
30.6: 植物感官系统和反应
- Page ID
- 202388

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 描述红光和蓝光如何影响植物生长和代谢活动
- 讨论 gravitropism
- 了解激素如何影响植物的生长和发育
- 描述 thigmotropism、thigmonastism 和 thigmogenesis
- 解释植物如何保护自己免受掠食者的侵害并对伤口做出反应
动物可以通过移动到新的地点来应对环境因素。 但是,植物是在原地扎根的,必须对周围的环境因素做出反应。 植物拥有先进的系统来检测和响应光线、重力、温度和物理接触。 受体感知环境因素并将信息传递给效应器系统(通常是通过中间化学信使),以产生植物反应。
植物对光的反应
植物对光有许多复杂的用途,远远超出了它们仅使用二氧化碳、光和水对低分子量糖进行光合作用的能力。 Photomorphogenesis 是植物响应光的生长和发育。 它允许植物优化对光线和空间的利用。 Photoperiodism 是利用光来追踪时间的能力。 植物可以通过感知和使用各种波长的阳光来分辨一天中的时间和一年中的时间。 Photropism 是一种定向响应,它允许植物朝向甚至远离光线生长。
环境中的光感知对植物很重要;它对竞争和生存至关重要。 植物对光的反应由不同的感光器介导,感光器由一种与称为发色团的吸光色素共价结合的蛋白质组成。 两者一起被称为染色蛋白。
可见光谱中的红色/远红色和紫蓝色区域会触发植物的结构发育。 感官感光感受器在可见光谱的这些特定区域吸收光,这是因为日光光谱中可用的光质量很高。 在陆地栖息地中,叶绿素对光的吸收在光谱的蓝色和红色区域达到顶峰。 当光线通过树冠滤光以及蓝色和红色波长被吸收时,光谱会转移到远红端,从而将植物群落转移到更适合应对远红光的植物身上。 蓝光受体使植物能够测量阳光的方向和丰度,阳光含有丰富的蓝绿色发射。 水会吸收红光,这使得检测蓝光对藻类和水生植物至关重要。
Phytochrome 系统和红/远红响应
phytochromes 是一个具有线性四吡咯发色团的色蛋白家族,类似于环状四吡咯吸光头组的叶绿素。 Phytochromes 有两种照片可互转换形式:Pr 和 Pfr。 Pr 吸收红光(约 667 nm)并立即转换为 Pfr。 Pfr 吸收远红光(约 730 nm),并快速转换回 Pr。 吸收红光或远红光会导致发色团的形状发生巨大变化,从而改变与之结合的植物色素蛋白的构象和活性。 Pfr 是蛋白质的生理活性形式;因此,暴露在红光下会产生生理活性。 暴露在远红光下会抑制植物色素活性。 这两种形式共同代表植物色素系统(图\(\PageIndex{1}\))。
植物色素系统充当生物灯开关。 它监视环境光的水平、强度、持续时间和颜色。 红光的效果是可逆的,方法是立即将远红光照射到样品上,从而将色蛋白转化为非活性的 Pr 形式。 此外,Pfr 可以在黑暗中慢慢恢复为 Pr,或者随着时间的推移而分解。 在所有情况下,红光引起的生理反应都是相反的。 活性形式的植物色素(Pfr)可以直接激活细胞质中的其他分子,也可以被运送到细胞核,在那里它直接激活或抑制特定的基因表达。
一旦植物色素系统进化,植物就会对其进行调整,以满足各种需求。 未经过滤的充足阳光所含的红光比远红光多得多。 由于叶绿素在可见光谱的红色区域吸收很强,但在远红色区域则不吸收,因此任何处于森林地面上另一种植物阴凉处的植物都将暴露在消耗红色、富含远红的光线下。 大量的远红光将阴影叶片中的植物色素转化为 Pr(非活跃)形态,从而减缓生长。 森林地面上最近的无阴影(甚至阴影较少)区域有更多的红光;暴露于这些区域的树叶会感知红光,从而激活 Pfr 形态并诱发生长。 简而言之,植物芽使用植物色素系统从阴影向光线生长。 由于在密集的植物群落中对光的竞争如此激烈,因此植物色素系统的进化优势显而易见。
