
13.2: 遗传性疾病的染色体基础
- Page ID
- 202842

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 描述核图是如何创建的
- 解释非分离是如何导致染色体数量失调的
- 比较非整倍体引起的疾病
- 描述染色体结构错误是如何通过反转和易位发生的
当染色体在减数分裂期间表现异常时,就会出现遗传性疾病。 染色体疾病可分为两类:染色体数量异常和染色体结构重排。 因为即使是很小的染色体片段也可以跨越许多基因,所以染色体疾病通常是严重的,而且往往是致命的。
染色体的鉴定
染色体的分离和微观观察构成了细胞遗传学的基础,也是临床医生检测人类染色体异常的主要方法。 核型是染色体的数量和外观,包括染色体的长度、带状图案和着丝粒位置。 为了获得个体核型的视图,细胞学家对染色体进行拍照,然后将每条染色体剪切并粘贴到图表或核图中,也称为表意文字(图\(\PageIndex{1}\))。
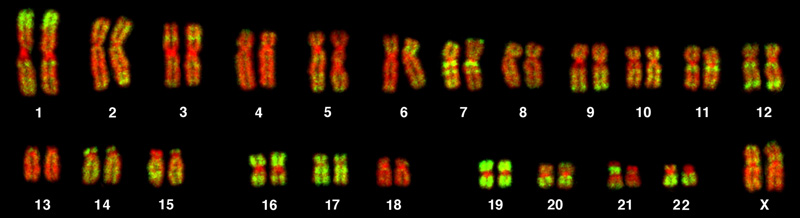
在给定物种中,染色体可以通过其数量、大小、着丝粒位置和带状模式进行识别。 在人类核型中,常染色体或 “体染色体”(所有非性染色体)通常按从最大(1号染色体)到最小(22号染色体)的大致大小顺序排列。 X 和 Y 染色体不是常染色体。 但是,21号染色体实际上比22号染色体短。 这是在将唐氏综合症命名为21号三体症之后发现的,这反映了这种疾病是由于拥有一个额外的21号染色体(总共三条)而产生的。 尽管21号染色体描述了最短的染色体集,但不想更改这种重要疾病的名称,但仍保留了其编号。 从着丝粒两端伸出的染色体 “臂” 可以被指定为短或长,具体取决于它们的相对长度。 短臂缩写为 p(代表 “petite”),而长臂缩写为 q(因为它按字母顺序跟在 “p” 之后)。 每只手臂被进一步细分并用数字表示。 使用这种命名系统,可以在科学文献中一致地描述染色体上的位置。
职业联系:遗传学家使用核图识别染色体畸变
尽管孟德尔被称为 “现代遗传学之父”,但他在进行实验时没有使用当今遗传学家经常使用的工具。 其中一种如此强大的细胞学技术是核型分型,这种方法可以从单个细胞中识别出以染色体异常为特征的特征。 为了观察个体的核型,首先从血液样本或其他组织中采集一个人的细胞(如白细胞)。 在实验室中,分离的细胞受到刺激,开始主动分裂。 然后将一种叫做秋水仙碱的化学物质应用于细胞,以在中期阶段阻断浓缩的染色体。 然后使用低张溶液使细胞肿胀,这样染色体就会分开。 最后,将样品保存在固定剂中并涂在载玻片上。
