
13.1: 染色体理论和遗传联系
- Page ID
- 202826

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 讨论萨顿的染色体遗传理论
- 描述遗传联系
- 解释同源重组或交叉的过程
- 描述染色体图谱是如何创建的
- 使用三点测试十字架计算染色体上三个基因之间的距离
早在显微镜下可视化染色体之前,现代遗传学之父格雷戈尔·孟德尔就在1843年开始研究遗传。 随着1800年代后期微观技术的改进,细胞生物学家可以用染料对亚细胞结构进行染色和可视化,并观察它们在细胞分裂和减数分裂过程中的作用。 每次有丝分裂分裂时,染色体都会复制,从无定形(不恒定形状)的核块凝结成不同的 X 形体(成对相同的姐妹染色体),然后迁移到单独的细胞极。
染色体遗传理论
有人猜测染色体可能是理解遗传的关键,这促使几位科学家研究了孟德尔的出版物,并根据有丝分裂和减数分裂期间染色体的行为重新评估了他的模型。 1902年,西奥多·博维里观察到,除非存在染色体,否则海胆的胚胎发育不会正常。 同年,沃尔特·萨顿观察到减数分裂期间染色体分离为子细胞(图\(\PageIndex{1}\))。 这些观察共同促成了染色体遗传理论的发展,该理论将染色体确定为造成孟德尔遗传的遗传物质。

染色体遗传理论与孟德尔定律一致,并得到以下观察结果的支持:
- 在减数分裂期间,同源染色体对作为独立于其他染色体对的离散结构迁移。
- 将每对同源染色体分为配子前似乎是随机的。
- 每个亲本合成的配子仅含有其染色体补体的一半。
- 尽管雄配子和雌配子(精子和卵子)的大小和形态不同,但它们的染色体数量相同,这表明父母双方的遗传贡献相等。
- 配子染色体在受精过程中结合在一起,产生与父母染色体数量相同的后代。
尽管减数分裂期间染色体的行为与孟德尔的抽象定律之间存在令人信服的相关性,但染色体遗传理论早在没有直接证据表明染色体上存在特征之前就已经提出。 批评者指出,与染色体相比,个人分离特征的独立性要高得多。 托马斯·亨特·摩根(Thomas Hunt Morgan)在与果蝇果蝇 melanogaster 进行了几年交叉之后,才提供了支持染色体遗传理论的实验证据。
遗传连锁和距离
