
14.5: 真核生物中的 DNA 复制
- Page ID
- 202987

培养技能
- 讨论真核生物和原核生物中 DNA 复制的异同
- 陈述端粒酶在 DNA 复制中的作用
真核生物基因组比原核生物基因组复杂得多,体积也更大。 人类基因组每组单倍体染色体有30亿个碱基对,在细胞周期的S阶段复制了60亿个碱基对。 真核生物染色体上有多个复制来源;人类最多可以有 100,000 个复制来源。 复制速度约为每秒 100 个核苷酸,比原核复制慢得多。 酵母是一种真核生物,在染色体上发现了称为自主复制序列(ARS)的特殊序列。 它们等同于大肠杆菌的复制起源。
真核生物中DNA聚合酶的数量远远超过原核生物:已知有14个,其中5个已知在复制过程中起着重要作用,并已得到充分研究。 它们被称为 pol α、pol β、pol γ、po l β 和 pol α。
复制的基本步骤与原核生物中的复制步骤相同。 在开始复制之前,必须将 DNA 作为模板提供。 真核生物DNA与称为组蛋白的基本蛋白质结合,形成称为核小体的结构。 染色质(DNA和蛋白质之间的复合物)可能会经过一些化学修饰,因此DNA可能能够从蛋白质中滑落或被DNA复制机制的酶接触。 在复制的起源,复制前复合物是由其他引发剂蛋白制成的。 然后招募其他蛋白质以开始复制过程(表\(\PageIndex{1}\))。
利用 ATP 水解产生的能量的解旋酶打开 DNA 螺旋结构。 随着 DNA 的展开,复制叉会在每个复制源处形成。 双螺旋的打开会导致复制分叉之前的 DNA 过卷或超级线圈。 这些问题是通过拓扑异构酶的作用解决的。 引物由引物酶形成,使用引物,DNA pol 可以开始合成。 当前导链由 pol β 酶持续合成时,滞后链由 pol ε 合成。 一种名为PCNA(增殖细胞核抗原)的滑动钳制蛋白可将DNA pol固定在适当的位置,使其不会从DNA上滑落。 RNase H 去除 RNA 引物,然后将其替换为 DNA 核苷酸。 在用 DNA 取代 RNA 引物后,滞后链中的冈崎片段会结合在一起。 剩下的空隙由形成磷酸二酯键的 DNA 连接酶封住。
端粒复制
与原核染色体不同,真核染色体是线性的。 如你所知,DNA pol 酶只能在 5' 到 3' 方向添加核苷酸。 在前导链中,合成一直持续到染色体的末端。 在滞后链上,DNA是在短时间内合成的,每个序列都是由单独的引物引发的。 当复制分叉到达线性染色体的末端时,没有地方为在染色体末端复制 DNA 片段制作引物。 因此,这些末端保持未配对,随着时间的推移,随着细胞的继续分裂,这些末端可能会逐渐变短。
线性染色体的末端被称为端粒,其重复序列不编码特定的基因。 从某种意义上说,这些端粒可以保护基因在细胞继续分裂时不被删除。 在人类中,六碱基对序列 TTAGGG 会重复 100 到 1000 次。 端粒酶的发现(图\(\PageIndex{1}\))有助于了解染色体末端是如何维持的。 端粒酶含有催化部分和内置的 RNA 模板。 它附着在染色体的末端,并在DNA链的3'末端添加RNA模板的互补碱基。 一旦滞后链模板的3'末端足够拉长,DNA聚合酶就可以添加与染色体末端互补的核苷酸。 因此,染色体的末端被复制。