然后,遗传学家用几种染料中的一种对染色体进行染色,以更好地可视化每对染色体不同且可重复的带状图案。 染色后,使用明场显微镜观察染色体。 常见的污渍选择是 Giemsa 染色剂。 Giemsa 染色在所有 23 对染色体上排列了大约 400—800 条带(紧密盘绕的 DNA 和浓缩蛋白);经验丰富的遗传学家可以识别每个带。 除了带状图案外,还可以根据大小和着丝粒位置进一步识别染色体。 为了获得核型的经典描述,其中同源染色体按从长到短的数字顺序排列,遗传学家获得了一张数字图像,识别了每条染色体,然后手动将染色体排列成这种模式(图\(\PageIndex{1}\))。
最基本的是,核型图可以揭示遗传异常,即个体每个细胞的染色体过多或太少。 这方面的例子有唐氏综合症(由21号染色体的第三个拷贝鉴定)和特纳综合症(其特征是女性中只有一条X染色体,而不是正常的两条)。 遗传学家还可以识别出大量缺失或插入的DNA。 例如,雅各布森综合症(涉及独特的面部特征以及心脏和出血缺陷)是通过11号染色体的缺失来识别的。 最后,核型可以精确定位易位,当一段遗传物质从一条染色体中断并重新连接到另一条染色体或同一染色体的另一部分时,就会发生易位。 易位与某些癌症有关,包括慢性髓系白血病。
在孟德尔的一生中,继承是一个抽象的概念,只有通过交叉和观察后代所表达的特征才能推断出来。 通过观察核组图,当今的遗传学家实际上可以直观地看到个体的染色体组成,以确认或预测后代的遗传异常,甚至在出生之前。
染色体数异常
在所有染色体疾病中,染色体数量的异常是从核型图中最明显识别出来的。 染色体数量失调包括整条染色体的重复或丢失,以及整组染色体数量的变化。 它们是由非分离引起的,这种情况发生在减数分裂期间成对的同源染色体或姊妹染色体无法分离时。 突触错位或不完整,或促进染色体迁移的主轴装置功能失调,都可能导致非分离。 发生非分离的风险随着父母年龄的增长而增加。
在减数分裂 I 或 II 期间都可能出现非分离,结果不同(图\(\PageIndex{2}\))。 如果在减数分裂 I 期间同源染色体无法分离,则结果是两个配子缺少该特定染色体,两个配子有两个染色体拷贝。 如果在减数分裂 II 期间姐妹染色体无法分离,则结果是一个配子缺少该染色体,两个普通配子有一份染色体,一个配子有两个染色体拷贝。
艺术连接
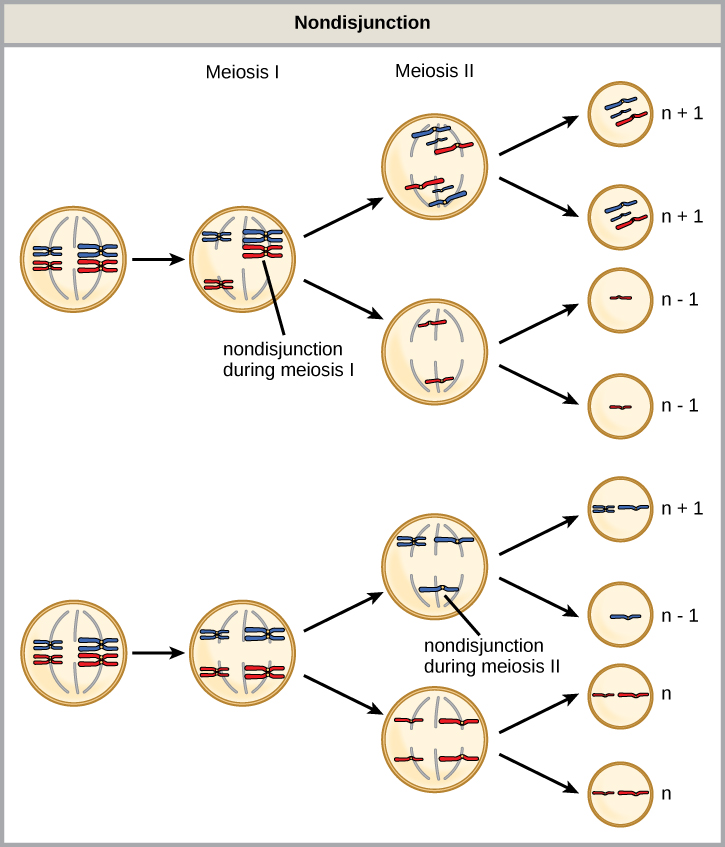
以下关于非分解的陈述中哪一项是正确的?
