
17.4:病原体识别和吞噬作用
- Page ID
- 200042

学习目标
- 解释白细胞如何从外周血迁移到受感染的组织
- 解释白细胞识别病原体的机制
- 解释吞噬过程以及吞噬细胞破坏和降解病原体的机制
上一节中讨论的几种细胞类型可以描述为吞噬细胞,即其主要功能是寻找、摄取和杀死病原体的细胞。 1880年代,诺贝尔奖得主动物学家伊利亚·梅奇尼科夫(Ilya Metchnikoff)(1845—1916 年)首次在海星中观察到这种被称为吞噬作用的过程,他与人类和其他动物的白细胞(WBC)建立了联系。 当时,巴斯德和其他科学家认为白细胞是在传播病原体而不是杀死病原体(结核病等某些疾病也是如此)。 但是在大多数情况下,吞噬细胞可以对各种微生物提供强大、快速和有效的防御,使其成为先天非特异性免疫的关键组成部分。 本节将重点介绍吞噬细胞寻找、识别和摧毁病原体的机制。
白细胞的外渗作用(Diapedesis)
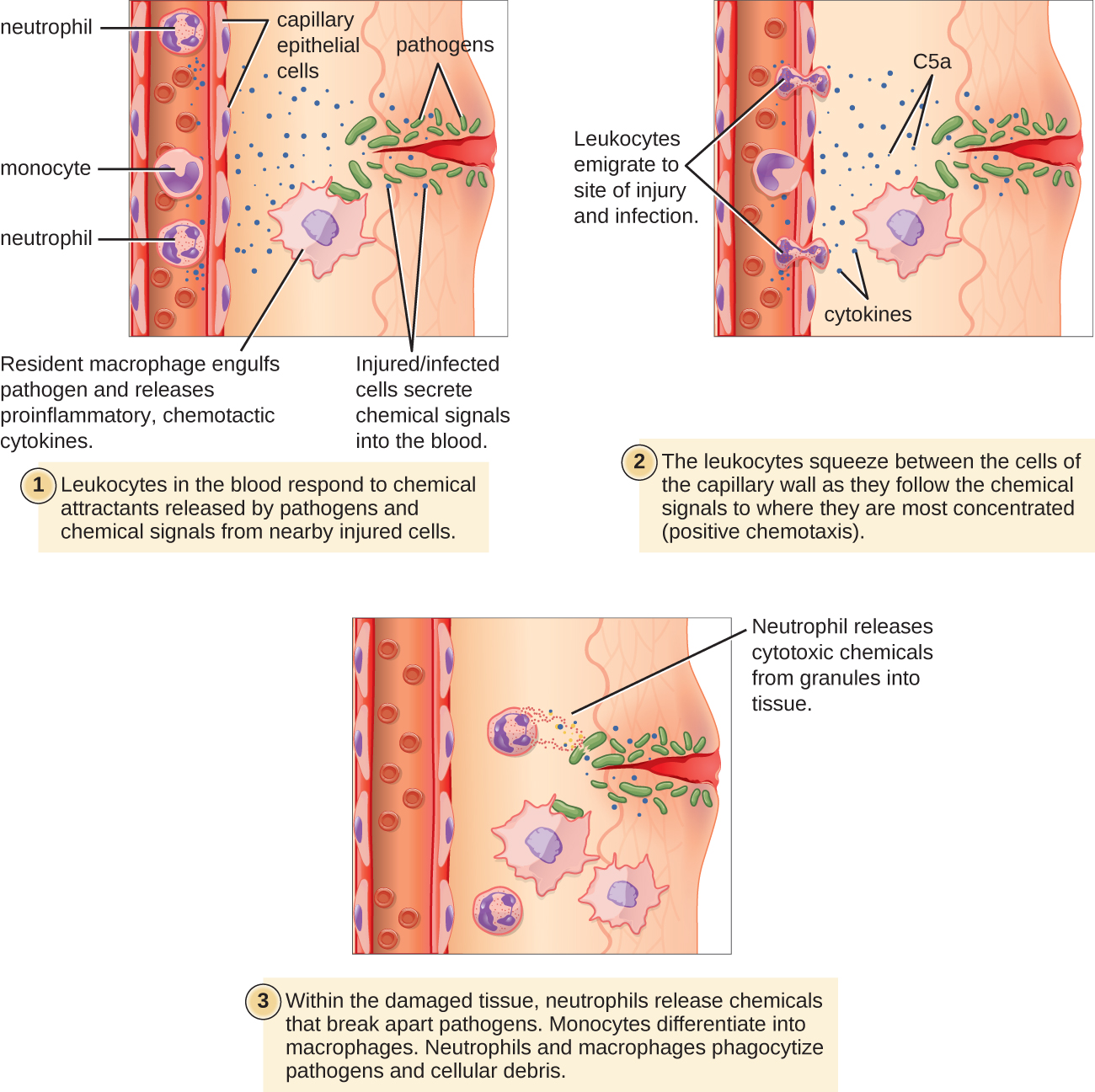
有些吞噬细胞是白细胞(WBC),通常在血液中循环。 要到达位于受感染组织中的病原体,白细胞必须穿过组织内小毛细血管的壁。 这种过程被称为 extravasation 或 diapedesis,是由补体因子 C5a 以及驻留巨噬细胞和组织细胞在传染病原体存在时释放到附近的细胞因子启动的(图\(\PageIndex{1}\))。 与 C5a 类似,这些细胞因子中有许多具有促炎和趋化作用,它们与小毛细血管细胞结合,在血管壁内侧的内皮细胞中引发反应。 