在种子中,植物色素系统不用于确定光的方向和质量(阴影与无阴影)。 相反,它是否仅用于确定是否有光线。 这对于种子很小的物种(例如生菜)尤其重要。 由于生菜种子的大小,食物储备很少。 它们的幼苗在耗尽燃料之前不能长时间生长。 如果它们在土壤表面下哪怕一厘米处发芽,幼苗就永远不会在阳光下发芽,而且会死亡。 在黑暗中,植物色素处于Pr(非活性形式),种子不会发芽;只有在土壤表面的光照下它才会发芽。 暴露在光线下后,Pr 会转化为 Pfr 并开始发芽。
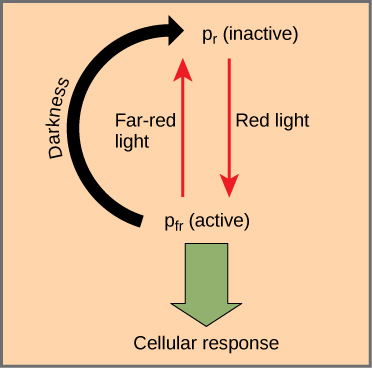
植物还使用植物色素系统来感知季节的变化。 Photoperiodism 是对白天和黑夜的时间和持续时间的生物学反应。 它控制开花、冬芽的落花和营养生长。 检测季节变化对植物的存活至关重要。 尽管温度和光照强度会影响植物的生长,但它们并不是季节的可靠指标,因为它们可能因年而异。 日长是衡量一年中时间的更好指标。
如上所述,未经过滤的阳光富含红光,但远红光不足。 因此,黎明时分,叶子中的所有植物色素分子都会迅速转化为活性的 Pfr 形态,并保持这种形态直到日落。 在黑暗中,Pfr 形态需要几个小时才能慢慢恢复到 Pr 形态。 如果夜晚很长(如冬天),则所有 Pfr 形态都会恢复。 如果夜晚很短(比如夏天),日出时可能会有相当多的 Pfr。 通过在黎明时感知Pr/Pfr比率,植物可以确定日夜周期的长度。 此外,树叶会将这些信息保留好几天,从而可以比较前一晚和前几个晚上的时长。 较短的夜晚表示植物的春天;当夜晚变长时,秋天即将来临。 这些信息,加上感知温度和水的可用性,使植物能够确定一年中的时间并相应地调整其生理状况。 短日(长夜)植物利用这些信息在夏末和初秋开花,那时夜晚超过了临界长度(通常八个小时或更短)。 长日(短夜)植物在春季开花,那时黑暗不到临界长度(通常为八到十五个小时)。 并非所有植物都以这种方式使用植物色素系统。 日间中性植物的开花不受日长调节。
职业联系:园艺家
“园艺家” 一词来自拉丁语中的花园(hortus)和文化(cultura)。 由于在理解植物对环境刺激的反应方面取得的进展,这一职业发生了革命性的变化。 农作物、水果、蔬菜和花卉的种植者以前受到限制,因为他们必须根据季节安排播种和收获时间。 现在,园艺家可以通过了解环境因素如何影响植物的生长和发育来操纵植物以增加叶子、花卉或果实的产量。
温室管理是园艺家教育的重要组成部分。 为了延长夜晚,用遮光遮阳布覆盖植物。 冬季,长日植物会受到红光照射,以促进早开花。 例如,蓝色波长较高的荧光(冷白色)可促进叶子的生长,非常适合初期幼苗。 白炽灯(标准灯泡)富含红光,可促进某些植物的开花。 通过施用植物激素,可以增加或延迟果实的成熟时间。 最近,在开发适应不同气候、抗虫害和运输损害的植物品种方面取得了长足的进展。 由于植物对外部刺激和激素的反应知识的实际应用,作物产量和质量都有所提高。
园艺家在私人和政府实验室、温室、植物园以及生产或研究领域找到工作。 他们通过运用遗传学和植物生理学知识来改善作物。 为了为园艺生涯做准备,学生要上植物学、植物生理学、植物病理学、景观设计和植物育种方面的课程。 为了补充这些传统课程,园艺专业的学生增加了经济学、商业、计算机科学和通信方面的研究。
蓝光反应
Photropism(植物朝向或远离光源的方向弯曲)是对蓝色波长光的响应。 正向光是向光源生长(图\(\PageIndex{2}\)),而负光向性(也称为 skototropism)是远离光线生长。