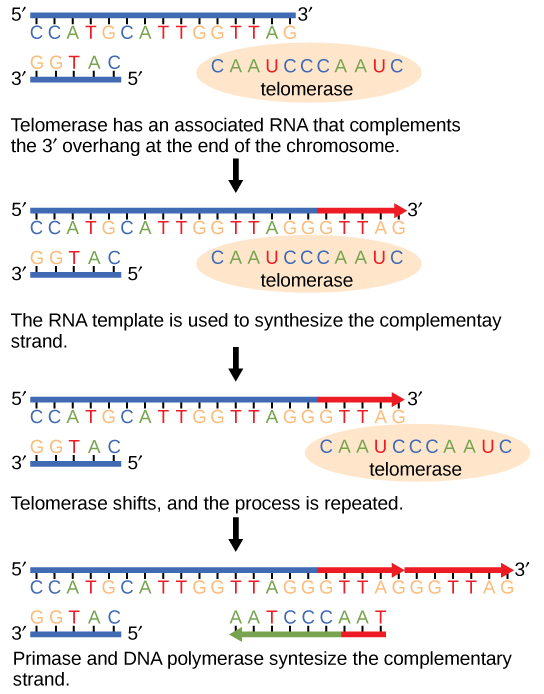
端粒酶通常活跃于生殖细胞和成体干细胞。 它在成体体细胞中不活跃。 由于发现端粒酶及其作用,伊丽莎白·布莱克本(图\(\PageIndex{2}\))于2009年获得了诺贝尔医学和生理学奖。

端粒酶和衰老
由于大多数体细胞不产生端粒酶,经历细胞分裂的细胞的端粒会继续缩短。 从本质上讲,这意味着端粒缩短与衰老有关。 随着现代医学、预防性医疗保健和更健康生活方式的出现,人类的寿命延长了,人们越来越需要随着年龄的增长看起来更年轻,生活质量也更好。
2010年,科学家发现端粒酶可以逆转小鼠体内一些与年龄相关的疾病。 这可能在再生医学中具有潜力。 1 这些研究中使用了缺乏端粒酶的小鼠;这些小鼠有组织萎缩、干细胞枯竭、器官系统衰竭和组织损伤反应受损。 这些小鼠的端粒酶重新激活导致端粒延伸,减少DNA损伤,逆转神经变性,改善睾丸、脾脏和肠道的功能。 因此,端粒再激活可能具有治疗人类年龄相关疾病的潜力。
癌症的特征是异常细胞不受控制的细胞分裂。 细胞积累突变,无法控制地增殖,并可以通过称为转移的过程迁移到身体的不同部位。 科学家们观察到,癌细胞的端粒大大缩短,端粒酶在这些细胞中具有活性。 有趣的是,只有在癌细胞中的端粒缩短之后,端粒酶才开始活跃。 如果在癌症治疗期间药物可以抑制端粒酶在这些细胞中的作用,那么癌细胞就有可能被阻止进一步分裂。
| 财产 | 原核生物 | 真核生物 |
|---|---|---|
| 复制的起源 | 单身 | 多个 |
| 复制速率 | 1000 个核苷酸/s | 50 到 100 个核苷酸/秒 |
| DNA 聚合酶类型 | 5 | 14 |
| 端粒酶 | 不在场 | 当下 |
| 去除 RNA 引物 | DNA pol I | 核糖核酸酶 H |
| 股线伸长率 | DNA pol I | pol β、pol α |
| 滑动钳 | 滑动钳 | PCNA |
摘要
真核生物中的复制始于多个复制来源。 其机制与原核生物非常相似。 启动合成需要引物,然后由DNA聚合酶扩展,因为它会将核苷酸逐一添加到生长链中。 前导链是连续合成的,而滞后的链则在短时间内合成,称为冈崎片段。 RNA 引物被 DNA 核苷酸所取代;通过将 DNA 片段与 DNA 连接酶连接起来,DNA 保持一条连续链。 染色体的末端存在问题,因为没有引物,聚合酶无法延伸染色体的末端。 端粒酶是一种内置 RNA 模板的酶,它通过复制 RNA 模板并延伸染色体的一端来延伸末端。 然后,DNA 聚合酶可以使用引物扩展 DNA。 这样,染色体的末端就受到了保护。
脚注
- 1 Jaskelioff 等人,“端粒酶再激活可逆转老年端粒酶缺乏小鼠的组织变性”,《自然》469(2011):102-7。
词汇表
- 端粒酶
- 含有催化部分和内置 RNA 模板的酶;它的作用是维持染色体末端的端粒
- 端粒
- 线性染色体末端的 DNA