这种反应涉及各种细胞粘附分子和受体的上调和表达。 经过的白细胞会稍微粘附在粘附分子上,减速并沿着感染区域附近的血管壁滚动。 当它们到达细胞交界处时,它们将与更多的粘附分子结合,在称为经内皮迁移的过程中变平并挤压细胞交界处。 这种 “滚动粘附” 机制允许白细胞离开血液进入感染区域,在那里它们可以开始吞噬入侵的病原体。
请注意,外渗不会发生在动脉或静脉中。 这些血管周围环绕着较厚的多层保护壁,而毛细血管的单细胞层薄壁形成鲜明对比。 此外,动脉中的血流过于动荡,无法进行滚动粘附。 此外,有些白细胞对感染的反应往往比其他白细胞更快。 最先到达的通常是中性粒细胞,通常在细菌感染后数小时内。 根据合同,单核细胞可能需要几天时间才能离开血液并分化为巨噬细胞。
练习\(\PageIndex{1}\)
解释粘附分子在外渗过程中的作用。
病原体识别
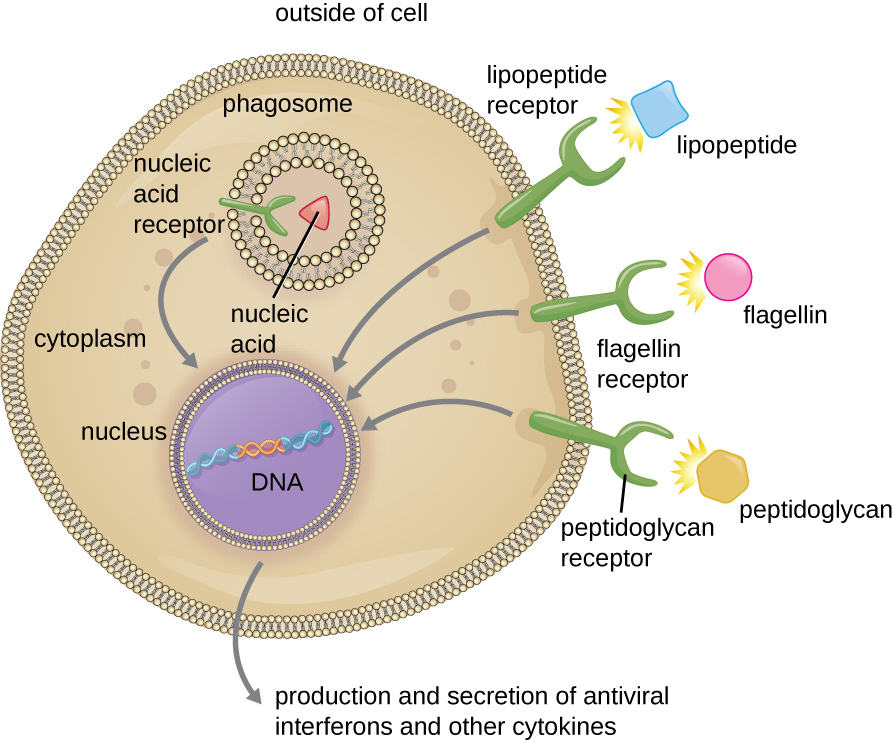
如上一节所述,通过抗体、补体因子 C1q、C3b 和 C4b 以及凝集素对病原体进行精子化可以帮助吞噬细胞识别病原体和附着,从而引发吞噬作用。 但是,并非所有的病原体识别都依赖于 opsonin。 吞噬细胞还可以识别许多致病微生物共有的分子结构。 这种结构被称为病原体相关分子模式(PAMP)。 常见的 PAMP 包括以下内容:
- 肽聚糖,存在于细菌细胞壁中;
- 鞭毛蛋白,一种存在于细菌鞭毛中的蛋白质;
- 来自革兰阴性细菌外膜的脂多糖(LPS);
- 脂肽,大多数细菌表达的分子;以及
- 核酸,例如病毒 DNA 或 RNA。
