
30.5: 植物中水和溶质的运输
- Page ID
- 202429

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 定义水势并解释其如何受到溶质、压力、重力和基质电位的影响
- 描述水电位、蒸散量和气孔调节如何影响植物中水的输送方式
- 解释光合物是如何在植物中运输的
植物根、茎和叶的结构促进了水分、养分和光合物在整个植物中的运输。 韧皮部和木质部是造成这种运动的主要组织。 水电位、蒸散量和气孔调节会影响水分和养分在植物中的输送方式。 要了解这些过程是如何运作的,我们必须首先了解水潜力的能量。
水的潜力
工厂是杰出的液压工程师。 仅使用基本的物理定律和对势能的简单操纵,植物就可以将水移到一棵116米高的树顶(图\(\PageIndex{1}\) a)。 工厂还可以使用液压系统产生足够的力来分裂岩石和弯曲人行道(图\(\PageIndex{1}\) b)。 植物之所以能做到这一点,是因为水的潜力。

水势是衡量水中潜在能量的指标。 植物生理学家对任何一个特定水系中的能量都不感兴趣,但对两个系统之间的水流动非常感兴趣。 因此,实际上,水势是给定水样本和纯水(在大气压力和环境温度下)之间势能的差异。 水势由希腊字母 (psi) 表示,以称为兆帕斯卡 (MPa) 的压力单位(压力是一种能量形式)表示。 顾名思义,纯水(w p ure H2O)的电位被指定为零值(尽管纯净水含有大量的势能,但该能量被忽略)。 因此,植物根、茎或叶中水的水位值是相对于纯水的 H2O 表示的。
植物溶液中的水位受溶质浓度、压力、重力和称为基质效应的因素的影响。 使用以下方程式可以将水势分解为其各个组成部分:
其中 s、p、g 和 m 分别指溶质、压力、重力和基质电位。 “系统” 可以指土壤水(土壤)、根水(根)、茎水(茎)、叶水(叶)或大气中的水(大气)的水位:无论考虑哪种水系。 随着各个组件的变化,它们会提高或降低系统的总水电位。 发生这种情况时,水会移动以保持平衡,从水位较高的系统或隔室移动到水位较低的系统或隔间。 这使两个系统之间的水位差 (Δ) 恢复为零(Δ= 0)。 因此,要使水从土壤流向空气(这种过程称为蒸腾),土壤必须是 > 根 > 茎 > 叶 > 大气层。
水只在响应 Δ时移动,而不是响应单个成分而移动。 但是,由于单个成分会影响整个系统,因此通过操纵各个组件(尤其是 s),植物可以控制水的流动。
溶质潜力
溶质电位 (s),也称为渗透电位,在植物细胞中为负,在蒸馏水中为零。 细胞质的典型值为 —0.5 至 —1.0 MPa。 溶质通过消耗水中可用的部分潜在能量来降低水电位(导致负的 w)。 溶质分子可以溶解在水中,因为水分子可以通过氢键与它们结合;像油这样的疏水分子无法与水结合,无法进入溶液。 溶质分子和水之间的氢键中的能量在系统中不再起作用,因为它被绑在键中。 换句话说,将溶质添加到水系中时,可用的势能量会减少。 因此,s 随着溶质浓度的增加而降低。 因为 s 是 system 或 t ot al 的四个分量之一,所以 s 的减少将导致总数减少。 由于细胞质的溶质含量高,植物细胞的内部水电位比纯水更为负(图\(\PageIndex{2}\))。 由于水位的这种差异,水将通过渗透过程从土壤流入植物的根细胞。 这就是为什么溶质电位有时被称为渗透电位的原因。
植物细胞可以通过添加或去除溶质分子来代谢地操纵 s(推而言之,tot al)。 因此,植物可以通过对 s 施加代谢控制的能力来控制总量。
艺术连接
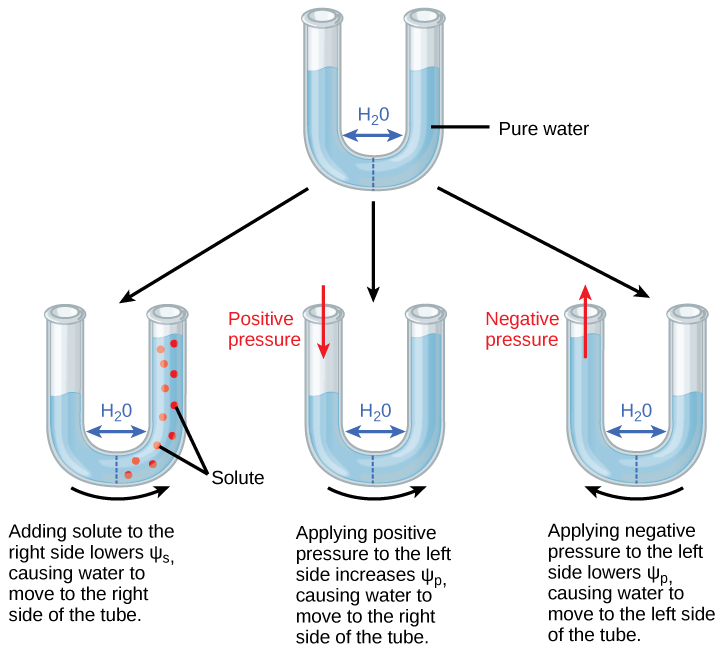
通过增加 p,使右侧的水位升高,将正水位置于管的左侧。 你能否通过添加溶质来平衡管子两侧的水位,如果是的话,怎么做?