- 非分离只能生成具有 n+1 或 n—1 染色体的配子。
- 减数分裂 II 期间出现的非分离会导致 50% 的正常配子。
- 减数分裂 I 期间的非分离会导致 50% 的正常配子。
- 非分离总是会产生四种不同的配子。
非整倍体
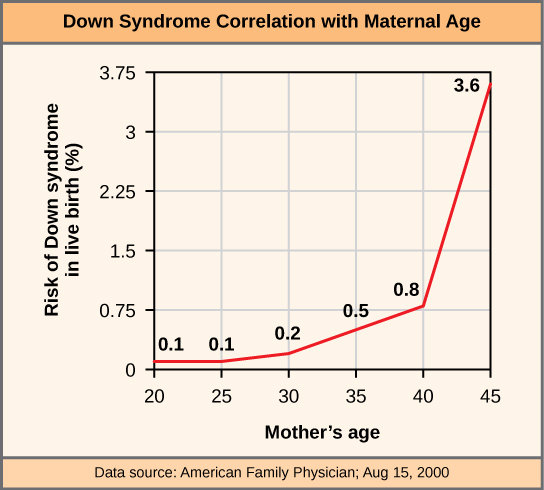
拥有适合其物种染色体数量的个体称为整倍体;在人类中,整倍体相当于22对常染色体和一对性染色体。 染色体数目错误的个体被描述为非整倍体,该术语包括单体性(失去一条染色体)或三体性(外来染色体的增加)。 缺少任何一个常染色体拷贝的单体人类合子总是无法发育到出生,因为它们缺乏必需的基因。 这突显了 “基因剂量” 在人体中的重要性。 大多数常染色体三体也无法在出生时发育;但是,一些较小的染色体(13、15、18、21 或 22)的重复可能导致后代存活数周至数年。 Trisomic 个体患有另一种类型的遗传失衡:基因剂量过多。 拥有额外染色体的个体可以合成该染色体编码的大量基因产物。 这种额外剂量(150%)的特定基因可能导致许多功能挑战,并且往往会阻碍发育。 存活新生儿中最常见的三体性是21号染色体,它对应于唐氏综合症。 患有这种遗传性疾病的人的特征是身材矮小、手指发育迟缓、面部差异(包括颅骨宽大、舌头大)以及严重的发育迟缓。 唐氏综合症的发病率与孕产妇年龄相关;老年女性更有可能怀上携带21型三体基因型的胎儿(图\(\PageIndex{3}\))。
链接到学习
在此视频模拟中可视化添加导致唐氏综合症的染色体。
多倍体
染色体集数超过正确数量(二倍体物种为两组)的个体称为多倍体。 例如,将异常的二倍体卵与正常的单倍体精子受精会产生三倍体合子。 多倍体动物极为罕见,扁虫、甲壳类动物、两栖动物、鱼类和蜥蜴中只有少数例子。 多倍体动物是不育的,因为减数分裂无法正常进行,而是主要产生无法产生活合子的非整倍体子细胞。 在极少数情况下,多倍体动物可以通过单倍体无性繁殖,在这种情况下,未受精的卵通过有丝分裂分裂产生后代。 相比之下,多倍体在植物界非常常见,多倍体植物往往比其物种的整倍体更大、更强壮(图\(\PageIndex{4}\))。

人类的性染色体非分离
人类对常染色体三体和单体表现出剧烈的有害影响。 因此,尽管携带不同数量的X染色体,但人类雌性和雄性仍能正常运作,这似乎违反直觉。 性染色体数量的变化与常染色体的增加或减少无关,而是与相对较轻的影响有关。 这在一定程度上是由于一种叫做 X 失活的分子过程造成的。 在发育初期,当雌性哺乳动物胚胎仅由几千个细胞(相对于新生儿中的数万亿个细胞)组成时,每个细胞中的一条 X 染色体会紧密凝结成一种称为巴尔体的静态(休眠)结构而失活。 X染色体(母系或父系衍生)在每个细胞中失活的几率是随机的,但是一旦失活,来自该染色体的所有细胞将具有相同的非活性X染色体或Barr体。 通过这个过程,雌性补偿了X染色体的双重遗传剂量。 在所谓的 “龟甲” 猫中,胚胎 X 失活被观察为颜色杂色(图\(\PageIndex{5}\))。 对于 X 连锁外套颜色基因具有杂合性的雌性将在其身体的不同部位表现出两种不同的外套颜色中的一种,对应于该区域的胚胎细胞祖先中灭活的 X 染色体。

携带异常数量的 X 染色体的个体将使每个细胞中除一条 X 染色体之外的所有染色体失活。 但是,即使是灭活的X染色体也能继续表达一些基因,X染色体必须重新激活才能使雌性卵巢正常成熟。 因此,X 染色体异常通常与轻微的精神和身体缺陷以及不育有关。 如果X染色体完全不存在,则该个体不会在子宫内发育。