与许多其他 PAMP 一样,这些物质是各类微生物结构不可或缺的一部分。
允许吞噬细胞检测 PAMP 的结构称为模式识别受体 (PRR)。 一组 PRR 是 toll 样受体(TLR),它与各种 PAMP 结合并与吞噬细胞核通信以引起反应。 许多 TLR(和其他 PRR)位于吞噬细胞表面,但也有一些嵌入内部隔室和细胞器的膜中(图\(\PageIndex{2}\))。 这些内部 PRR 可用于结合和识别可能在吞噬作用发生之前进入细胞内部的细胞内病原体。 例如,病毒核酸可能会遇到内部 PRR,从而触发抗病毒细胞因子干扰素的产生。
除了提供病原体识别的第一步外,巨噬细胞上的 PAMP 和 PRR 之间的相互作用还提供细胞内信号,激活吞噬细胞,使其从休眠准备状态和缓慢增殖状态转变为多动状态、增殖状态细胞因子的产生/分泌,以及增强的细胞内杀伤力。 巨噬细胞上的 PRR 还会对来自受损或受压细胞的化学困扰信号做出反应。 这使巨噬细胞能够将其反应从传染病的保护范围扩展到在损伤或其他疾病引发的炎症反应中发挥更广泛的作用。
练习\(\PageIndex{2}\)
- 列举四种与病原体相关的分子模式(PAMP)。
- 描述吞噬细胞激活过程。
病原体降解
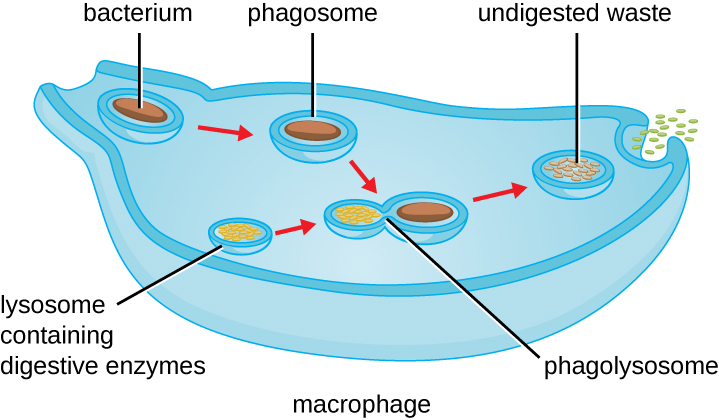
一旦发生病原体识别和附着,病原体就会被囊泡吞没,并以一种称为吞噬作用的过程带入吞噬细胞的内部隔室(图\(\PageIndex{3}\))。 PRR 可以通过首先与病原体表面结合来帮助吞噬作用,但吞噬细胞即使未与特定受体结合,也能够吞没附近的物品。 