及分泌抗炎细胞因子来参与外周耐受。
目前尚不完全了解哪些事件特别引导胸腺细胞成熟为调节性 T 细胞。 目前的理论表明,关键事件可能发生在胸腺选择的第三步中,即大多数自反应性T细胞被消除。 调节性 T 细胞可能会收到一个独特的信号,该信号低于靶向它们进行阴性选择和凋亡所需的阈值。 因此,这些细胞继续成熟,然后离开胸腺,武装起来,抑制自反应 T 细胞的激活。
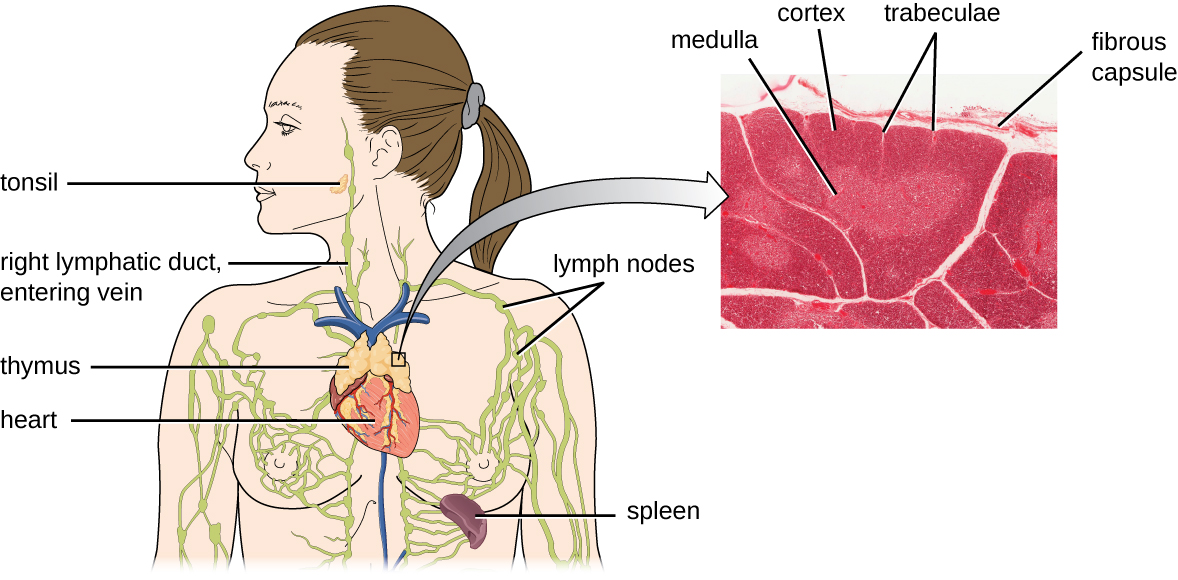
据估计,胸腺选择的三个步骤可消除 98% 的胸腺细胞。 其余 2% 的胸腺通过血液和淋巴系统迁移到次生淋巴器官/组织部位,例如淋巴结、脾脏和扁桃体(图\(\PageIndex{3}\)),等待装甲运兵车呈现特定抗原激活。 在它们被激活之前,它们被称为成熟的天真 T 细胞。
练习\(\PageIndex{1}\)
- 哪些解剖部位参与了 T 细胞的产生和成熟?
- 胸腺选择涉及哪三个步骤?
- 为什么中心容差和周边容差很重要? 它们防止了什么?
