
10.2: Estrutura e função do DNA
- Page ID
- 181243

Objetivos de
- Descreva a estrutura bioquímica dos desoxirribonucleotídeos
- Identifique os pares de bases usados na síntese de desoxirribonucleotídeos
- Explique por que a dupla hélice do DNA é descrita como antiparalela
Em Metabolismo microbiano, discutimos três classes de macromoléculas: proteínas, lipídios e carboidratos. Neste capítulo, discutiremos uma quarta classe de macromoléculas: ácidos nucléicos. Como outras macromoléculas, os ácidos nucléicos são compostos por monômeros, chamados nucleotídeos, que são polimerizados para formar grandes cadeias. Cada fita de ácido nucléico contém certos nucleotídeos que aparecem em uma determinada ordem dentro da fita, chamada de sequência base. A sequência base do ácido desoxirribonucléico (DNA) é responsável por transportar e reter a informação hereditária em uma célula. Em Mecanismos de Genética Microbiana, discutiremos em detalhes as maneiras pelas quais o DNA usa sua própria sequência de bases para direcionar sua própria síntese, bem como a síntese de RNA e proteínas, o que, por sua vez, dá origem a produtos com estrutura e função diversas. Nesta seção, discutiremos a estrutura e função básicas do DNA.
Nucleotídeos de DNA
Os blocos de construção dos ácidos nucléicos são os nucleotídeos. Os nucleotídeos que compõem o DNA são chamados de desoxirribonucleotídeos. Os três componentes de um desoxirribonucleotídeo são um açúcar de cinco carbonos chamado desoxirribose, um grupo fosfato, e uma base nitrogenada, uma estrutura em anel contendo nitrogênio que é responsável pelo emparelhamento complementar de bases entre cadeias de ácido nucléico (Figura\(\PageIndex{1}\)). Os átomos de carbono da desoxirribose de cinco carbonos são numerados 1, 2, 3, 4e 5( 1é lido como “um primo”). Um nucleosídeo compreende o açúcar de cinco carbonos e a base nitrogenada.
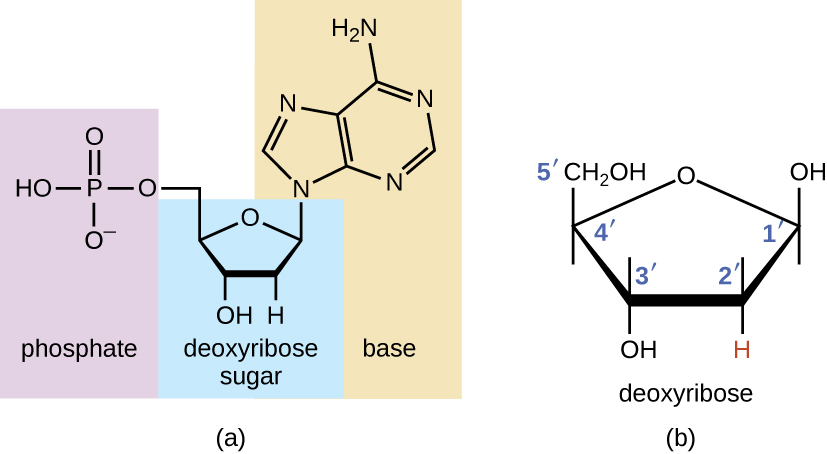
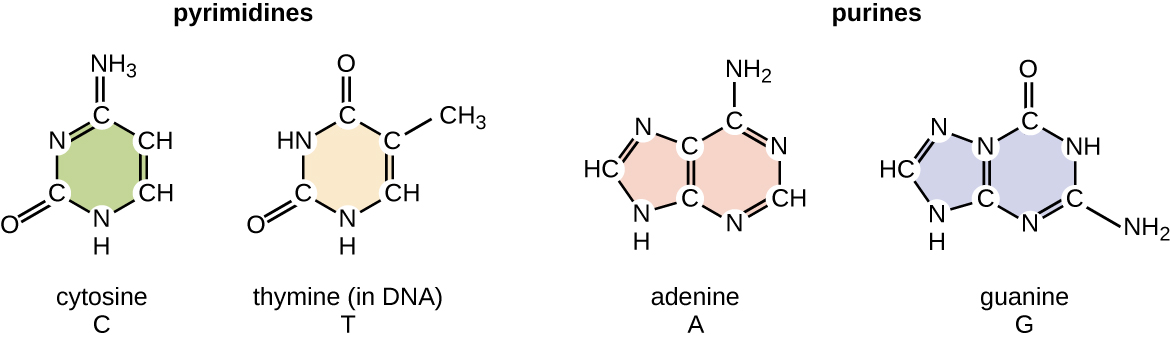
O desoxirribonucleotídeo é nomeado de acordo com as bases nitrogenadas (Figura\(\PageIndex{2}\)). As bases nitrogenadas adenina (A) e guanina (G) são as purinas; elas têm uma estrutura de anel duplo com um anel de seis carbonos fundido a um anel de cinco carbonos. As pirimidinas, citosina (C) e timina (T), são bases nitrogenadas menores que têm apenas uma estrutura de anel de seis carbonos.