为了吞噬病原体,吞噬细胞形成伪足类,包裹病原体,然后将其捏成称为吞噬体的膜囊。 吞噬体的酸化(pH 值降至 4—5 的范围)提供了一种重要的早期抗菌机制。 含有病原体的吞噬体与一个或多个溶酶体融合,形成吞噬体。 吞噬体的形成增强了酸化,这对于激活依赖pH值的消化溶酶体酶以及产生过氧化氢和有毒活性氧至关重要。 溶菌酶、磷脂酶和蛋白酶等溶酶体酶会消化病原体。 其他酶涉及呼吸爆发。 在呼吸爆发期间,吞噬细胞会增加其对氧气的吸收和消耗,但不会用于产生能量。 氧气消耗量的增加集中在产生超氧化物阴离子、过氧化氢、羟基自由基和其他具有抗菌作用的活性氧上。
除了呼吸爆发产生的活性氧外,还可能形成具有细胞毒性(杀死细胞)潜力的活性氮化合物。 例如,一氧化氮可以与超氧化物反应形成过氧亚硝酸盐,这是一种高活性的氮化合物,其降解能力与活性氧类似。 一些吞噬细胞甚至含有杀微生物防御蛋白(例如中性粒细胞颗粒)的内部仓库。 这些破坏力可以释放到细胞周围的区域,从而在外部降解微生物。 尤其是中性粒细胞在这种次要抗微生物机制下可能非常有效。
降解完成后,剩余的废物将通过外吞囊泡从细胞中排出。 但是,重要的是要注意,并非所有病原体残留物都作为废物排出体外。 巨噬细胞和树突状细胞也是参与特异性适应性免疫反应的抗原呈递细胞。 这些细胞进一步处理降解病原体的残留物,并在其细胞表面呈现关键抗原(特定病原体蛋白)。 这是刺激某些适应性免疫反应的重要步骤,下一章将对此进行更详细的讨论。
访问此链接,查看吞噬细胞在追逐和吞没病原体的情况。
练习\(\PageIndex{3}\)
吞噬体和溶酶体有什么区别?