T 细胞的类别
T 细胞可以分为三个不同的类别:辅助性 T 细胞、调节性 T 细胞和细胞毒性 T 细胞。 这些类别是根据其某些表面分子的表达、激活模式及其在适应性免疫中的功能作用来区分的(表\(\PageIndex{1}\))。
所有 T 细胞都会产生分化簇 (CD) 分子,即细胞表面糖蛋白,可用于识别和区分各种类型的白细胞。 尽管 T 细胞可以产生各种各样的 CD 分子,但 CD4 和 CD8 是用于分化这些类别的两个最重要的分子。 辅助性T细胞和调节性T细胞的特征是其表面CD4的表达,而细胞毒性T细胞的特征是CD8的表达。
T细胞的类别也可以通过它们相互作用以激活的特定的MHC分子和APC来区分。 辅助性T细胞和调节性T细胞只能由呈现与MHC II相关的抗原的APC激活。 相比之下,细胞毒性 T 细胞识别与 MHC I 相关的抗原,无论是由 APC 还是感染了细胞内病原体的有核细胞。
不同类别的T细胞在免疫系统中也起着不同的功能作用。 辅助性T细胞充当中央协调器,帮助激活和指导体液和细胞免疫的功能。 此外,辅助性T细胞增强了巨噬细胞和先天免疫的NK细胞的致病杀灭功能。 相比之下,调节性T细胞的主要作用是防止不良和潜在的破坏性免疫反应。 例如,如前所述,它们在外周耐受性中的作用可以预防自身免疫性疾病。 最后,细胞毒性 T 细胞是细胞免疫的主要效应细胞。 它们识别并靶向被细胞内病原体感染的细胞,破坏受感染的细胞以及内部的病原体。
| 班级 | 表面 CD 分子 | 激活 | 函数 |
|---|---|---|---|
| 辅助性 T 细胞 | CD4 | 呈现与 MHC II 相关的抗原的装甲运兵车 | 协调体液和细胞免疫 |
| 参与巨噬细胞和 NK 细胞的激活 | |||
| 调节性 T 细胞 | CD4 | 呈现与 MHC II 相关的抗原的装甲运兵车 | 参与外周耐受和自身免疫反应的预防 |
| 细胞毒性 T 细胞 | CD8 | 呈现与 MHC I 相关的抗原的 APC 或受感染的有核细胞 | 摧毁被细胞内病原体感染的细胞 |
练习\(\PageIndex{2}\)
- 这三类 T 细胞的独特功能是什么?
- 除了 APC 以外的细胞所呈现的抗原可以激活哪些 T 细胞?
T 细胞受体
对于辅助性T细胞和细胞毒性T细胞来说,激活是一个复杂的过程,需要多个分子相互作用并暴露于细胞因子。 在激活过程中,T 细胞受体 (TCR) 参与病原体表位识别的第一步。