压力潜力
压力电位 (p),也称为膨胀电位,可以是正的也可以是负的(图\(\PageIndex{3}\))。 因为压力是能量的表达,所以压力越高,系统中的势能越多,反之亦然。 因此,正的 p(压缩)会增加 tot al,负的 p(张力)会减小 tot al。 细胞内部的正压被细胞壁所控制,产生膨胀压力。 压力电位通常在0.6—0.8 MPa左右,但在浇水充足的工厂中可以高达1.5 MPa。 1.5 MPa 的 p 等于每平方英寸 210 磅(- 2 MPa 中的 1.5 MPa x 140 磅 = 210 磅/英寸 -2)。 相比之下,大多数汽车轮胎的压力保持在30—34 psi。 膨胀压力影响的一个例子是植物浇水后叶片枯萎及其恢复(图\(\PageIndex{3}\))。 水分通过蒸腾从叶片中流失(在枯萎点接近 p = 0 MPa),然后通过根部吸收恢复。
植物可以通过其操纵 s 的能力和渗透过程来操纵 p。 如果植物细胞增加细胞质溶质浓度,s 将下降,总浓度将下降,细胞与周围组织之间的 Δ将降低,水将通过渗透进入细胞,p 将增加。 p 还通过气孔的打开和关闭受到间接的植物控制。 气孔开口允许水从叶子中蒸发,减少叶子的总量,增加叶片中的水和叶柄之间的 ii,从而允许水从叶柄流入叶子。

重力电位
在没有高度的植物中,重力势 (g) 始终为负至零。 它总是从系统中移除或消耗潜在能量。 重力将水向下拉到土壤中,减少了植物水中的潜在能量总量(总计)。 植物越高,水柱越高,g 的影响力就越大。 在细胞尺度上,在短暂的植物中,这种影响可以忽略不计,很容易被忽视。 但是,在像巨型沿海红杉这样的高大树的高度上,—0.1 MPa m -1 的引力相当于额外的 1 MPa 阻力,水才能到达最高树木的树叶。 植物无法操纵 g.