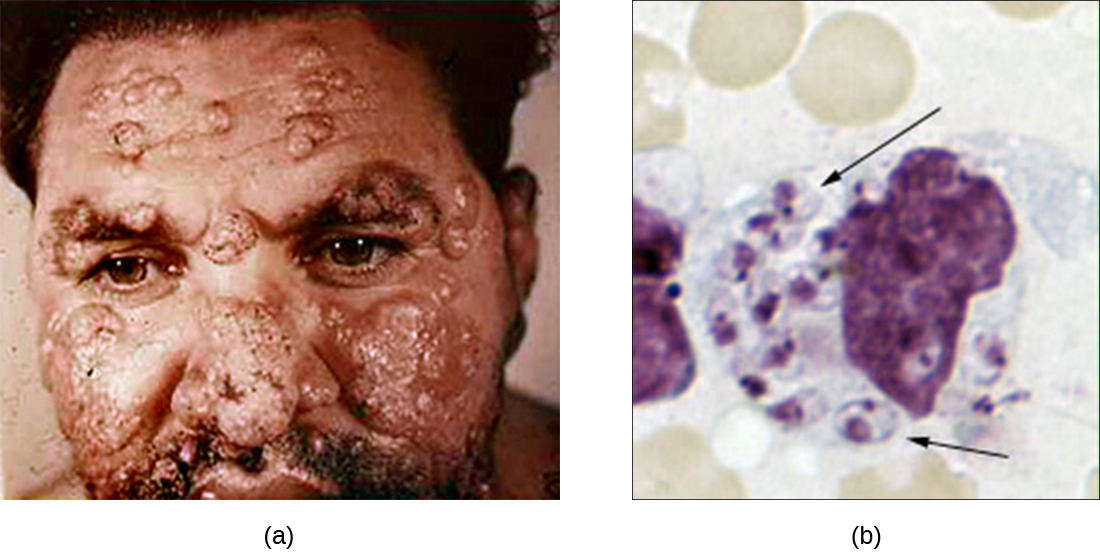
尽管吞噬作用成功摧毁了许多病原体,但有些病原体能够存活,甚至可以利用这种防御机制在体内繁殖并引起广泛的感染。 利什曼原虫属的原生动物就是一个例子。 这些专用的细胞内寄生虫是通过沙蝇叮咬传播给人类的鞭毛虫。 感染会导致皮肤和其他组织出现严重的、有时甚至是毁容性的溃疡和溃疡(图\(\PageIndex{4}\))。 据估计,全球每年有130万人新感染利什曼病。 1
来自沙蝇的唾液肽在宿主巨噬细胞叮咬部位激活宿主巨噬细胞。 补体激活的经典或替代途径是寄生虫的 C3b opsonization。 利什曼原虫细胞被吞噬,失去鞭毛,然后在吞噬体内以一种被称为 amastigote(Leishman-Donovan 体)的形式繁殖。 尽管吞噬体中有许多其他病原体被破坏,但利什曼原虫 amastigotes 的存活是通过表面脂磷聚糖和酸性磷酸酶的存在来维持的。 这些物质抑制巨噬细胞呼吸爆发和溶酶体酶。 然后,寄生虫在细胞内繁殖并裂解受感染的巨噬细胞,释放出 amastigotes 感染同一宿主内的其他巨噬细胞。 如果另一只沙蝇咬住感染者,它可能会摄入 amastigotes,然后通过另一口将其传播给另一个人。
利什曼病有几种不同的形式。 最常见的是由 L. tropica 引起的局部皮肤疾病,通常会随着时间的推移自发消退,但会有一些明显的淋巴细胞浸润和永久性疤痕。 由巴西大肠杆菌引起的一种皮肤粘膜疾病,会在鼻子和口腔组织中产生病变,并可能危及生命。 这种疾病的一种内脏形式可能是由几种不同的利什曼原虫物种引起的。 它会影响各种器官系统,导致肝脏和脾脏异常肿大。 不规则发热、贫血、肝功能障碍和体重减轻都是内脏利什曼病的体征和症状。 如果不加以治疗,通常是致命的。
关键概念和摘要
- 吞噬细胞是识别病原体并通过吞噬作用摧毁病原体的细胞。
- 识别通常通过使用吞噬细胞受体来实现,这些受体结合病原体上常见的分子,称为病原体相关分子模式(PAMP)。
- 结合 PAMP 的受体称为模式识别受体或 PRR。 T@@ oll 样受体 (TLR) 是在吞噬细胞中发现的一种PRR。
- 白细胞从血液@@ 外渗到受感染组织中是通过经内皮迁移过程发生的。
- 吞噬细胞通过吞噬作用降解病原体,吞噬作用包括吞没病原体,将其杀死并消化在吞噬体内,然后排出未消化的物质。
脚注
- 1 世界卫生组织。 “利什曼病。” 2016。 http://www.who.int/mediacentre/factsheets/fs375/en/。