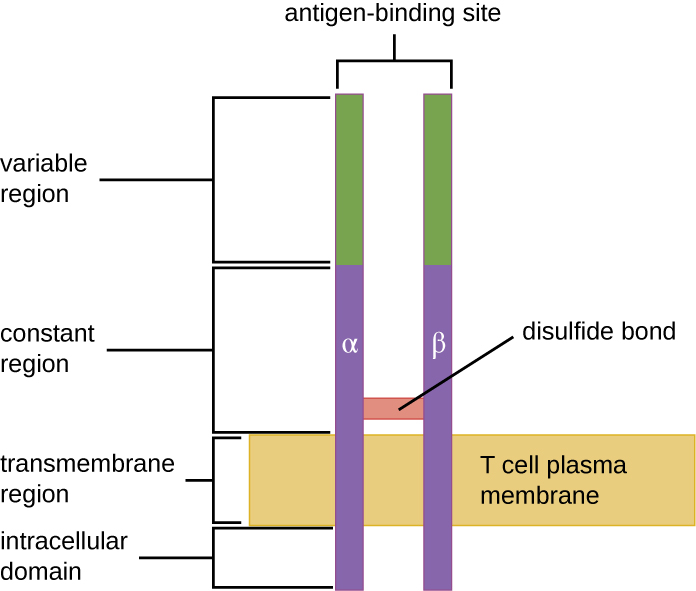
TCR 与抗体 IgD 和 IgM(B 细胞膜表面的抗原受体)来自同一个受体家族,因此具有共同的结构元素。 与抗体类似,TCR 具有可变区域和恒定区域,可变区域提供抗原结合位点(图\(\PageIndex{4}\))。 但是,与免疫球蛋白分子相比,TCR的结构更小,也不那么复杂(图18.1.4)。 免疫球蛋白有四条肽链和Y形结构,而TCR仅由两条肽链(α和β链)组成,两者均横跨T细胞的细胞质膜。
TCR 具有表位特异性,据估计,需要有 2,500 万个具有独特表位结合的 TCR 的 T 细胞来保护个体免受各种微生物病原体的侵害。 由于人类基因组仅包含大约 25,000 个基因,因此我们知道每个特定的 TCR 都无法由其自己的基因组编码。 这就提出了一个问题,那就是如何实现拥有数百万个特定 TCR 的如此庞大的 T 细胞群体。 答案是一个称为遗传重排的过程,在胸腺选择的第一步中,它发生在胸腺中。
编码 TCR 可变区域的基因分为不同的基因片段,分别称为变量 (V)、多样性 (D) 和连接 (J) 片段。 与 TCR 的 α 链相关的基因片段由 70 个或更多不同的 V α 片段和 61 个不同的 J α 片段组成。 与 TCR 的 β 链相关的基因片段由 52 个不同的 V β 片段、两个不同的 D β 片段和 13 个不同的 J β 片段组成。 在胸腺功能性 TCR 发育过程中,T 细胞中的遗传重排将一个 V α 片段和一个 J α 片段聚集在一起,编码 α 链的可变区域。 同样,遗传重排将其中一个 V β 片段与一个 D β 片段和一个 TJ β 片段组合在一起,编码 β 链的可变区域。 V、D 和 J 不同片段之间所有可能的重排组合提供了产生数百万个具有独特表位特异性可变区域的 TCR 所需的遗传多样性。
练习\(\PageIndex{3}\)
- TCR 和免疫球蛋白有什么相似之处和区别?
- 使用什么工艺来提供数百万个独特的 TCR 结合位点?
辅助性T细胞的激活和分化