矩阵电位
矩阵势 (m) 始终为负到零。 在干燥系统中,干种子中的含量可能低至—2 MPa,而在水饱和系统中则为零。 水与基质的结合总是会去除或消耗系统的潜在能量。 m 与溶质电位相似,因为它涉及通过在水和其他成分之间形成氢键来吸收水系中的能量。 但是,在溶质电位中,其他成分是可溶的亲水性溶质分子,而在 m 中,其他成分是植物细胞壁的不溶性亲水分子。 每个植物细胞都有纤维素细胞壁,细胞壁中的纤维素是亲水性的,产生水粘附的基质:因此被命名为基质电位。 m 在种子或受干旱影响的土壤等干燥组织中非常大(负值)。 但是,当种子吸收水分或土壤水合时,它很快就会变为零。 m 无法被植物操纵,在浇水充足的根、茎和叶中通常会被忽略。
水和矿物质在木质部中的流动
溶质、压力、重力和基质势对于植物中水的输送都很重要。 水从总水位较高(吉布斯自由能量较高)的区域转移到总水位较低的区域。 吉布斯自由能是与化学反应相关的能量,可用于起作用。 这表示为 Δ。
蒸腾是指通过叶片表面的蒸发从植物中流失水分。 它是木质部水流的主要驱动力。 蒸腾是由叶片-大气界面处的水分蒸发引起的;它会在叶片表面产生相当于—2 MPa的负压(张力)。 该值因蒸汽压不足而有很大差异,在较高的相对湿度 (RH) 下可以忽略不计,而在较低的相对湿度下可忽略不计。 这种张力会拉起根部的水。 到了晚上,当气孔关闭而蒸腾停止时,水会通过水粘附在木质部血管和气管的细胞壁上,以及水分子相互间的凝聚力,将水保持在茎和叶中。 这就是所谓的汁液上升的凝聚力—张力理论。
在细胞层面上,叶肉细胞表面的水会使原代细胞壁的纤维素微纤维饱和。 叶子含有许多大型细胞间空气空间,用于将氧气交换成二氧化碳,而二氧化碳是光合作用所必需的。 湿细胞壁暴露在叶片内部空气空间中,细胞表面的水蒸发到空气空间中,从而减少了叶肉细胞表面的薄膜。 这种减少会增加叶肉细胞中水的张力(图\(\PageIndex{4}\)),从而增加木质部血管中对水的拉力。 木质部血管和气管在结构上经过调整,可以应对压力的巨大变化。 容器中的环保持管状形状,就像吸尘器软管上的环在压力下保持软管处于打开状态一样。 容器元件之间的小穿孔减少了气泡的数量和大小,气泡可以通过称为空化过程形成。 木质部气泡的形成会中断从植物底部到顶部的连续水流,导致木质部汁液流中出现称为栓塞的断裂。 树越高,拉水所需的张力就越大,空化事件也越多。 在较大的树木中,由此产生的栓塞会堵塞木质部血管,使其失去功能。
艺术连接
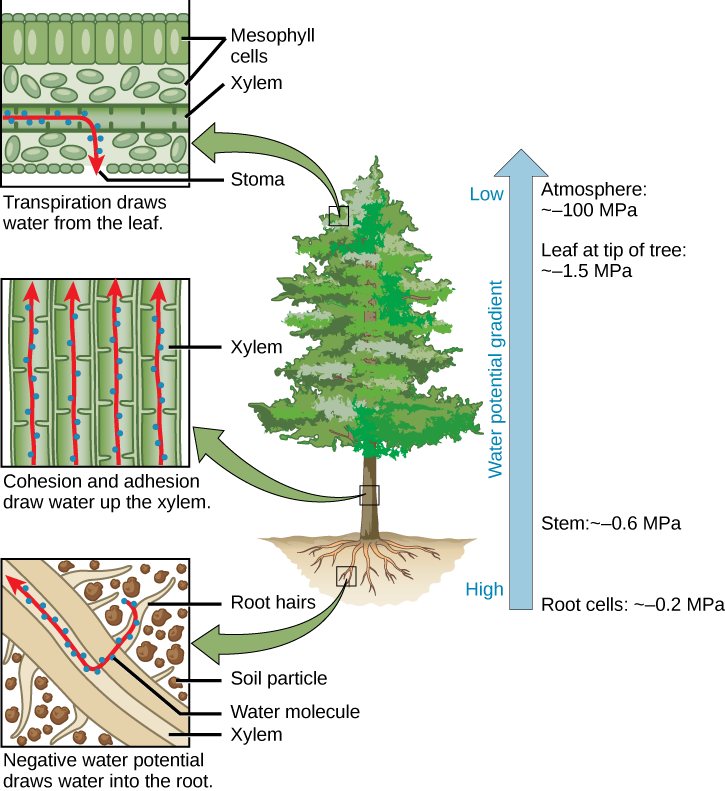
以下陈述中哪一项是错误的?