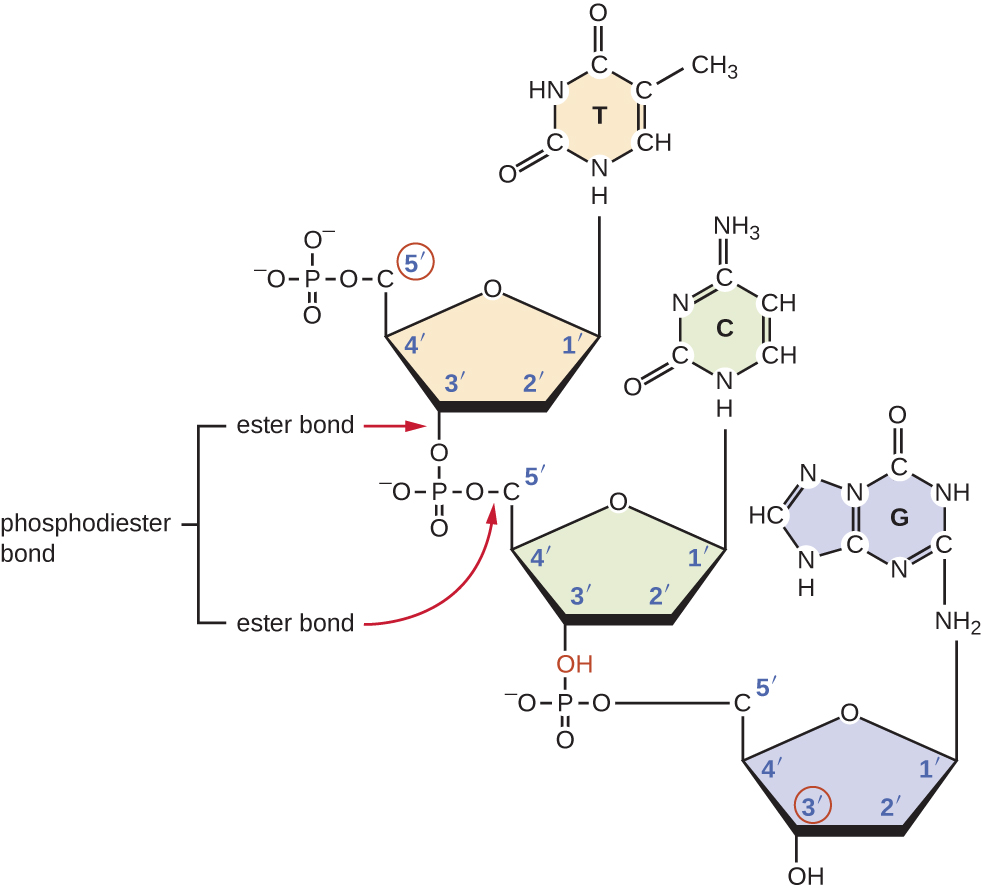
Os trifosfatos de nucleosídeos individuais se combinam entre si por ligações covalentes conhecidas como ligações 5'-3fosfodiéster, ou ligações pelas quais o grupo fosfato ligado ao carbono 5do açúcar de um nucleotídeo se liga ao grupo hidroxila do carbono 3do açúcar do próximo nucleotídeo. A ligação fosfodiéster entre nucleotídeos forma a espinha dorsal açúcar-fosfato, a estrutura alternada de açúcar-fosfato que compõe a estrutura de uma fita de ácido nucléico (Figura\(\PageIndex{3}\)). Durante o processo de polimerização, são usados trifosfatos desoxinucleotídicos (dNTP). Para construir a espinha dorsal de açúcar-fosfato, os dois fosfatos terminais são liberados do dNTP como um pirofosfato. A fita resultante de ácido nucléico tem um grupo fosfato livre na extremidade de carbono 5e um grupo hidroxila livre na extremidade de carbono 3. Os dois grupos fosfato não utilizados do nucleotídeo trifosfato são liberados como pirofosfato durante a formação da ligação fosfodiéster. O pirofosfato é posteriormente hidrolisado, liberando a energia usada para impulsionar a polimerização de nucleotídeos.
Exercício\(\PageIndex{1}\)
O que se entende pelas extremidades 5e 3de uma fita de ácido nucléico?
Descobrindo a dupla hélice
No início da década de 1950, evidências consideráveis haviam se acumulado indicando que o DNA era o material genético das células, e agora a corrida começou a descobrir sua estrutura tridimensional. Nessa época, o bioquímico austríaco Erwin Chargaff 1 (1905—2002) examinou o conteúdo do DNA em diferentes espécies e descobriu que adenina, timina, guanina e citosina não eram encontradas em quantidades iguais e que variavam de espécie para espécie, mas não entre indivíduos da mesma. espécie. Ele descobriu que a quantidade de adenina era muito próxima da quantidade de timina, e a quantidade de citosina estava muito próxima de igualar a quantidade de guanina, ou A = T e G = C. Essas relações também são conhecidas como regras de Chargaff.