辅助性T细胞只能由具有与MHC II相关的经过处理的外来表位的APC激活。 激活过程的第一步是 TCR 识别 MHC II 抗原结合裂缝中存在的特定外来表位。 第二步涉及辅助性T细胞上的CD4与MHC II分子中与抗原结合裂缝分开的区域的相互作用。 第二次相互作用巩固了MHC II-TCR复合物,并确保辅助T细胞同时识别APC的外来(“非自身”)表位和 “自身” 抗原;这两种识别都是激活细胞所必需的。 在第三步中,APC 和 T 细胞分泌激活辅助性 T 细胞的细胞因子。 然后,激活的辅助 T 细胞会增殖,除以有丝分裂,产生克隆性天真的辅助 T 细胞,分化为具有不同功能的亚型(图\(\PageIndex{5}\))。
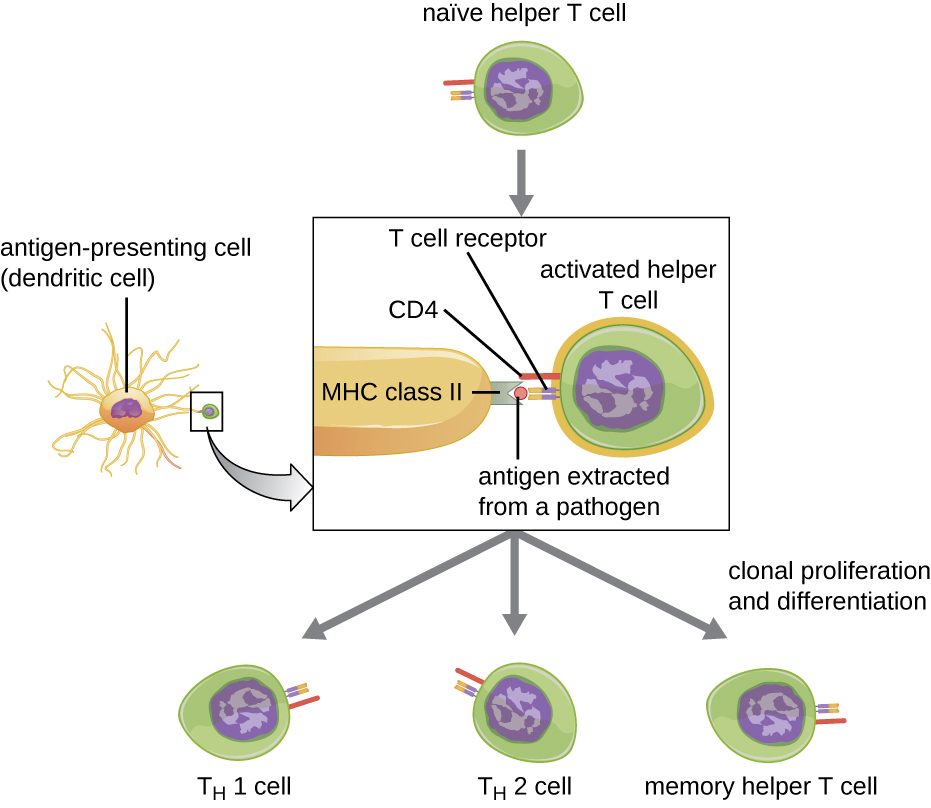
活化的辅助性T细胞可以分化为四种不同的亚型之一,总结见表\(\PageIndex{2}\)。 分化过程由 APC 分泌的细胞因子引导。 根据哪些 APC 分泌的细胞因子与激活的辅助 T 细胞相互作用,该细胞可能会分化为 T 辅助细胞 1(T H 1)、T 辅助细胞 2(T H 2)或记忆辅助细胞。 这两种类型的辅助性T细胞是寿命相对较短的效应细胞,这意味着它们具有即时免疫反应的各种功能。 相比之下,记忆辅助性T细胞的寿命相对较长;它们被编程为 “记住” 特定的抗原或表位,以便对随后的暴露产生快速、强烈的二次反应。
T H 1 细胞分泌自己的细胞因子,这些因子参与刺激和协调参与适应性和先天免疫的其他细胞。 例如,它们刺激细胞毒性 T 细胞,增强其对受感染细胞的杀伤力并促进分化为记忆细胞毒性 T 细胞。 T H 1 细胞还刺激巨噬细胞和中性粒细胞在杀死细胞内细菌方面变得更有效。 它们还可以刺激 NK 细胞变得更有效地杀死靶细胞。
T H 2 细胞通过分泌激活 B 细胞、指导 B 细胞分化和抗体产生的细胞因子,在协调体液免疫反应方面起着重要作用。 T H 2 细胞产生的各种细胞因子协调抗体类别切换,这使得 B 细胞能够根据需要在产生 IgM、IgG、IgA 和 IgE 之间切换,以发挥特异性抗体功能和提供病原体特异性体液免疫反应。
通过观察发现了对某些感染的免疫与 T H 1 或 T H 2 细胞无关的第三种辅助性 T 细胞亚型,即 T H 17 细胞。 T H 17 细胞及其产生的细胞因子似乎是人体抵御慢性皮肤粘膜感染的特殊原因。 粘膜中缺乏足够的 T H 17 细胞的患者(例如 HIV 患者)可能更容易受到菌血症和胃肠道感染的影响。 1
| 子类型 | 函数 |
|---|---|
| T H 1 细胞 | 刺激细胞毒性 T 细胞并产生记忆细胞毒性 T 细胞 |
| 刺激巨噬细胞和中性粒细胞 (PMN) 以更有效地在细胞内杀死病原体 | |