孟德尔的研究表明,特质是相互独立继承的。 摩根发现分离特征与X染色体之间有 1:1 的对应关系,这表明染色体的随机分离是孟德尔模型的物理基础。 这也证明相关基因会破坏孟德尔的预测结果。 每条染色体可以携带许多关联基因这一事实解释了个体如何具有比染色体更多的特征。 但是,摩根实验室研究人员的观察表明,位于同一染色体上的等位基因并不总是一起遗传。 在减数分裂期间,关联基因以某种方式失去了联系。
同源重组
1909 年,弗兰斯·詹森在减数分裂第一次分裂之前观察到了 chiasmata,即染色体相互接触并可能交换片段的点。 他建议等位基因失去联系,染色体在物理上交换片段。 当染色体凝结并与同系物配对时,它们似乎在不同的点相互作用。 詹森认为,这些点对应于交换染色体片段的区域。 现在众所周知,同源染色体(称为突触)之间的配对和相互作用不仅仅是组织同系物以便迁移到单独的子细胞。 当突触时,同源染色体在手臂上进行相互的物理交换,这种过程称为同源重组,或者更简单地说,“交叉”。
为了更好地了解研究人员目前获得的实验结果类型,可以考虑一个杂合个体,该个体继承了同一染色体上两个基因(例如 AB)的显性母体等位基因,以及相同基因(例如 AB)的两个隐性父系等位基因ab)。 如果这些基因相互关联,人们会期望这个人产生 AB 或 ab 的配子,比例为 1:1。 根据孟德尔独立分类的概念,如果基因没有关联,则个体应产生相同频率的 AB、Ab、aB和ab 配子。 由于它们对应于新的等位基因组合,Ab 和 aB 是非亲本类型,由减数分裂期间的同源重组产生。 父母类型是与父母表现出相同等位基因组合的后代。 但是,摩根和他的同事发现,当这样的杂合个体被测试交叉到纯合隐性父母(aaBb × aabb)时,父母和非父母病例都会发生。 例如,可以找回 950 个 aabb 或 aabb 的后代,但还会获得 50 个后代,分别是 Aabb 或 aabb。 这些结果表明,这种联系最常发生,但有相当一部分的后代是重组的产物。
艺术连接
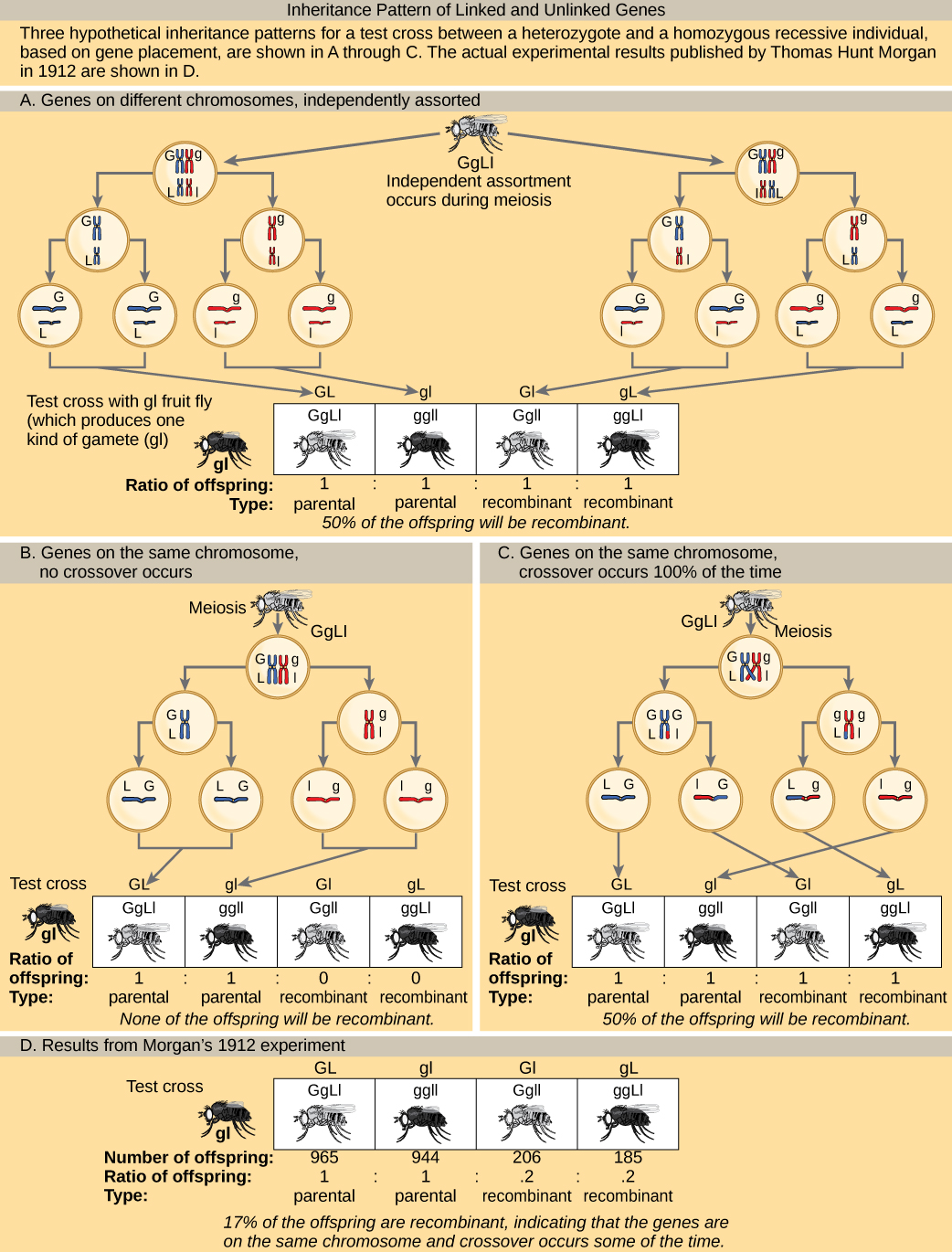
在两个特征(如图所示)的测试十字架中,重组后代的预测频率能否达到 60%? 为什么或者为什么不呢?
