
8.3: תמלול אוקריוטי
- Page ID
- 208475

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)התעתיק באיקריוטים מסובך יותר, אך עוקב אחר אותם רעיונות כלליים. רצפי האמרגן מגוונים הרבה יותר הן במיקום (ביחס לאתר ההתחלה) והן בגודל. כפי שנראה בפרק הבא, לגנים אוקריוטיים יש הרבה יותר מרכיבי בקרה המסדירים את הביטוי שלהם מאשר לגנים פרוקריוטיים. לא רק שיש יותר רכיבי בקרה, יש גם יותר פולימראזות RNA, המשרתות פונקציות סלולריות ספציפיות שונות. ברור שהפונקציה והמיקום הרחב של כל פולימראזות ה- RNA זהים: קרא תבנית DNA ותמלל עותק RNA שלה; ומכיוון שה- DNA נמצא רק בגרעין, כך גם הפולימראזות. עם זאת, הפולימראזות נבדלות בדיוק באילו סוגי RNA הם מייצרים. RNA פולי-מרז I מתמחה בייצור פרה-rRNA (rRNA = RNA ריבוזומלי). ה- pre-rRNA מבוקע לאחר תעתיק ומשולב בריבוזומים. מכיוון שהריבוזומים מורכבים בגרעין, זהו החלק של הגרעין בו מרוכז רוב ה- RNA פולימראז I. RNA פולימראז III מייצר גם RNA (5S) המשולב בריבוזום. זה גם מייצר RNA לא מתורגמים אחרים כגון tRNAs ומגוון RNAs גרעיניים קטנים. פולימראז ה-RNA היחיד שיוצר את ה-RNA הניתן לתרגום (mRNA, או RNA שליח) שרוב האנשים חושבים עליו כאשר מתייחסים ל-RNA באופן כללי, הוא RNA פולימראז II. זהו פולימראז ה- RNA המייצר פרה-mRNA, שאחרי עיבוד מסוים הופך ל-mRNA, מועבר אל מחוץ לגרעין, ולבסוף מתורגם לחלבונים. כל פולימראזות ה- RNA האוקריוטיות מורכבות משתי יחידות משנה גדולות, המקבילות בערך ליחידות המשנה β ו- β של RNAP פרוקריוטי, אך במקום רק שלוש או ארבע יחידות משנה אחרות, יש יותר מתריסר יחידות משנה קטנות יותר להולואנזימים של RNA פולימראז האוקריוטי.
התחלת התמלול היא גם הרבה יותר מסובכת. לא רק שיש מגוון גדול במקדמים המוכרים על ידי RNAP II, הן RNAP I והן RNAP III מזהים מקדמים בעלי מאפיינים מבניים מסוימים.
פולימראזות ה-RNA האוקריוטיות נקראו I, II ו-III בהתבסס על סדר הפליטה שלהן מטיהור כרומטוגרפיה של חילופי יונים. הם נבדלים חלקית גם על ידי רגישותם ל-α-amanitin ולרעלים הקשורים לפטריות ממשפחת האמטוקסין. RNAP I (ו- RNAP פרוקריוטי) אינו רגיש לרעלים אלה, RNAP III רגיש במקצת (K d ~ 10 -6 M), ו- RNAP II רגיש מאוד (K d ~ 10 -8 M). רעלים אלה פועלים על ידי קשירה לאתר בשסע ה- RNA-DNA ומפריעים לטרנסלוקציה של ה- RNA. כלומר, אין שום בעיה לייבא נוקלאוטיד או לחבר אותו ל- RNA החדש, אך גדיל ה- RNA אינו יכול לנוע באתר הפעיל ולאפשר להוסיף את הנוקלאוטיד הבא.
אחד ממקדמי ה- RNAP II האוקריוטיים הנפוצים ביותר הוא תיבת TATA, על שם המוטיב השמור ביותר המגדיר אותה. למרות שהיא נראית דומה לתיבת Pribnow בפרוקריוטים, היא ממוקמת בדרך כלל במעלה הזרם מאתר ההתחלה, ומיקומה משתנה הרבה יותר. בעוד שתיבת Pribnow ממוקמת ב -10, תיבת TATA עשויה להיות ממוקמת קרוב יותר ל -30 +/- 4. כמו כן, במקום רק גורם סיגמא לזיהוי הפרומוטור בשילוב עם אנזים הליבה של הפולימראז, האמרגן האיקריוטי מזוהה על ידי קומפלקס רב-תת-יחידות הנקרא גורם שעתוק IID (TFIID). TFIID מורכב מחלבון קושר TATA (TBP) ומספר גורמים הקשורים ל- TBP (TAFs).