| 刺激 NK 细胞更有效地杀死 | |
| T H 2 细胞 | 刺激 B 细胞活化并分化为浆细胞和记忆 B 细胞 |
| B 细胞中的直接抗体类别切换 | |
| T H 17 个细胞 | 刺激对特定感染(例如慢性皮肤粘膜感染)的免疫力 |
| 记忆助手 T 细胞 | “记住” 一种特定的病原体,并在再次暴露时产生强烈、快速的二次反应 |
细胞毒性 T 细胞的激活和分化
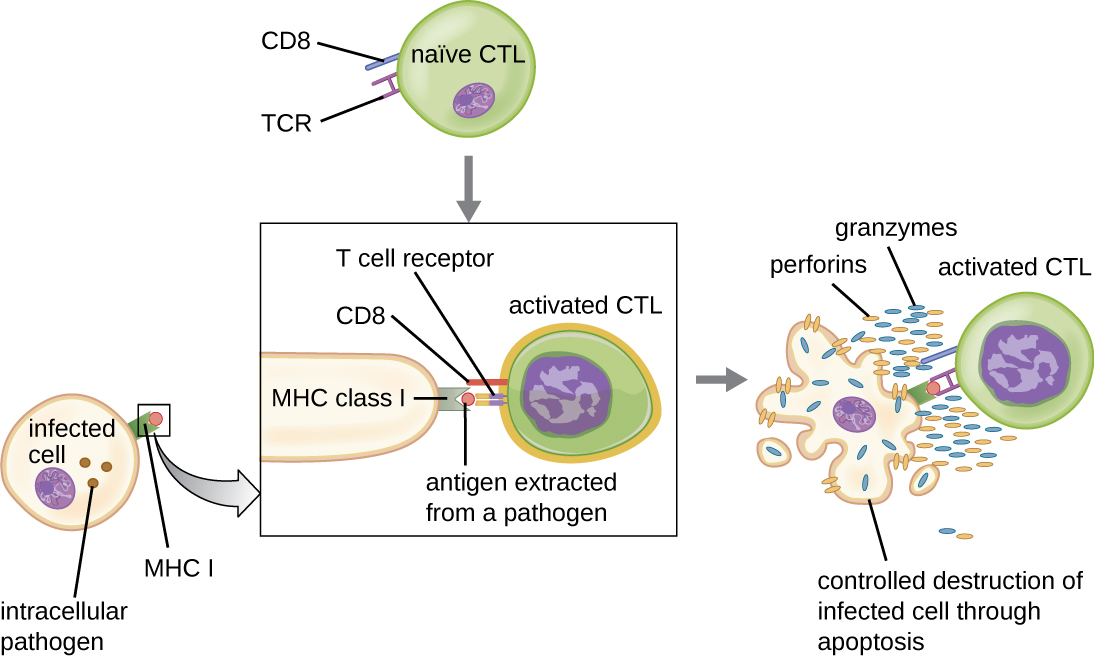
细胞毒性 T 细胞(也称为细胞毒性 T 淋巴细胞,或 CTL)由 APC 通过类似于辅助性 T 细胞的三步过程激活。 关键区别在于,细胞毒性T细胞的激活涉及识别以MHC I(而不是MHC II)呈现的抗原以及CD8(而不是CD4)与受体复合物的相互作用。 成功共识别外源表位和自身抗原后,APC 和细胞毒性 T 细胞产生的细胞因子会激活克隆增殖和分化。 活化的细胞毒性 T 细胞可以分化为靶向病原体进行破坏的效应细胞毒性 T 细胞或准备对后续暴露做出反应的记忆细胞。
如前所述,细胞毒性 T 细胞的增殖和分化也受到由相同外来表位激活的 T H 1 细胞分泌的细胞因子的刺激。 来自这些 T H 1 细胞的共同刺激是由分泌的细胞因子提供的。 尽管在没有 T H 1 细胞刺激的情况下激活细胞毒性 T 细胞是可能的,但这种激活并不那么有效或持久。
一旦激活,细胞毒性 T 细胞就会成为细胞免疫的效应细胞,通过与 NK 细胞非常相似的机制识别和杀死感染了细胞内病原体的细胞。 但是,虽然 NK 细胞识别细胞应激或异常的非特异性信号,但细胞毒性 T 细胞通过抗原呈现与 MHC I 相关的病原特异性表位来识别受感染细胞。一旦识别出受感染细胞,细胞毒性 T 细胞的 TCR 就会与表位结合并释放破坏受感染细胞的穿孔素和颗粒酶(图\(\PageIndex{6}\))。 Perforin 是一种在靶细胞中产生毛孔的蛋白质,而 granzymes 是进入毛孔并诱导细胞凋亡的蛋白酶。 这种程序性细胞死亡机制是一种可控且有效的手段,可以在不释放内部的病原体感染邻近细胞的情况下摧毁和移除受感染细胞,就像仅仅裂解受感染的细胞一样。
在这段视频中,你可以看到一个细胞毒性 T 细胞在靶细胞中诱导细胞凋亡。
练习\(\PageIndex{4}\)
- 比较和对比辅助性T细胞和细胞毒性T细胞的激活情况。
- 辅助性T细胞亚型有哪些不同功能?