遗传图谱
Janssen 没有技术来演示 crossing over,因此它仍然是一个抽象的概念,没有被广泛接受。 科学家们认为 chiasmata 是突触的一种变体,他们无法理解染色体如何会断裂并重新连接。 然而,数据清楚地表明,联系并不总是发生。 最终,花了一位年轻的本科生和一个 “通宵达旦” 的人才用数学方法阐明了联系和重组的问题。
1913年,摩根实验室的学生阿尔弗雷德·斯特特万特从实验室的研究人员那里收集了结果,并有一天晚上带他们回家仔细考虑。 第二天早上,他创建了第一张 “染色体图谱”,这是染色体上基因顺序和相对距离的线性表示(图 13.1.3)。
艺术连接
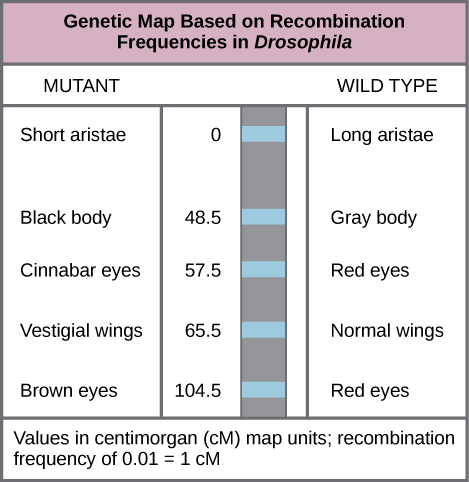
以下陈述中哪一项是正确的?
- 体色和红色/朱砂眼等位基因的重组比翅膀长度和 aristae 长度等位基因的重组更频繁。
- 体色和 aristae 长度等位基因的重组比红/棕眼等位基因和 aristae 长度等位基因的重组更频繁。
- 灰色/黑色体色和长/短 aristae 等位基因不会重组。
- 红/棕眼和长/短 aristae 等位基因的重组比翅膀长度和体色等位基因的重组更频繁。
如图所示\(\PageIndex{3}\),通过使用重组频率预测遗传距离,可以推断出 2 号染色体上基因的相对顺序。 显示的值表示以 centimorgans (cM) 为单位的地图距离,对应于重组频率(以百分比为单位)。 因此,体色和翅膀大小的基因相距为65.5 − 48.5 = 17 cM,这表明这些基因的母系和父系等位基因平均在17%的后代中重组。
为了构建染色体图谱,Sturtevant 假设基因是在线状染色体上连续排序的。 他还假设,在染色体长度的任何地方,两条同源染色体之间重组的发生率可能相等。 根据这些假设,Sturtevant假设染色体上相距较远的等位基因在减数分裂期间更有可能分离,这仅仅是因为存在更大的重组区域。 相反,染色体上彼此接近的等位基因很可能会一起遗传。 两个等位基因(即它们的重组频率)之间的平均交叉次数与它们之间的遗传距离相关,相对于该染色体上其他基因的位置。 考虑到上面的 aaBB 和 aabb 之间的交叉示例,重组频率可以计算为 50/1000 = 0.05。 也就是说,基因 a/a 和 b/ b 之间发生交叉的可能性为 0.05,或 5%。 这样的结果表明这些基因之间存在明确的联系,但它们之间的距离足够远,偶尔会发生交叉。 Sturtevant 将他的遗传图谱划分为地图单位或 c entimorgans (cM),其中 0.01 的重组频率对应于 1 cM。
通过在线性图谱中表示等位基因,Sturtevant提出,当基因位于不同的染色体上或基因在同一染色体上分离得很远时,基因的范围可以从完美连接(重组频率= 0)到完全脱钩(重组频率= 0.5)不等。 完全未连接的基因对应于孟德尔预测的在双杂交叉中独立分组的频率。 重组频率为0.5表示50%的后代是重组体,另外50%是亲本类型。 也就是说,每种类型的等位基因组合都以相等的频率表示。 这种表示法使 Sturtevant 能够加法计算同一染色体上几个基因之间的距离。 但是,随着遗传距离接近0.50,他的预测变得不那么准确了,因为尚不清楚基因在同一条染色体上还是不同染色体上相距很远。
1931年,芭芭拉·麦克林托克和哈丽雅特·克雷顿展示了玉米植物中同源染色体的交叉。 几周后,柯特·斯特恩用显微镜证明了果蝇中的同源重组。 斯特恩观察到几种 X 连锁表型,这些表型与结构上不寻常且不同的 X 染色体对有关,其中一个 X 缺少一个很小的末端片段,另一个 X 与 Y 染色体的一段融合在一起。 通过交叉苍蝇,观察它们的后代,然后可视化后代的染色体,斯特恩证明每当后代等位基因组合偏离任一亲本组合时,都会相应地交换 X 染色体片段。 使用具有不同结构的 X 染色体的突变苍蝇是观察重组产物的关键,因为 DNA 测序和其他分子工具尚不可用。 现在众所周知,同源染色体经常在减数分裂中交换片段,在精确的位置相互断开和重新连接它们的 DNA。

孟德尔的映射特征
同源重组是一种常见的遗传过程,但孟德尔从未观察到过。 如果他同时调查了关联和非关联基因,那么在概率计算的基础上创建统一的数据模型就会困难得多。 此后,研究人员将孟德尔调查的七个特征映射到豌豆植物基因组的七条染色体上,他们证实,他检查的所有基因要么位于不同的染色体上,要么相距足够远,因此在统计学上没有关联。 有人认为孟德尔非常幸运能只选择未连接的基因,而另一些人则质疑孟德尔是否丢弃了任何表明存在关联的数据。 无论如何,孟德尔一直观察到独立的分类,因为他检查了实际上没有关联的基因。
摘要