Outros cientistas também estavam explorando ativamente esse campo em meados do século XX. Em 1952, o cientista americano Linus Pauling (1901—1994) foi o principal químico estrutural do mundo e o favorito para resolver a estrutura do DNA. Pauling já havia descoberto a estrutura das hélices da proteína α, usando difração de raios-X, e, com base em imagens de difração de raios-X do DNA feitas em seu laboratório, ele propôs um modelo de DNA de fita tripla. 2 Ao mesmo tempo, a pesquisadora britânica Rosalind Franklin (1920—1958) e seu aluno de graduação R.G. Gosling também estavam usando a difração de raios-X para entender a estrutura do DNA (Figura\(\PageIndex{4}\)). Foi a experiência científica de Franklin que resultou na produção de imagens de difração de raios X do DNA mais bem definidas que mostrariam claramente a estrutura geral de dupla hélice do DNA.

James Watson (1928—), um cientista americano, e Francis Crick (1916—2004), um cientista britânico, estavam trabalhando juntos na década de 1950 para descobrir a estrutura do DNA. Eles usaram as regras de Chargaff e as imagens de difração de raios-X de fibras de DNA de Franklin e Wilkins para juntar a combinação purina-pirimidina da molécula de DNA de dupla hélice (Figura\(\PageIndex{5}\)). Em abril de 1953, Watson e Crick publicaram seu modelo da dupla hélice de DNA na Nature. 3 A mesma edição também incluiu artigos de Wilkins e colegas, 4 bem como de Franklin e Gosling, 5 cada um descrevendo diferentes aspectos da estrutura molecular do DNA. Em 1962, James Watson, Francis Crick e Maurice Wilkins receberam o Prêmio Nobel de Fisiologia e Medicina. Infelizmente, até então Franklin havia morrido e os prêmios Nobel na época não foram concedidos postumamente. No entanto, o trabalho continuou no aprendizado sobre a estrutura do DNA. Em 1973, Alexander Rich (1924—2015) e seus colegas conseguiram analisar cristais de DNA para confirmar e elucidar melhor a estrutura do DNA. 6

Exercício\(\PageIndex{2}\)
Quais cientistas recebem a maior parte do crédito por descreverem a estrutura molecular do DNA?
