
14.4: 原核生物中的 DNA 复制
- Page ID
- 202968

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 解释原核生物中 DNA 复制的过程
- 讨论不同酶和蛋白质在支持这一过程中的作用
DNA复制在原核生物中得到了非常好的研究,这主要是因为基因组体积很小,而且有可用的突变体。 大肠杆菌在单个圆形染色体中有460万个碱基对,所有碱基对在大约42分钟内被复制,从单一复制起源开始,沿着圆圈向两个方向移动。 这意味着每秒添加大约 1000 个核苷酸。 这个过程非常快速,并且没有很多错误。
DNA复制使用大量的蛋白质和酶,每种蛋白质和酶在此过程中都起着至关重要的作用。 关键参与者之一是DNA聚合酶,也称为DNA pol,它在不断增长的DNA链中逐一添加核苷酸,这些核苷酸与模板链互补。 添加核苷酸需要能量;这种能量是从附着有三个磷酸盐的核苷酸中获得的,类似于附着三个磷酸基团的 ATP。 当磷酸盐之间的键被破坏时,释放的能量用于在传入的核苷酸和生长链之间形成磷酸二酯键。 在原核生物中,已知有三种主要的聚合酶类型:DNA pol I、DNA pol II 和 DNA pol III。 现在众所周知,DNA pol III 是 DNA 合成所需的酶;DNA pol I 和 DNA pol II 主要是修复所必需的。
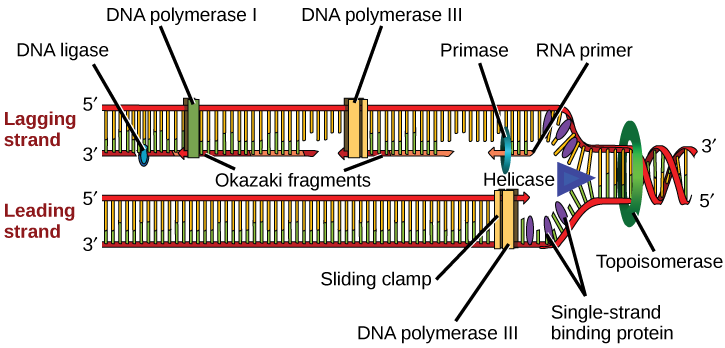
复制机制怎么知道从哪里开始? 事实证明,有特定的核苷酸序列称为复制起源,从那里开始复制。 在其一条染色体上具有单一复制源的大肠杆菌中(与大多数原核生物一样),它的长度约为245个碱基对,并且富含AT序列。 与该位点结合的某些蛋白质可以识别复制的起源。 一种叫做解旋酶的酶通过破坏含氮碱基对之间的氢键来解开 DNA。 此过程需要 ATP 水解。 随着DNA的开放,形成了称为复制叉的Y形结构。 两个复制分叉在复制的起点形成,随着复制的进行,它们会双向扩展。 单链结合蛋白覆盖复制叉附近的单链 DNA,以防止单链 DNA 回归双螺旋。 DNA聚合酶只能在5'到3'方向添加核苷酸(新的DNA链只能向这个方向延伸)。 它还需要一个游离的 3'-OH 基团,它可以通过在 3'-OH 末端和下一个核苷酸的 5' 磷酸之间形成磷酸二酯键来添加核苷酸。 这本质上意味着,如果没有游离的 3'-OH 基团,它就无法添加核苷酸。 那么它是如何添加第一个核苷酸的呢? 这个问题在提供免费的 3'-OH 端的入门的帮助下得以解决。 另一种酶 RNA primase 合成了一种 RNA 引物,该引物长约五到十个核苷酸,与 DNA 互补。 由于该序列引发了 DNA 的合成,因此它被恰当地称为引物。 DNA聚合酶现在可以扩展这种RNA引物,逐一添加与模板链互补的核苷酸(图\(\PageIndex{1}\))。
练习\(\PageIndex{1}\)
你分离出一种细胞菌株,其中冈崎片段的结合受到损害,并怀疑在复制叉上发现的酶发生了突变。 哪种酶最有可能发生突变?