已经确定了性染色体数目中的几个错误。 拥有三条 X 染色体(称为 Triplo-X)的个体表型为女性,但表现为发育迟缓和生育能力降低。 XXY基因型对应于一种类型的Klinefelter综合征,对应于表型为睾丸小、乳房肿大、体毛减少的男性个体。 存在更复杂的Klinefelter综合征类型,其中个体有多达五条X染色体。 在所有类型中,除一条染色体外,每条 X 染色体都会失活,以补偿多余的遗传剂量。 这可以看作是每个细胞核中有几个 Barr 体。 特纳综合征的特征是X0基因型(即只有一条性染色体),对应于一个表型的女性个体,身材矮小,颈部有网状皮肤,听力和心脏损伤以及不育。
重复和删除
除了整条染色体的丢失或增加外,染色体片段还可能重复或丢失。 重复和缺失通常会产生存活但表现出身心异常的后代。 重复的染色体片段可能与现有染色体融合,也可能在细胞核中游离。 Cri-du-chat(来自法语,意为 “猫的哭泣”)是一种与神经系统异常和可识别的身体特征相关的综合征,这些异常是由于缺失了大部分5p(5号染色体的小臂)而导致的(图\(\PageIndex{6}\))。 患有这种基因型的婴儿会发出典型的尖叫声,这是该疾病的名字的基础。
染色体结构重排
细胞学家描述了染色体中的许多结构重排,但最常见的是染色体反转和易位。 两者都是在减数分裂期间通过将重新排列的染色体与其以前的同系物进行适应性配对来鉴定的,以保持适当的基因对齐。 如果两个同系物上携带的基因方向不正确,则重组事件可能导致一条染色体上的基因丢失,而另一条染色体上的基因增加。 这将产生非整倍体配子。
染色体反转
染色体反转是指部分染色体的脱离、180° 旋转和重新插入。 在自然界中,反转可能是由于机械剪切或转座元素(借助剪切和粘贴 DNA 序列的酶,能够促进染色体片段重排的特殊 DNA 序列)的作用而发生的。 除非它们破坏基因序列,否则反转只会改变基因的方向,并且可能比非整倍体错误产生更温和的影响。 但是,基因取向的改变可能导致功能变化,因为基因表达的调节剂相对于其靶标可能会偏离位置,从而导致基因产物水平异常。
反转可以是周中心的,包括着丝粒,也可以是副中心的,发生在着丝粒之外 ()。 围绕着丝粒不对称的周中心反转可以改变染色体臂的相对长度,从而使这些反转很容易识别。
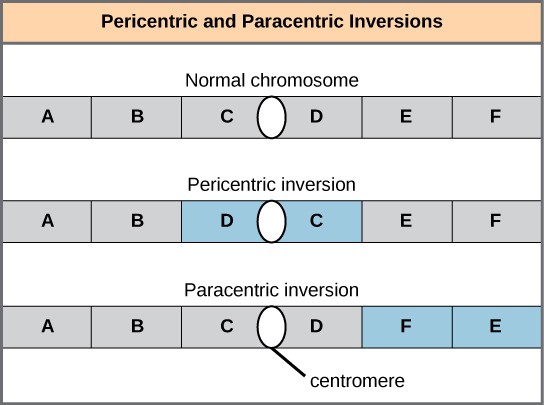
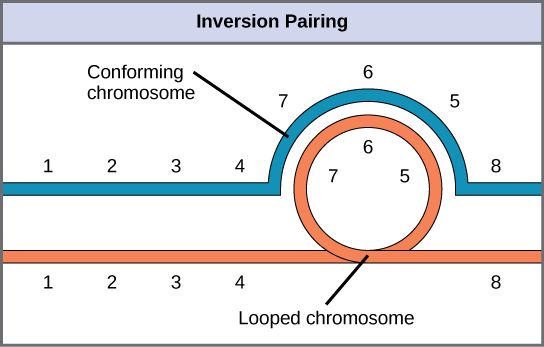
当一条同源染色体经历反转而另一条未发生反转时,该个体被描述为反转杂合子。 为了在减数分裂期间保持点对点突触,一个同系物必须形成一个循环,另一个同系物必须围绕它塑造。 尽管这种拓扑结构可以确保基因正确对齐,但它也会迫使同系物伸展,并可能与突触不精确的区域相关联(图\(\PageIndex{8}\))。
进化连接:18 号染色体反转
并非所有染色体的结构重排都会产生无法存活、受损或不育的个体。 在极少数情况下,这种变化可能导致新物种的进化。 实际上,18号染色体的周中心反转似乎促成了人类的进化。 这种反转不存在于我们最亲近的遗传亲属黑猩猩身上。 人类和黑猩猩的细胞遗传学差异在于多条染色体的周中心反转以及黑猩猩中与人类二号染色体相对应的两条单独染色体的融合。
据信,18号染色体的反转发生在早期人类中,大约在五百万年前他们与黑猩猩的共同祖先分歧之后。 描述这种反转特征的研究人员表明,在18p上复制了大约19,000个核苷酸碱基,复制的区域反转并重新插入了祖先人类的18号染色体上。