Estrutura do DNA
Watson e Crick propuseram que o DNA é feito de duas fitas que são torcidas uma em volta da outra para formar uma hélice destra. As duas fitas de DNA são antiparalelas, de forma que a extremidade 3de uma fita está voltada para a extremidade 5da outra (Figura\(\PageIndex{6}\)). A extremidade 3de cada fita tem um grupo hidroxila livre, enquanto a extremidade 5de cada fita tem um grupo fosfato livre. O açúcar e o fosfato dos nucleotídeos polimerizados formam a espinha dorsal da estrutura, enquanto as bases nitrogenadas são empilhadas em seu interior. Essas bases nitrogenadas no interior da molécula interagem umas com as outras, emparelhando bases.
A análise dos padrões de difração do DNA determinou que existem aproximadamente 10 bases por turno no DNA. O espaçamento assimétrico das espinhas dorsais de açúcar-fosfato gera ranhuras maiores (onde a coluna vertebral está distante) e ranhuras menores (onde a espinha dorsal está próxima) (Figura\(\PageIndex{6}\)). Esses sulcos são locais onde as proteínas podem se ligar ao DNA. A ligação dessas proteínas pode alterar a estrutura do DNA, regular a replicação ou regular a transcrição do DNA em RNA.
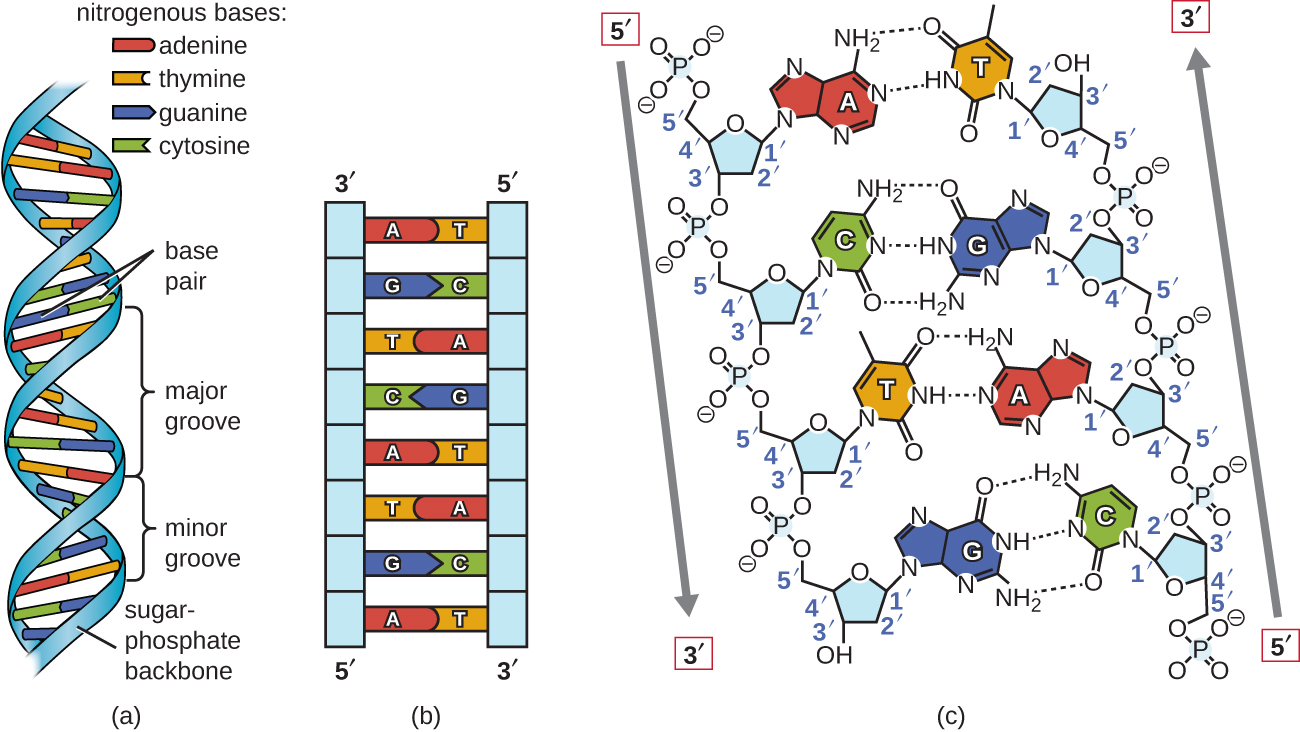
O emparelhamento de bases ocorre entre uma purina e uma pirimidina. No DNA, a adenina (A) e a timina (T) são pares de bases complementares, e a citosina (C) e a guanina (G) também são pares de bases complementares, explicando as regras de Chargaff (Figura\(\PageIndex{7}\)). Os pares de bases são estabilizados por ligações de hidrogênio; adenina e timina formam duas ligações de hidrogênio entre eles, enquanto a citosina e a guanina formam três ligações de hidrogênio entre eles.