恰当命名的 p hotropins 是基于蛋白质的受体,负责调解光反应。 像所有植物感光器一样,photropins 由蛋白质部分和光吸收部分组成,称为发色团。 在 photropins 中,发色团是一种共价结合的黄素分子;因此,photropins 属于一类称为黄蛋白的蛋白质。
在 photropins 控制下的其他反应包括叶片的开闭、叶绿体的运动和气孔的打开。 但是,在 photropins 控制的所有反应中,photropins 的研究时间最长,也是最容易理解的。
查尔斯·达尔文和他的儿子弗朗西斯在 1880 年的论文《植物运动的力量》中首先将光向性描述为幼苗向光弯曲。 达尔文观察到光是通过植物的尖端(顶端分生体)感知到的,但反应(弯曲)发生在植物的另一部分。 他们得出的结论是,信号必须从顶端分生组织传播到工厂底部。

1913年,彼得·博伊森-詹森证明植物尖端产生的化学信号是底部弯曲的原因。 他切掉了幼苗的尖端,用一层明胶覆盖了切好的部分,然后更换了尖端。 幼苗在照亮时向灯光弯曲。 但是,当将不透水的云母片插入尖端和切割的底部之间时,幼苗没有弯曲。 实验的改进表明,信号在幼苗的阴影侧传播。 当云母板插入照明的一侧时,植物确实向光线弯曲。 因此,化学信号是一种生长刺激物,因为光致反应涉及阴影侧的细胞伸长速度比照明侧的细胞伸长速度更快。 我们现在知道,当光线穿过植物茎时,它会被衍射并在茎上产生光生素激活。 大多数激活发生在点亮的一面,导致植物激素吲哚乙酸(IAA)积聚在阴影侧。 干细胞在 IAA 的影响下会拉长。
Cryptochromes 是另一类吸收蓝光的感光器,它也含有基于黄素的发色团。 Cryptochromes 使用蓝光线索设定植物 24 小时活动周期,也称为其昼夜节律。 有证据表明,cryptochromes 与 photropins 协同作用以介导光反应。
链接到学习
使用本网站左侧面板中的导航菜单查看运动中的植物的图像。
植物对重力的反应
无论它们是在光明中还是在完全黑暗的环境中发芽,芽通常会从地面发芽,根部向下生长到地下。 如果有足够的时间,在黑暗中放置在旁边的植物会向上发出枝条。 Gravitropism 确保根部在土壤中生长,枝条朝向阳光生长。 芽顶端向上生长称为负引力,而根部向下生长称为正引力。
Amyloplast s(也称为静态体)是特殊的质体,含有淀粉颗粒,在重力作用下沉降。 淀粉样体存在于枝条和根帽的特殊细胞中。 当植物倾斜时,静态石会掉落到新的底层细胞壁上。 几个小时后,枝条或根部将显示出朝新的垂直方向生长。
调解引力主义的机制已经相当清楚了。 当淀粉样体沉淀到根部或芽中的重力感应细胞底部时,它们会物理接触内质网(ER),从而导致钙离子从急诊室内部释放。 细胞中的这种钙信号传导会导致植物激素 IAA 向细胞底部的极性转运。 在根系中,高浓度的 IAA 会抑制细胞伸长。 这种作用会减缓根部下侧的生长,而细胞在上侧发育正常。 IAA 在芽中的作用恰恰相反,芽下侧的浓度较高会刺激细胞扩张,从而导致芽长大。 芽或根部开始垂直生长后,淀粉样体恢复到正常位置。 还提出了其他假设——涉及整个细胞参与引力作用效应——来解释为什么一些缺乏淀粉样体的突变体可能仍然表现出较弱的引力反应。
增长反应
植物对外部刺激的感官反应依赖于化学信使(激素)。 植物激素影响植物生命的方方面面,从开花到结实和成熟,从光向性到落叶。 植物中的每个细胞都有可能产生植物激素。 它们可以在其起源细胞中起作用,也可以被运送到植物体的其他部分,许多植物反应涉及两种或多种激素的协同或拮抗相互作用。 相比之下,动物激素是在特定的腺体中产生的,然后运送到遥远的部位起作用,它们单独起作用。
植物激素是一组影响植物形态发生的无关化学物质。 传统上描述了五种主要的植物激素:生长素(尤其是IAA)、细胞分裂素、赤霉素、乙烯和脱落酸。 此外,其他营养素和环境条件可以说是生长因子。