- 负水电位将水分吸入根毛。 内聚力和附着力将水分吸到木质部上。 蒸腾从树叶中吸取水分。
- 负水电位将水分吸入根毛。 凝聚力和附着力会吸收韧皮部的水分。 蒸腾从树叶中吸取水分。
- 水电位从植物的根部到顶部都有所降低。
- 水通过根毛进入植物,然后通过造口流出。
蒸腾 ——水蒸气通过口腔流失到大气中——是一个被动过程,这意味着水流不需要ATP形式的代谢能。 驱动蒸腾的能量是土壤中的水和大气中的水之间的能量差。 但是,蒸腾受到严格控制。
控制蒸腾
叶子暴露在的大气中会推动蒸腾,但也会导致植物大量水分流失。 根部吸收的水中有多达90%可能会因蒸腾而流失。
叶子被外表面的蜡状角质层覆盖,可防止水分流失。 因此,蒸腾的调节主要是通过打开和关闭叶片表面的气孔来实现的。 气孔周围有两个称为保护细胞的特殊细胞,它们根据光强度和质量、叶水状态和二氧化碳浓度等环境线索打开和关闭。 气孔必须打开,以允许含有二氧化碳和氧气的空气扩散到叶子中进行光合作用和呼吸。 但是,当气孔打开时,水蒸气会流失到外部环境中,从而提高蒸腾速度。 因此,植物必须在有效的光合作用和水分流失之间保持平衡。
随着时间的推移,植物不断进化,以适应当地环境并减少蒸腾(图\(\PageIndex{5}\))。 沙漠植物(干生植物)和生长在其他植物上的植物(附生植物)获得水的机会有限。 此类植物的蜡质角质层通常比在更温和、水分充足的环境(中生物)中生长的角质层厚得多。 水生植物(Hydrophytes)也有自己的一套解剖学和形态学叶片适应能力。

Xerophytes 和 epiphytes 通常有厚厚的毛状体或气孔覆盖物,这些毛状体或气孔沉入叶子表面以下。 毛状体是分泌油脂和物质的特殊毛发样表皮细胞。 这些适应性会阻碍气孔的空气流动,减少蒸腾。 在这些类型的植物中也经常发现多个表皮层。
在 Phloem 中运送光合物
植物需要能量来源才能生长。 在种子和球茎中,食物储存在聚合物(例如淀粉)中,这些聚合物通过代谢过程转化为蔗糖,供新发育的植物使用。 一旦绿芽和树叶生长,植物就能通过光合作用生产自己的食物。 光合作用的产物被称为光合物,通常以蔗糖等简单糖的形式出现。
为生长中的植物产生光合物的结构被称为来源。 树叶等来源产生的糖需要通过韧皮部输送到植物生长部分,这种过程称为易位。 糖的输送点,例如根、幼芽和发育中的种子,被称为水槽。 种子、块茎和球茎可以是来源,也可以是水槽,这取决于植物的发育阶段和季节。
来自源头的产物通常通过韧皮转移到最近的水槽。 例如,最高的叶子会将光合物向上发送到生长的枝尖,而较低的叶子会将光合物向下引导到根部。 中间的叶子会双向输送产物,这与木质部中的流动不同,木质部总是单向的(土壤到树叶再到大气层)。 光合物流动模式随着植物的生长和发育而变化。 Photosynthates 主要在早期引导到根部,在营养生长过程中引导到枝条和树叶,在繁殖发育过程中被引导到种子和果实。 它们还被引导到块茎中进行储存。
转移:从源头到水槽的运输