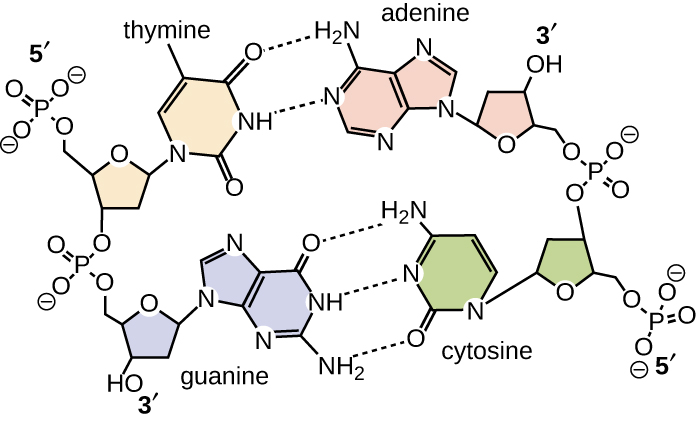
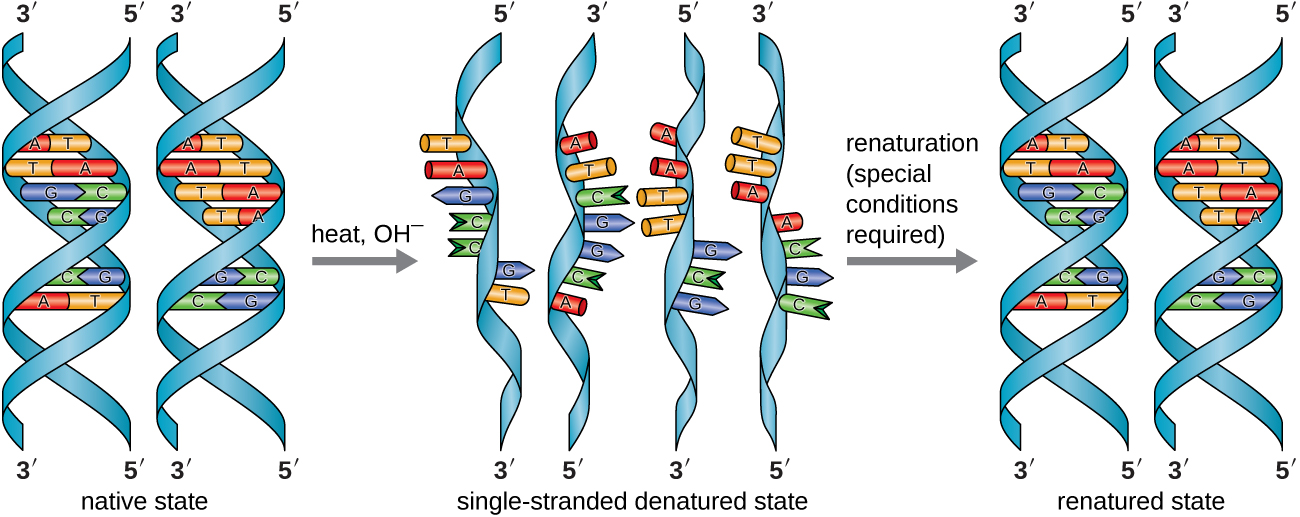
Em laboratório, expor as duas fitas de DNA da dupla hélice a altas temperaturas ou a certos produtos químicos pode quebrar as ligações de hidrogênio entre bases complementares, separando assim as fitas em duas fitas simples separadas de DNA (DNA de fita simples [ssDNA]). Esse processo é chamado de desnaturação do DNA e é análogo à desnaturação de proteínas, conforme descrito em Proteínas. As fitas de ssDNA também podem ser reunidas novamente como DNA de fita dupla (dsDNA), por meio de recozimento ou renaturação, resfriando ou removendo os desnaturantes químicos, permitindo que essas ligações de hidrogênio se reformem. A capacidade de manipular artificialmente o DNA dessa maneira é a base de várias técnicas importantes em biotecnologia (Figura\(\PageIndex{8}\)). Devido à ligação adicional de hidrogênio entre o par de bases C = G, o DNA com alto teor de GC é mais difícil de desnaturar do que o DNA com menor teor de GC.
Veja uma animação sobre a estrutura do DNA no Centro de Aprendizagem de DNA para saber mais.
Exercício\(\PageIndex{3}\)
Quais são os dois pares de bases complementares do DNA e como eles se unem?
Função do DNA