- CTL 介导破坏受感染细胞的机制是什么?
超抗原和不受控制的 T 细胞激活
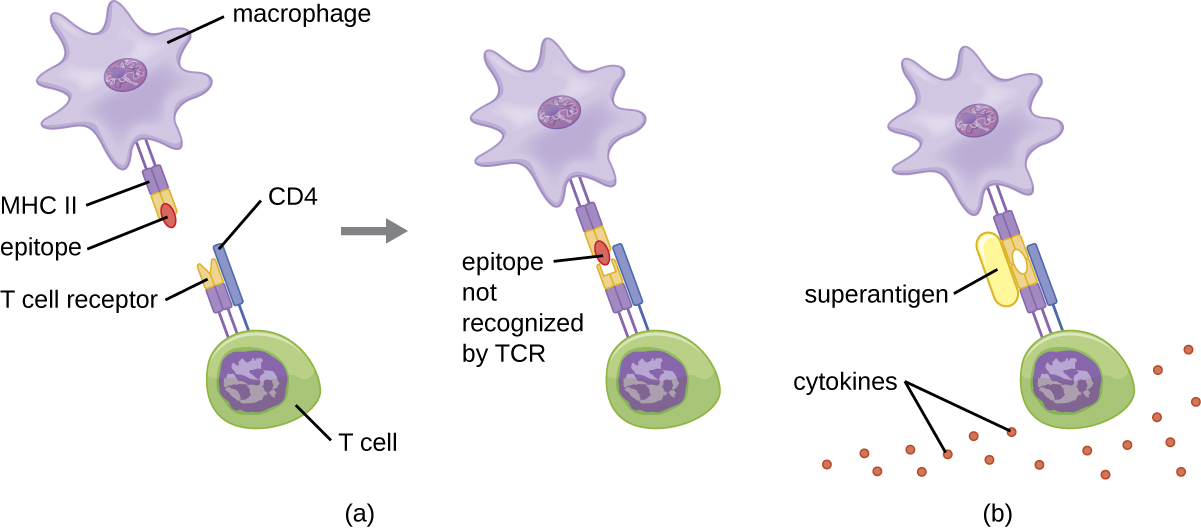
当控制和调节 T 细胞活化时,结果是有效对抗感染的保护性反应。 但是,如果 T 细胞激活不受管制且过度激活,结果可能会危及生命。 某些细菌和病毒病原体会产生被称为超抗原的毒素(参见细菌和病毒病原体的毒力因子),这些毒素可以引发这种不受管制的反应。 已知的细菌超抗原包括毒性休克综合征毒素 (TSST)、葡萄球菌肠毒素、链球菌热原毒素、链球菌超抗原和链球菌有丝分裂外毒素。 已知会产生超抗原的病毒包括爱泼斯坦-巴尔病毒(人类疱疹病毒 4)、巨细胞病毒(人类疱疹病毒 5)等。
超抗原激活 T 细胞的机制涉及它们与 APC 的 MHC II 分子和 TCR β 链的可变区域同时结合。 这种结合发生在MHC II的抗原结合裂缝之外,因此超抗原将在不识别特异性外来表位的情况下桥接在一起并激活MHC II和TCR(图\(\PageIndex{7}\))。 结果是细胞因子过量、不受控制地释放,通常称为细胞因子风暴,这会刺激过度的炎症反应。 这可能导致危险的血压降低、休克、多器官衰竭,甚至可能导致死亡。
练习\(\PageIndex{5}\)
- 超抗原的例子有哪些?
- 超抗原如何激活辅助性T细胞?
- 超抗原对T细胞有什么影响?