光合物,例如蔗糖,是在光合作用叶片的叶肉细胞中产生的。 从那里,它们通过韧皮部转移到使用或储存的地方。 叶肉细胞通过称为 plasmodesmata 的细胞质通道连接。 Photosynthates 通过这些通道到达血管束中的韧皮部筛管元素(SET)。 从叶肉细胞中,光合物被加载到 phloem Set 中。 利用质子梯度的电化学电位,蔗糖按照其浓度梯度(需要ATP的过程)主动输送到韧皮细胞中。 这与一种叫做 Sucrose-H + 转运蛋白的载体蛋白一起吸收蔗糖。
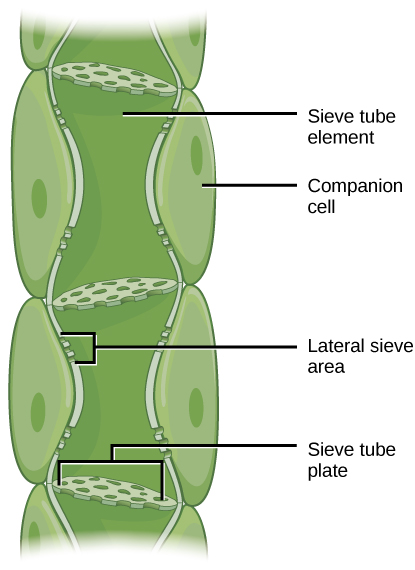
韧皮部减少了细胞质含量,并通过带有孔隙的筛板相连,允许韧皮部汁液在压力驱动下进行体积流动或易位。 伴侣细胞与 STE 有关。 它们有助于新陈代谢活动并为STE产生能量(图\(\PageIndex{6}\))。
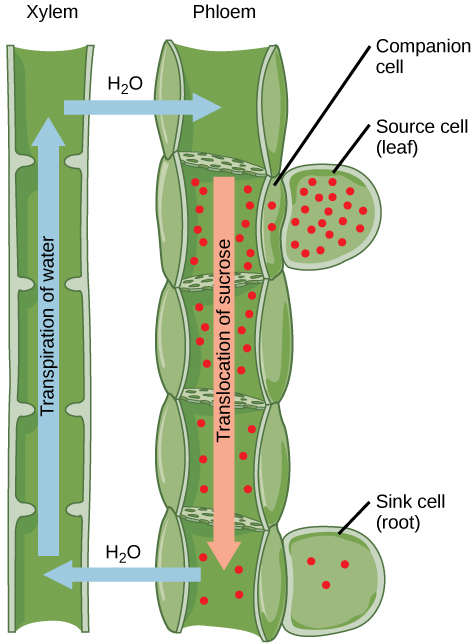
一旦进入韧皮,光合器就会被转移到最近的水槽。 韧皮液是一种水溶液,含有高达30%的糖、矿物质、氨基酸和植物生长调节剂。 高百分比的糖会降低 s,这会降低总水电位,并导致水通过渗透从相邻的木质部流入韧皮管,从而增加压力。 总水位的增加导致韧皮从源头大量流向水槽(图\(\PageIndex{7}\))。 汇细胞中的蔗糖浓度低于韧皮部中的蔗糖浓度,因为沉淀蔗糖已被代谢以促进生长,或者转化为淀粉进行储存,或者转化为其他聚合物,例如纤维素,以保持结构完整性。 韧皮管下沉端的卸载是通过蔗糖分子从高浓度区域扩散或主动输送到低浓度区域而发生的。 水通过渗透从韧皮部扩散,然后通过木质部蒸发或回收回韧皮部汁液中。
摘要
水势 () 是衡量水样本和纯水之间势能差异的指标。 植物溶液中的水势受溶质浓度、压力、重力和基质电位的影响。 水位和蒸腾会影响水在植物中通过木质部输送的方式。 这些过程受气孔打开和关闭的调节。 光合物(主要是蔗糖)通过植物的韧皮从源头转移到水槽。 蔗糖被主动加载到韧皮部的筛管元件中。 溶质浓度的增加会导致水通过渗透从木质部流入韧皮。 产生的正压将水和溶质推低压力梯度。 蔗糖被卸到水槽中,水返回到木质部容器中。
艺术联系
图\(\PageIndex{2}\):通过增加 p 使右侧水位升高,将正水位置于管的左侧。 你能否通过添加溶质来平衡管子两侧的水位,如果是的话,怎么做?
- 回答
-
是的,你可以通过在管子的左侧添加溶质来平衡水位,这样水就会向左移动直到水位相等。
图\(\PageIndex{4}\):以下陈述中哪一项是错误的?
- 负水电位将水分吸入根毛。 内聚力和附着力将水分吸到木质部上。 蒸腾从树叶中吸取水分。
- 负水电位将水分吸入根毛。 凝聚力和附着力会吸收韧皮部的水分。 蒸腾从树叶中吸取水分。
- 水电位从植物的根部到顶部都有所降低。
- 水通过根毛进入植物,然后通过造口流出。
- 回答
-
B。
词汇表
- 角质层
- 叶子和茎外侧有蜡质覆盖物,可防止水分流失
- 兆帕斯卡 (MPa)
- 测量水位的压力单位
- 下沉
- 植物的生长部分,例如根和幼叶,需要光合作用
- 资源
- 为植物产生光合物的器官
- 转移
- 维管植物中光合物从源头到水槽的质量传输
- 蒸腾
- 水蒸气通过气孔流失到大气中
- 水电位 (w)
- 在大气压力和环境温度下,单位体积水溶液相对于纯水的势能