O DNA armazena as informações necessárias para construir e controlar a célula. A transmissão dessas informações das células-mãe para a filha é chamada de transferência vertical de genes e ocorre por meio do processo de replicação do DNA. O DNA é replicado quando uma célula faz uma cópia duplicada de seu DNA e, em seguida, a célula se divide, resultando na distribuição correta de uma cópia do DNA para cada célula resultante. O DNA também pode ser degradado enzimaticamente e usado como fonte de nucleosídeos e nucleotídeos para a célula. Ao contrário de outras macromoléculas, o DNA não desempenha um papel estrutural nas células.
Exercício\(\PageIndex{4}\)
Como o DNA transmite informações genéticas para a prole?
Abrindo o caminho para mulheres nas profissões da ciência e da saúde
Historicamente, as mulheres têm sido sub-representadas nas ciências e na medicina e, muitas vezes, suas contribuições pioneiras passaram relativamente despercebidas. Por exemplo, embora Rosalind Franklin tenha realizado os estudos de difração de raios-X demonstrando a estrutura dupla helicoidal do DNA, foram Watson e Crick que ficaram famosos por essa descoberta, com base em seus dados. Ainda há uma grande controvérsia sobre se a aquisição de seus dados foi apropriada e se os conflitos de personalidade e o preconceito de gênero contribuíram para o atraso no reconhecimento de suas contribuições significativas. Da mesma forma, Barbara McClintock fez um trabalho pioneiro na genética do milho (milho) das décadas de 1930 a 1950, descobrindo transposons (genes saltadores), mas só foi reconhecida muito mais tarde, recebendo o Prêmio Nobel de Fisiologia ou Medicina em 1983 (Figura\(\PageIndex{9}\)).