Auxins
生长素一词源自希腊语 au xein,意思是 “成长”。 生长素是负责光向性和引力作用中细胞伸长的主要激素。 它们还控制分生组织向血管组织的分化,促进叶片的发育和排列。 虽然许多合成生长素被用作除草剂,但IAA是唯一具有生理活性的天然生长素。 顶端优势——抑制侧芽形成的——是由顶端分生体中产生的生长素触发的。 开花、结实和成熟以及抑制脱落(落叶)是生长素直接或间接控制下的其他植物反应。 Auxins 还充当蓝光和红/远红响应效果的继电器。
生长素的商业用途广泛用于植物苗圃和作物生产。 IAA 用作生根激素,促进插条和脱落叶片上不定根的生长。 向温室中的番茄植物施用合成生长素可促进正常的水果发育。 户外施用生长素可促进水果定型和落果的同步,以协调收获季节。 通过用生长素处理未受精的植物花,可以诱导诸如无核黄瓜之类的水果结出果实。
细胞分裂素
首次报道了细胞分裂素的作用,当时人们发现,在培养物中向发育中的植物胚胎中添加椰子的液态胚乳会刺激椰子的生长。 发现刺激性生长因子是细胞分裂素,一种促进细胞分裂(细胞分裂)的激素。 迄今为止,已知将近200种天然存在的或合成的细胞分裂素。 细胞分裂素在生长组织中最为丰富,例如发生细胞分裂的根、胚胎和果实。 众所周知,细胞分裂素可以延缓叶片组织的衰老,促进有丝分裂,并刺激芽和根部分生体的分化。 对植物发育的许多影响都受到细胞分裂素的影响,要么与生长素结合使用,要么是另一种激素。 例如,顶端优势似乎源于抑制侧芽的生长素和促进丛林生长的细胞分裂素之间的平衡。
赤霉素
赤霉素(GaS)是一组约125种密切相关的植物激素,可刺激芽长、种子发芽以及果实和花的成熟。 GaS 是在根和茎顶分生体、幼叶和种子胚胎中合成的。 在城市地区,有时会对电力线下的树木施用 GA 拮抗剂,以控制生长并减少修剪频率。
天然气会打破需要暴露在寒冷或光照下才能发芽的植物种子的休眠状态(一种生长发育受抑制的状态)。 脱落酸是 GA 作用的强效拮抗剂。 GaS 的其他影响包括性别表达、无籽果实发育以及叶子和果实的衰老延迟。 无籽葡萄是通过标准育种方法获得的,含有不起眼的种子,无法发育。 由于 GAs 是由种子产生的,而且由于果实发育和茎伸长受到 GA 的控制,因此这些葡萄品种通常会以紧凑的簇状产出小果实。 成熟的葡萄通常使用 GA 进行处理,以促进更大的果实大小,以及松散的果束(更长的茎),从而减少霉菌感染的发生率(图\(\PageIndex{3}\))。

脱落酸
植物激素脱落酸(ABA)最初被发现是导致棉铃脱落或掉落的药剂。 但是,最近的研究表明,ABA在脱落过程中的作用很小。 ABA 会积累起来,以应对紧张的环境条件,例如脱水、寒冷的温度或日程缩短。 它的活性抵消了 GaS 和 auxins 的许多促进生长的作用。 ABA 抑制茎伸长并诱导侧芽处于休眠状态。
ABA 通过阻断发芽和促进储存蛋白的合成来诱导种子休眠。 适应温带气候的植物需要很长一段时间的寒冷温度才能发芽。 这种机制可以保护幼苗在冬季异常温暖的天气中免于过早发芽。 随着荷尔蒙在冬季逐渐分解,种子会从休眠状态中释放出来,并在春季条件有利时发芽。 ABA 的另一个作用是促进冬芽的发育;它介导顶端分生组织转化为休眠芽。 土壤湿度低会导致ABA增加,从而导致气孔关闭,从而减少冬季芽的水分流失。
乙烯
乙烯与果实成熟、花枯萎和落叶有关。 乙烯不寻常,因为它是一种挥发性气体(C 2 H 4)。 几百年前,当城市街道安装煤气路灯时,生长在灯柱附近的树木会形成扭曲、变厚的树干,树叶脱落的时间比预期的要早。 这些影响是由灯泡中的乙烯挥发引起的。