对这种反转区域的人类和黑猩猩基因的比较表明,黑猩猩 17 号染色体(对应于人类 18 号染色体)上相邻的两个基因(ROCK1 和 USP14)在人类 18 号染色体上的位置更远。 这表明其中一个反转断点发生在这两个基因之间。 有趣的是,人类和黑猩猩在包括皮质细胞和成纤维细胞在内的特定细胞类型中以不同的水平表达 USP14。 也许祖先人类的18号染色体反转重新定位了特定的基因,并以一种有用的方式重置了它们的表达水平。 由于 ROCK1 和 USP14 都编码细胞酶,因此其表达的变化可能会改变细胞功能。 目前尚不清楚这种反转是如何促成人类进化的,但它似乎是人类与其他灵长类动物分歧的重要因素。 1
易位
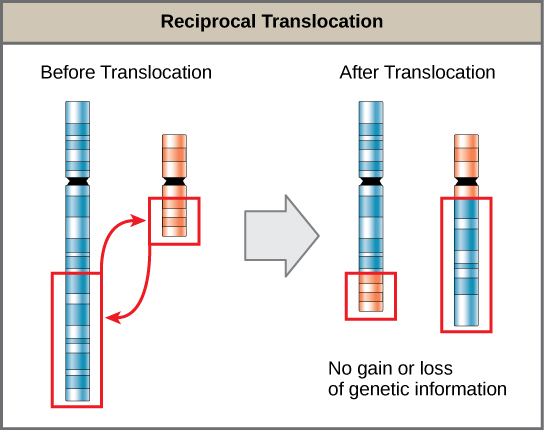
当一段染色体分离并重新连接到另一条非同源染色体时,就会发生易位。 易位可能是良性的,也可以具有毁灭性影响,这取决于基因相对于调控序列的位置是如何改变的。 值得注意的是,特定的易位与几种癌症和精神分裂症有关。 相互易位是由于两条非同源染色体之间的染色体片段交换造成的,因此遗传信息不会获得或丢失(图\(\PageIndex{9}\))。
摘要
染色体的数量、大小、形状和带状模式使染色体在核图中易于识别,并允许评估许多染色体异常。 尽管一些三组基因型是可行的,但染色体数量失调或非整倍体通常对胚胎是致命的。 由于 X 失活,性染色体中的畸变通常会产生较轻的表型效应。 非整倍体还包括染色体片段被复制或删除的情况。 染色体结构也可以重新排列,例如通过反转或易位。 这两种畸变都可能导致有问题的表型效应。 由于它们迫使染色体在减数分裂期间呈现不自然的拓扑结构,因此反转和易位通常与生育能力下降有关,因为可能出现非分离。
艺术联系
图\(\PageIndex{2}\):以下关于非分解的陈述中哪一项是正确的?
- 非分离只能生成具有 n+1 或 n—1 染色体的配子。
- 减数分裂 II 期间出现的非分离会导致 50% 的正常配子。
- 减数分裂 I 期间的非分离会导致 50% 的正常配子。
- 非分离总是会产生四种不同的配子。
- 回答
-
B。
脚注
- 1 Violaine Goidts 等人,“与 18 号染色体的人类特异性反转相关的节段重复:分段重复对灵长类动物核型和基因组进化的影响的又一个例子”,H uman Gen etics。115 (2004): 116-122
词汇表
- 非整倍体
- 染色体数目有误的个体;包括染色体片段的缺失和重复
- 常染色体
- 任何非性染色体
- 染色体反转
- 脱离、180° 旋转、染色体手臂重新插入
- 整倍体
- 拥有与其物种相应数量的染色体的个体
- 核图
- 核型的照片图像
- 核型
- 个体染色体的数量和外观;包括大小、带状图案和着丝粒位置
- 单体性
- 否则是缺少一条染色体的二倍体基因型
- 非分离
- 在减数分裂的第一次细胞分裂期间,突触同系物未能完全分离并迁移到单独的极点
- paracentric
- 在着丝粒之外发生的反转
- pericentric
- 涉及着丝粒的反转
- 多倍体
- 染色体组数不正确的个体
- 转移
- 染色体的一个片段分离并重新连接到另一条非同源染色体的过程
- 三体性
- 否则是二倍体基因型,其中整条染色体是重复的
- X 失活
- 在雌性胚胎发育过程中,X 染色体凝结到 Barr 体内,以补偿双倍的遗传剂量