Hoje, as mulheres ainda permanecem sub-representadas em muitos campos da ciência e da medicina. Enquanto mais da metade dos cursos de graduação em ciências são concedidos a mulheres, apenas 46% dos graus de doutorado em ciências são concedidos a mulheres. Na academia, o número de mulheres em cada nível de progressão na carreira continua diminuindo, com mulheres ocupando menos de um terço dos cargos de cientistas de nível de doutorado em cargos permanentes e menos de um quarto dos cargos de professor titular em faculdades e universidades de 4 anos. 7 Mesmo nas profissões da saúde, como em quase todas as outras áreas, as mulheres geralmente estão sub-representadas em muitas carreiras médicas e ganham significativamente menos do que os homens, conforme mostrado em um estudo de 2013 publicado pelo Journal of the American Medical Association. 8
Por que essas disparidades continuam existindo e como quebramos esses ciclos? A situação é complexa e provavelmente resulta da combinação de vários fatores, incluindo como a sociedade condiciona os comportamentos das meninas desde tenra idade e apóia seus interesses, tanto profissional quanto pessoalmente. Alguns sugeriram que as mulheres não pertencem ao laboratório, incluindo o ganhador do Prêmio Nobel Tim Hunt, cujos comentários públicos de 2015 sugerindo que as mulheres são muito emocionais para a ciência 9 foram recebidos com ampla condenação.
Talvez as meninas devessem receber mais apoio desde tenra idade nas áreas de ciências e matemática (Figura\(\PageIndex{9}\)). Os programas de ciência, tecnologia, engenharia e matemática (STEM) patrocinados pela Associação Americana de Mulheres Universitárias (AAUW) 10 e pela Administração Nacional de Aeronáutica e Espaço (NASA) 11 são excelentes exemplos de programas que oferecem esse apoio. As contribuições das mulheres na ciência devem ser divulgadas mais amplamente ao público, e o marketing direcionado a meninas deve incluir mais imagens de cientistas e profissionais médicos bem-sucedidos histórica e profissionalmente, incentivando todas as mentes jovens brilhantes, incluindo meninas e mulheres, a buscarem carreiras em ciência e medicina.

Foco clínico: Parte 2
Com base em seus sintomas, o médico de Alex suspeita que ele esteja sofrendo de uma doença de origem alimentar que ele adquiriu durante suas viagens. As possibilidades incluem infecção bacteriana (por exemplo, E. coli enterotoxigênica, Vibrio cholerae, Campylobacter jejuni, Salmonella), infecção viral (rotavírus ou norovírus) ou infecção por protozoários (Giardia lamblia, Cryptosporidium parvum , ou Entamoeba histolytica).
Seu médico solicita uma amostra de fezes para identificar possíveis agentes causadores (por exemplo, bactérias, cistos) e verificar a presença de sangue porque certos tipos de agentes infecciosos (como C. jejuni, Salmonella e E. histolytica) estão associados à produção de sangue fezes.
A amostra de fezes de Alex não mostrou sangue nem cistos. Após a análise de sua amostra de fezes e com base em seu histórico recente de viagens, o médico do hospital suspeitou que Alex estava sofrendo de diarreia do viajante causada pela E. coli enterotoxigênica (ETEC), o agente causador da diarreia da maioria dos viajantes. Para verificar o diagnóstico e descartar outras possibilidades, o médico de Alex solicitou um teste de laboratório de diagnóstico de sua amostra de fezes para procurar sequências de DNA que codificam fatores de virulência específicos da ETEC. O médico instruiu Alex a beber muitos líquidos para substituir o que ele estava perdendo e o dispensou do hospital.
A ETEC produz vários fatores de virulência codificados por plasmídeo que a tornam patogênica em comparação com a E. coli típica. Isso inclui as toxinas secretadas enterotoxina termo-lábil (LT) e enterotoxina termo-estável (ST), bem como o fator de colonização (CF). Tanto o LT quanto o ST causam a excreção de íons cloreto das células intestinais para o lúmen intestinal, causando uma consequente perda de água das células intestinais, resultando em diarreia. A CF codifica uma proteína bacteriana que ajuda a permitir que a bactéria adira ao revestimento do intestino delgado.
Exercício\(\PageIndex{5}\)
Por que o médico de Alex usou a análise genética em vez do isolamento de bactérias da amostra de fezes ou da coloração direta de Gram somente da amostra de fezes?
Conceitos principais e resumo
- Os ácidos nucléicos são compostos por nucleotídeos, cada um contendo um açúcar pentose, um grupo fosfato e uma base nitrogenada. Os desoxirribonucleotídeos no DNA contêm desoxirribose como açúcar pentose.