老化的组织(尤其是感官叶子)和茎节会产生乙烯。 但是,这种激素最著名的作用是促进果实成熟。 乙烯刺激淀粉和酸转化为糖。 有些人将未成熟的水果(例如鳄梨)储存在密封的纸袋中以加速成熟;第一个成熟的水果释放的气体将加速剩余果实的成熟。 乙烯还会引发叶子和果实脱落、花朵褪色和掉落,促进某些谷物的发芽以及球茎和马铃薯的发芽。
乙烯广泛用于农业。 商业水果种植者通过施用气体来控制水果的成熟时间。 园艺家通过使用风扇和通风装置去除温室中的乙烯来抑制观赏植物落叶。
非传统激素
最近的研究发现了许多也会影响植物发育的化合物。 与迄今为止描述的主要激素的作用相比,人们对它们的作用知之甚少。
茉莉花在草药象牙的防御反应中起着重要作用。 当植物被捕食者伤害时,它们的水平会增加,从而导致有毒次生代谢物的增加。 它们有助于产生挥发性化合物,吸引捕食者的天敌。 例如,毛毛虫咀嚼番茄植物会导致茉莉酸含量增加,这反过来又会触发挥发性化合物的释放,吸引害虫的捕食者。
寡糖素在植物抵御细菌和真菌感染方面也起着作用。 它们在受伤部位局部起作用,也可以运送到其他组织。 Strigolactones 可促进某些物种的种子发芽,并在没有生长素的情况下抑制根尖侧向发育。 Strigolactones 还在菌根的建立中发挥了作用,菌根是植物根和真菌的互惠关联。 油菜素类固醇对许多发育和生理过程都很重要。 这些化合物与其他激素(尤其是生长素和气体)之间的信号增强了它们的生理作用。 顶端优势、种子发芽、引力作用和抗冻能力都受到激素的积极影响。 类固醇会抑制根系的生长和落果。
植物对风和触觉的反应
豌豆植物的射击围绕着格子蜿蜒而行,而一棵树则以一定角度生长,以应对盛行的强风。 这些是植物如何对触觉或风做出反应的示例。
植物在恒定方向压力下的运动被称为 thigmotropism,来自希腊语 thigma 的意思是 “触摸”,向性意味着 “方向”。 卷须就是一个例子。 卷须的分生区域对触摸非常敏感;轻触会唤起快速的卷曲反应。 与支撑表面接触的细胞会收缩,而支撑物另一侧的细胞会膨胀。 使用茉莉酸足以在没有机械刺激的情况下触发卷须卷须。
thigmonastic 反应是一种与刺激方向无关的触摸响应。 在 Venus flytrap 中,两片经过修改的叶子在铰链处连接在一起,外缘衬有细叉状的尖齿。 小毛发位于陷阱内。 当昆虫刷上这些触发器毛发,连续触摸其中的两根或更多根毛发时,树叶会迅速关闭,困住猎物。 叶片表面的腺体分泌出缓慢消化昆虫的酶。 释放的营养物质被叶子吸收,叶子会重新开放供下一顿饭食用。
Thigmomorphogenesis 是一种缓慢的发育变化,其形状呈现为承受持续机械应力的植物。 例如,当树木在风中弯曲时,生长通常会受到阻碍,树干变厚。 强化组织,尤其是木质部,是为了增加硬度以抵抗风的力量。 研究人员假设,机械应变会诱导生长和分化以增强组织。 乙烯和茉莉酸盐可能参与了 thigmomorphogenesis。

对食草动物和病原体的防御反应
植物面对两种类型的敌人:食草动物和病原体。 食草动物,无论大小,都使用植物作为食物,并积极咀嚼它们。 病原体是疾病的媒介。 这些传染性微生物,例如真菌、细菌和线虫,以植物为生并破坏其组织。 植物已经开发出各种策略来阻止或杀死攻击者。
植物中的第一道防线是完好无损且无法穿透的屏障。 树皮和蜡质角质层可以抵御掠食者的侵害。 其他针对草药象牙的改编方法包括荆棘(改良后的树枝)和刺(改良的叶子)。 它们通过造成人身伤害、诱发皮疹和过敏反应来阻止动物。 机械损伤可能会破坏植物的外部保护,这可能为病原体提供切入点。 如果第一道防线被突破,植物必须诉诸不同的防御机制,例如毒素和酶。