一个很好的例子:超抗原
梅利莎(Melissa)是一位原本健康的22岁女性,被她关心的男友带到急诊室。 她抱怨突然出现高烧、呕吐、腹泻和肌肉酸痛。 在初次采访中,她告诉主治医生,她正在接受荷尔蒙节育,而且已经进入周期的月经部分两天了。 她没有服用其他药物,也没有滥用任何药物或酒精。 她不是吸烟者。 她没有糖尿病,据她所知,目前没有任何形式的感染。
在急诊室等候时,梅利莎的血压开始急剧下降,精神状态恶化为普遍混乱。 医生认为她可能患有中毒性休克综合症(TSS)。 TSS 由毒素 TSST-1 引起,这是一种与金黄色葡萄球菌相关的超抗原,卫生棉条使用不当是导致 TSS 的常见感染原因。 超抗原不恰当地刺激了广泛的 T 细胞激活和细胞因子的过度释放,从而导致大规模的全身性炎症反应,可能致命。
可以用阴道或宫颈拭子来确认微生物的存在,但根据梅利莎的症状和病史,这些检查对进行并不重要。 医生开出补液、支持疗法和抗生素来阻止细菌感染。 她还开了增加梅利莎血压的药物。 梅利莎在医院待了三天接受治疗;此外,由于与TSS相关的肾衰竭风险很高,她的肾功能受到监测。 72 小时后,梅利莎身体健康,可以出院,继续在家中康复。
练习\(\PageIndex{6}\)
抗生素疗法会以什么方式帮助对抗超抗原?
临床重点:第 2 部分
奥利维亚的淋巴结、腹部和脾脏肿大表明对正在进行的全身感染有强烈的免疫反应。 此外,小奥利维亚不愿转过头,似乎正在经历严重的颈部疼痛。 医生下令进行全血细胞计数、血液培养和腰椎穿刺。 获得的脑脊液(CSF)呈现混浊状态,需要通过革兰染色评估和培养来进一步评估潜在的细菌病原体。 全血细胞计数表明奥利维亚血液中的白细胞数量增加。 据记录,白细胞增高为28.5 K/µL(正常范围:6.0—17.5 K/µL)。 中性粒细胞百分比记录为60%(正常范围:23—45%)。 脑脊液中的葡萄糖水平记录为30 mg/100 mL(正常范围:50—80 mg/100 mL)。 脑脊液中的白细胞计数为1,163/mm3(正常范围:5—20/mm 3)。
练习\(\PageIndex{7}\)
- 根据这些结果,你有初步的诊断吗?
- 根据这一初步诊断,推荐的治疗方法是什么?
关键概念和摘要
- 未成熟的 T 淋巴细胞在红骨髓中产生,并传播到胸腺进行成熟。
- 胸腺选择是阴性选择和阳性选择的三步过程,它决定了哪些 T 细胞将成熟并从胸腺流入外周血液。
- 中心耐受性涉及对胸腺中自反应 T 细胞的负面选择,而外周耐受则涉及防止自反应免疫反应和自身免疫的过敏和调节 T 细胞。
- TCR 在结构上与免疫球蛋白相似,但不那么复杂。 数百万个独特的抗原表位结合 TCR 是通过 V、D 和 J 基因片段的遗传重排过程编码的。
- 根据T细胞在CD4或CD8 中的表达、与之相互作用以激活的MHC分子及其各自的功能,T细胞可以分为三类——辅助性T细胞、细胞毒性 T细胞和调节性T细胞。
- 活化的辅助 T 细胞分化为 T H 1、T H 2、T H 17 或记忆 T 细胞亚型。 分化是由它们所接触的特定细胞因子引导的。 T H 1、T H 2 和 T H 17 具有与刺激适应性和先天免疫防御相关的不同功能。 记忆 T 细胞是长寿细胞,可以对二次暴露做出快速反应。
- 一旦激活,细胞毒性 T 细胞就会靶向并杀死感染了细胞内病原体的细胞。 杀死需要使用MHC I分子识别细胞表面呈现的特定病原表位。 杀伤是由穿孔素和诱导细胞凋亡的颗粒酶介导的。
- 超抗原是细菌或病毒蛋白,可引起辅助性T细胞的非特异性激活,导致细胞因子过度释放(细胞因子风暴)和全身的、可能致命的炎症反应。
脚注
- 1 Blaschitz C.,Raffatellu M. “Th17 细胞因子和肠道粘膜屏障。” J Clin Immunol。2010 年 3 月;30 (2): 196-203。doi: 10.1007/s10875-010-9368-7。