- O DNA contém as pirimidinas citosina e timina e as purinas adenina e guanina.
- Os nucleotídeos estão ligados entre si por ligações fosfodiéster entre o grupo 5fosfato de um nucleotídeo e o grupo 3hidroxila de outro. Uma fita de ácido nucléico tem um grupo fosfato livre na extremidade 5e um grupo hidroxila livre na extremidade 3.
- Chargaff descobriu que a quantidade de adenina é aproximadamente igual à quantidade de timina no DNA e que a quantidade de guanina é aproximadamente igual à citosina. Posteriormente, essas relações foram determinadas como decorrentes do emparelhamento de bases complementar.
- Watson e Crick, com base no trabalho de Chargaff, Franklin e Gosling e Wilkins, propuseram o modelo de dupla hélice e o emparelhamento de bases para a estrutura do DNA.
- O DNA é composto por duas cadeias complementares orientadas antiparalelamente uma à outra com as espinhas dorsais do fosfodiéster no exterior da molécula. As bases nitrogenadas de cada fita se enfrentam e as bases complementares de hidrogênio se ligam umas às outras, estabilizando a dupla hélice.
- O calor ou os produtos químicos podem romper as ligações de hidrogênio entre as bases complementares, desnaturando o DNA. O resfriamento ou a remoção de produtos químicos podem levar à renaturação ou recozimento do DNA, permitindo que as ligações de hidrogênio se reformem entre bases complementares.
- O DNA armazena as instruções necessárias para construir e controlar a célula. Essas informações são transmitidas de pais para filhos por meio da transferência vertical de genes.
Notas de pé
- 1 N. Kresge et al. “Regras do Chargaff: o trabalho de Erwin Chargaff.” Journal of Biological Chemistry 280 (2005) :e21.
- 2 L. Pauling, “Uma estrutura proposta para os ácidos nucléicos”. Anais da Academia Nacional de Ciências dos Estados Unidos da América 39 no. 2 (1953) :84—97.
- 3 J.D. Watson, F.H.C. Crick. “Uma estrutura para o ácido nucléico desoxirribose.” Nature 171 nº 4356 (1953) :737—738.
- 4 M.H.F. Wilkins e cols.. “Estrutura molecular dos ácidos nucléicos da desoxipentose”. Nature 171 nº 4356 (1953) :738—740.
- 5 R. Franklin, R. G. Gosling. “Configuração molecular em timonucleato de sódio.” Nature 171 nº 4356 (1953) :740—741.
- 6 R.O. Day et al. “Um fragmento cristalino da dupla hélice: a estrutura do fosfato dinucleosídico guanilil-3',5'-citidina.” Anais da Academia Nacional de Ciências dos Estados Unidos da América 70 no. 3 (1973) :849—853.
- 7 N.H. Wolfinger “Para mulheres cientistas, não há um bom momento para ter filhos”. The Atlantic 29 de julho de 2013. www.theatlantic.com/sexes/arc... ildren/278165/.
- 8 S.A. Seabury e cols.. “Tendências nos ganhos de profissionais de saúde masculinos e femininos nos Estados Unidos, 1987 a 2010.” Jornal da Associação Médica Americana de Medicina Interna 173 nº 18 (2013) :1748—1750.
- 9 E. Chung. “Tim Hunt, sexismo e ciência: o verdadeiro 'problema com as meninas' nos laboratórios.” CBC News Technology and Science, 12 de junho de 2015. http://www.cbc.ca/news/technology/ti...labs-1.3110133. Acessado em 8/4/2016.
- 10 Associação Americana de Mulheres Universitárias. “Construindo um pipeline STEM para meninas e mulheres.” www.aauw.org/what-we-do/stem-education/. Acessado em 10 de junho de 2016.
- 11 Administração Nacional de Aeronáutica e Espaço. “Programas de extensão: iniciativa para mulheres e meninas”. http://women.nasa.gov/outreach-programs/. Acessado em 10 de junho de 2016.