次生代谢物是不是直接来自光合作用的化合物,也不是呼吸或植物生长发育所必需的化合物。 许多代谢物是有毒的,甚至可能对摄入它们的动物致命。 一些代谢物是生物碱,它会阻止具有有害气味(例如薄荷和鼠尾草的挥发油)或驱虫味道(例如奎宁的苦味)的捕食者。 其他生物碱通过引起过度刺激(咖啡因就是一个例子)或与阿片类药物相关的嗜睡来影响食草动物。 一些化合物在摄入后会产生毒性;例如,木薯根中的乙二醇氰化物只有在食草动物摄入时才会释放氰化物。
机械伤害和掠食者攻击会激活受损组织中和距离伤害部位较远的部位的防御和保护机制。 一些防御反应会在几分钟内发生,而另一些则在几个小时内发生。 受感染的细胞和周围的细胞可能会死亡,从而阻止感染的传播。
长距离信号引发旨在威慑捕食者的系统性反应。 当组织受损时,茉莉酸盐可能会促进对捕食者有毒的化合物的合成。 茉莉酸盐还引发挥发性化合物的合成,这些化合物会吸引寄生虫,寄生虫是昆虫在另一种昆虫身上或身上度过发育阶段,最终杀死宿主。 如果受损组织受损无法修复,该植物可能会激活受伤组织的脱落。
摘要
植物通过形态和活性的变化对光做出反应。 红光照射将感光器植物色素转化为其远红光吸收形式——PFR。 这种形态根据一天的长短控制发芽和开花,并触发休眠植物或刚从土壤中冒出的植物的光合作用。 蓝光受体、隐色素和 photropins 是造成光向性的原因。 含有重淀粉颗粒的淀粉质体可以感知重力。 枝条表现出负重力,而根部表现出正引力。 植物激素——少量合成的天然存在的化合物——既可以在产生它们的细胞中起作用,也可以在远处的组织和器官中起作用。 Auxins 负责顶端主导地位、根系生长、向光的定向生长以及许多其他生长反应。 细胞分裂素刺激细胞分裂并对抗枝条的顶端优势。 赤霉素抑制种子的休眠状态并促进茎的生长。 脱落酸可诱导种子和芽进入休眠状态,并通过促进气孔闭合来保护植物免受过多的水分流失。 乙烯气体加速果实成熟和落叶。 植物对触摸的反应是快速运动(thigmotropy 和 thigmonasty)和缓慢的差异生长(thigmomorphogenesis)。 植物已经进化出抵御捕食者和病原体的防御机制。 树皮和刺等物理屏障可以保护嫩组织。 植物还具有化学防御能力,包括有毒的次生代谢物和激素,它们会引发额外的防御机制。
词汇表
- 脱落酸 (ABA)
- 诱导种子和其他器官休眠的植物激素
- 脱落
- 导致植物器官掉落的生理过程(例如落叶或花瓣)
- 生长素
- 影响细胞伸长(光向性)、引力、顶端优势和根系生长的植物激素
- chromophore
- 吸收光的分子
- 密码色素
- 吸收光谱蓝色和紫外线区域光的蛋白质
- 细胞分裂素
- 促进细胞分裂的植物激素
- 乙烯
- 与果实成熟、花枯萎和落叶相关的挥发性植物激素
- 赤霉素 (GA)
- 刺激芽长、种子发芽以及果实和花朵成熟和掉落的植物激素
- 茉莉花
- 源自脂肪酸亚油酸的小家族化合物
- 负引力
- 在远离地球引力的情况下生长
- 低聚糖
- 激素对植物抵御细菌和真菌感染很重要
- photormogenesis
- 植物在光照下的生长和发育
- photoperiodism
- 根据一年中的时间发生植物过程,例如发芽和开花
- photropin
- 促进光向性、气孔打开和闭合以及其他促进光合作用的反应的蓝光受体
- photroposm
- 植物向光源方向弯曲
- 植物色素
- 植物色素蛋白,以两种可逆形式(Pr 和 Pfr)存在,在红光下介导形态变化
- 阳性引力
- 向地球的引力中心成长
- statolith
- (也是 amyloplast)含有重淀粉颗粒的植物细胞器
- strigolactone
- 促进某些物种的种子发芽并在缺乏生长素的情况下抑制根尖侧发育的激素
- thigmomorphogenesis
- 对触摸的发育反应
- thigmonastic
- 植物定向生长,不受接触方向的影响
- thigmotropism
- 植物因持续接触而定向生长