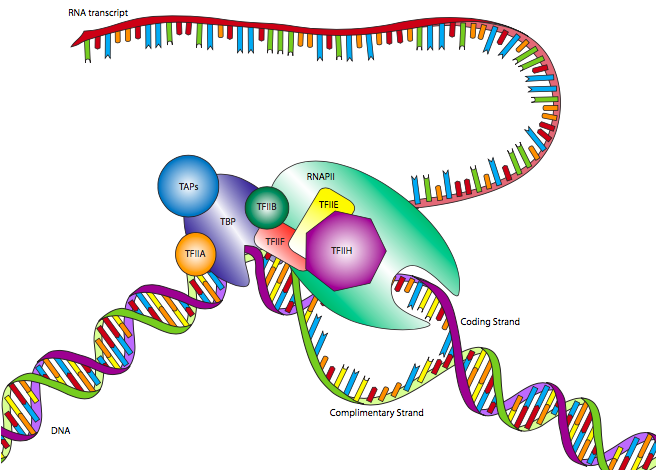
קישור זה של האמרגן על ידי TFIID מתרחש ללא תלות ב-RNA Polymerase II, ולמעשה, RNAP II לא יתחבר ל-TFIID בשלב זה. לאחר ש-TFIID קשר את תיבת ה-TATA, שני גורמי שעתוק נוספים, TFIIA ו-TFIIB, מתחברים ל-TFIID כמו גם ל-DNA הסמוך, ומייצבים את המתחם. TFIIF מתחבר ל-TFIID ו-TFIIB כדי לאפשר עגינה של ה-RNA פולימראז II. המתחם עדיין לא מוכן להתחיל בתעתיק: נדרשים שני גורמים נוספים. TFIIE קושר TFIIF ו-RNAP II, ולבסוף, TFIIH מתחבר ל-RNAP II, ומספק פעילות הליקאז הדרושה כדי לחטט את שני גדילי ה-DNA ולאפשר לפולימראז לקרוא אחד מהם. ל- TFIIH יש גם פעילות אנזימטית חשובה נוספת: זהו גם קינאז סרין המזרחן את התחום הקרבוקסיל-טרמינלי (CTD) של RNA פולימראז II. ישנם מספר סרינים ב-CTD, וככל שהם מזרחנים ברצף, ה-CTD משתרע כמו זנב (טעון שלילי) ועוזר לקדם הפרדה בין ה-RNAP II ל-TFIID/מקדם.
התארכות גדיל ה- RNA באוקריוטים דומה מאוד לזו שבפרוקריוטים עם ההבדל הברור כי שעתוק מתרחש בגרעין ולא בציטופלזמה. לפיכך, בפרוקריוטים, ניתן להשתמש ב-RNA לתרגום חלבונים גם כשהוא עדיין מתועתק מה-DNA! באאוקריוטים המצב מורכב יותר באופן משמעותי: ישנם מספר אירועים שלאחר תעתיק (מכסת קצה 5', פוליאדנילציה 3', ולעתים קרובות שחבור RNA) שחייבים להתרחש לפני שה-RNA מוכן להובלה אל מחוץ לגרעין וזמין לתרגום בציטופלזמה.
סיום התעתיק האיקריוטי אינו מתואר היטב בכתיבה זו. נראה כי RNAP I דורש גורם סיום מחייב DNA, שאינו מקביל לגורם Rho הפרוקריוטי, שהוא חלבון קושר RNA. RNAP III מסיים את השעתוק ללא כל גורם חיצוני, וסיום זה מתרחש בדרך כלל לאחר הוספת סדרה של שאריות אורידין. עם זאת, נראה שהוא אינו משתמש במבנה לולאת סיכת השיער המצוי בתעתיק חיידקי בלתי תלוי ב-rho. סיום תמלילי RNAP II המקודדים לחלבון מקושר לקומפלקס אנזים שגם מבקע חלק מקצה ה-3' של ה-RNA, ומוסיף זנב פולי-A. עם זאת, לא ברור כיצד קומפלקס הפוליאדנילציה מעורב בקביעת נקודת סיום השעתוק, שיכולה להיות מעל 1000 נוקלאוטידים מעבר לאתר פולי-A (למשל הגן β-globin ב Mus musculus). עם סיום ושחרור מה- RNAP II ו- DNA התבנית, ה- RNA ידוע בתור התמליל העיקרי, אך חייב לעבור עיבוד לאחר תעתיק לפני שהוא RNA שליח בוגר (mRNA) מוכן לייצוא לציטופלזמה ומשמש לכוון תרגום.