- 回答
-
DNA 连接酶,因为这种酶将冈崎片段结合在一起。
复制分叉以每秒 1000 个核苷酸的速度移动。 DNA聚合酶只能向5'至3'方向延伸,这在复制分叉上构成了一个小问题。 众所周知,DNA双螺旋是反平行的;也就是说,一条链在5'到3'的方向上,另一条在3'到5'的方向上。 一条与3'至5'亲本DNA链互补的链是向复制叉连续合成的,因为聚合酶可以在这个方向上添加核苷酸。 这种连续合成的链被称为前导链。 另一条链与5'至3'的亲本DNA互补,从复制叉延伸到被称为冈崎片段的小片段中,每个片段都需要引物才能开始合成。 冈崎碎片以首次发现它们的日本科学家的名字命名。 带有冈崎碎片的链条被称为滞后链。
前导链可以单独用一根引物延伸,而滞后的股线需要为每个较短的冈崎碎片添加新的引物。 滞后股线的总方向将为3英尺至5英尺,而前导股线的总方向为5英尺至3英尺。 一种叫做滑动夹的蛋白质在DNA聚合酶继续添加核苷酸时将其固定在适当的位置。 滑动夹是一种环形蛋白质,它与 DNA 结合并将聚合酶固定在适当的位置。 Topoisomerase 可防止 DNA 双螺旋在 DNA 开放时在复制分叉之前过度缠绕;它通过在 DNA 螺旋中造成暂时的划痕然后重新封住来做到这一点。 随着合成的进行,RNA 引物被 DNA 所取代。 引物通过DNA pol I的外切核酸酶活性去除,空隙由脱氧核糖核苷酸填补。 新合成的 DNA(取代了 RNA 引物)和先前合成的 DNA 之间残留的缺口由 DNA 连接酶密封,该酶催化一个核苷酸的 3'-OH 端与另一个片段的 5' 磷酸末端之间形成磷酸二酯连接。
一旦染色体被完全复制,两个 DNA 拷贝在细胞分裂过程中会移动到两个不同的细胞中。 DNA复制过程可以概括如下。
DNA 复制步骤
- DNA 在复制的起源处解开。
- Helicase 打开了形成 DNA 的复制分叉;这些分叉是双向扩展的。
- 单链结合蛋白覆盖复制叉周围的 DNA,以防止 DNA 倒带。
- 拓扑异构酶在复制分叉之前在该区域绑定,以防止超级线圈。
- Primase 合成与 DNA 链互补的 RNA 引物。
- DNA 聚合酶开始在引物的 3'-OH 末端添加核苷酸。
- 滞后和前导股的伸长率仍在继续。
- RNA 引物通过外切核酸酶活性被去除。
- DNA pol 通过添加 dnTP 来填补空白。
- 两个DNA片段之间的间隙由DNA连接酶封住,这有助于磷酸二酯键的形成。
表\(\PageIndex{1}\)总结了参与原核生物 DNA 复制的酶以及每种酶的功能。
| 酶/蛋白质 | 特定功能 |
|---|---|
| DNA pol I | 外切核酸酶活性去除 RNA 引物,取而代之的是新合成的 DNA |
| DNA pol I | 修复功能 |
| DNA pol I | 在 5'-3' 方向添加核苷酸的主要酶 |
| Helicase | 通过破坏含氮碱基之间的氢键来打开 DNA 螺旋 |
| Ligase | 封住冈崎片段之间的空隙以创建一条连续的 DNA 链 |
| Primase | 合成开始复制所需的 RNA 引物 |
| 滑动钳 | 添加核苷酸时有助于将 DNA 聚合酶保持在原位 |
| 拓扑异构酶 | 通过造成 DNA 断裂然后重新密封,帮助缓解 DNA 在解卷时承受的压力 |
| 单链结合蛋白 (SSB) | 与单链 DNA 结合,避免 DNA 倒带。 |
链接到学习
在此处查看 DNA 复制的完整过程。
摘要
原核生物中的复制从染色体上发现的称为复制起源的序列开始,这是 DNA 打开的点。 解旋酶打开了 DNA 双螺旋结构,从而形成了复制分叉。 单链结合蛋白与复制叉附近的单链 DNA 结合,以保持分叉开放。 Primase 合成 RNA 引物以启动 DNA 聚合酶的合成,DNA 聚合酶只能在 5' 到 3' 方向添加核苷酸。 一条链沿着复制分叉的方向连续合成;这被称为前导链。 另一条链是在远离复制分叉的方向上合成的,用称为冈崎片段的短片段DNA片段合成。 这条线被称为滞后链。 复制完成后,RNA引物被DNA核苷酸所取代,DNA用DNA连接酶密封,DNA连接酶在一端的3'-OH和另一端的5'磷酸盐之间产生磷酸二酯键。
词汇表
- 解旋酶
- 在复制过程中,这种酶通过破坏氢键来帮助打开 DNA 螺旋结构
- 滞后股线
- 在复制过程中,在短片段中复制的链,远离复制分叉
- 领先的股线
- 在 5'-3' 方向连续合成的链,沿复制分叉的方向合成
- 连接酶
- 催化在 DNA 的 3' OH 和 5' 磷酸末端之间形成磷酸二酯连接的酶
- 冈崎片段
- 在滞后链上短时间合成的 DNA 片段
- primase
- 合成 RNA 引物的酶;DNA pol 需要该引物才能开始合成新 DNA 链
- 底漆
- 启动复制所需的核苷酸很短;如果是复制,则引物含有 RNA 核苷酸
- 复制分叉
- 在复制开始时形成的 Y 形结构
- 单链结合蛋白
- 在复制过程中,与单链 DNA 结合的蛋白质;这有助于将两条 DNA 链分开,以便它们可以用作模板
- 滑动钳
- 环形蛋白,将 DNA pol 保持在 DNA 链上
- 拓扑异构酶
- 在 DNA 复制时导致 DNA 下卷或过卷的酶