萨顿和博维里提出的染色体遗传理论指出,染色体是遗传的载体。 孟德尔遗传学和基因连锁都不完全准确;相反,染色体行为涉及分离、独立分类,偶尔还涉及连锁反应。 Sturtevant设计了一种评估重组频率的方法,并根据基因之间中间区域的平均交叉次数推断出染色体上相关基因的相对位置和距离。 Sturtevant 正确地假定基因在染色体上按顺序排列,同系物之间的重组可能发生在染色体上的任何地方,可能性相同。 连锁导致同一染色体上的等位基因一起遗传,而同源重组则将等位基因偏向于独立分类的遗传模式。
艺术联系
图\(\PageIndex{2}\): In a test cross for two characteristics such as the one shown here, can the predicted frequency of recombinant offspring be 60 percent? Why or why not?
- Answer
-
No. The predicted frequency of recombinant offspring ranges from 0% (for linked traits) to 50% (for unlinked traits).
Figure \(\PageIndex{3}\): Which of the following statements is true?
- Recombination of the body color and red/cinnabar eye alleles will occur more frequently than recombination of the alleles for wing length and aristae length.
- Recombination of the body color and aristae length alleles will occur more frequently than recombination of red/brown eye alleles and the aristae length alleles.
- Recombination of the gray/black body color and long/short aristae alleles will not occur.
- Recombination of the red/brown eye and long/short aristae alleles will occur more frequently than recombination of the alleles for wing length and body color.
- Answer
-
D
Glossary
- centimorgan (cM)
- (also, map unit) relative distance that corresponds to a recombination frequency of 0.01
- Chromosomal Theory of Inheritance
- theory proposing that chromosomes are the vehicles of genes and that their behavior during meiosis is the physical basis of the inheritance patterns that Mendel observed
- homologous recombination
- process by which homologous chromosomes undergo reciprocal physical exchanges at their arms, also known as crossing over
- nonparental (recombinant) type
- progeny resulting from homologous recombination that exhibits a different allele combination compared with its parents
- parental types
- progeny that exhibits the same allelic combination as its parents
- recombination frequency
- average number of crossovers between two alleles; observed as the number of nonparental types in a population of progeny