
10.1: Usando a microbiologia para descobrir os segredos da vida
- Page ID
- 181261

Objetivos de
- Descreva a descoberta do ácido nucléico e dos nucleotídeos
- Explique os experimentos históricos que levaram à caracterização do DNA
- Descreva como a microbiologia e os microrganismos têm sido usados para descobrir a bioquímica dos genes
- Explique como os cientistas estabeleceram a ligação entre DNA e hereditariedade
Foco clínico: Parte 1
Alex é um estudante universitário de 22 anos que passou férias em Puerta Vallarta, México, nas férias de primavera. Infelizmente, dois dias depois de voltar para Ohio, ele começou a sentir cólicas abdominais e diarreia aquosa extensa. Por causa de seu desconforto, ele procurou atendimento médico em um grande hospital de Cincinnati nas proximidades.
Exercício\(\PageIndex{1}\)
Quais tipos de infecções ou outras condições podem ser responsáveis?
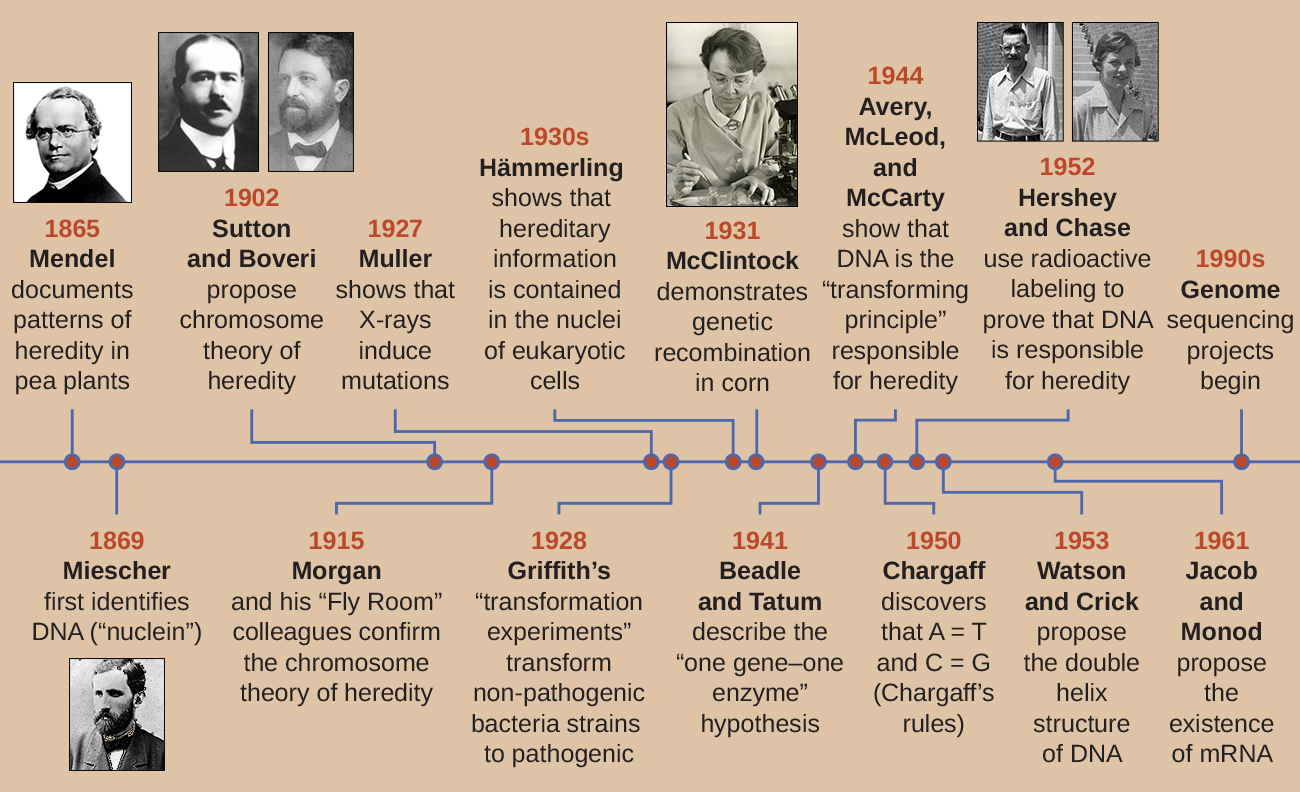
Durante o início do século XX, o DNA ainda não era reconhecido como o material genético responsável pela hereditariedade, pela passagem de características de uma geração para a outra. Na verdade, grande parte da pesquisa foi descartada até meados do século XX. A comunidade científica acreditava, incorretamente, que o processo de herança envolvia uma mistura de características parentais que produzia uma aparência física intermediária na prole; esse processo hipotético parecia estar correto devido ao que conhecemos agora como variação contínua, que resulta da ação de muitos genes para determinar uma característica específica, como a altura humana. Os filhos parecem ser uma “mistura” das características de seus pais quando observamos características que exibem variação contínua. A teoria da herança da mistura afirmava que as características parentais originais foram perdidas ou absorvidas pela mistura na prole, mas agora sabemos que esse não é o caso.
Duas linhas distintas de pesquisa, iniciadas em meados do século XIX, acabaram levando à descoberta e caracterização do DNA e dos fundamentos da genética, a ciência da hereditariedade. Essas linhas de pesquisa começaram a convergir na década de 1920, e a pesquisa usando sistemas microbianos acabou resultando em contribuições significativas para elucidar a base molecular da genética.
Descoberta e caracterização do DNA
A compreensão moderna do DNA evoluiu da descoberta do ácido nucléico para o desenvolvimento do modelo de dupla hélice. Na década de 1860, Friedrich Miescher (1844-1895), médico de profissão, foi a primeira pessoa a isolar substâncias químicas ricas em fósforo dos leucócitos (glóbulos brancos) do pus em bandagens usadas de uma clínica cirúrgica local. Ele chamou essas substâncias químicas (que eventualmente seriam conhecidas como RNA e DNA) de “nucleína” porque estavam isoladas dos núcleos das células. Seu aluno Richard Altmann (1852-1900) posteriormente o chamou de “ácido nucléico” 20 anos depois, quando descobriu a natureza ácida da nucleína. Nas últimas duas décadas do século XIX, o bioquímico alemão Albrecht Kossel (1853-1927) isolou e caracterizou as cinco bases nucleotídicas diferentes que compõem o ácido nucléico. São adenina, guanina, citosina, timina (no DNA) e uracil (no RNA). Kossell recebeu o Prêmio Nobel de Fisiologia ou Medicina em 1910 por seu trabalho com ácidos nucléicos e por seu considerável trabalho com proteínas, incluindo a descoberta da histidina.
Fundamentos da Genética
Apesar da descoberta do DNA no final do século XIX, os cientistas não fizeram a associação com a hereditariedade por muitas décadas. Para fazer essa conexão, cientistas, incluindo vários microbiologistas, realizaram muitos experimentos em plantas, animais e bactérias.
Plantas de ervilha de Mendel
Enquanto Miescher estava isolando e descobrindo o DNA na década de 1860, o monge e botânico austríaco Johann Gregor Mendel (1822-1884) estava experimentando ervilhas, demonstrando e documentando padrões básicos de herança, agora conhecidos como leis de Mendel.
Em 1856, Mendel começou sua pesquisa de uma década sobre padrões de herança. Ele usou a ervilha diploide, Pisum sativum, como seu principal sistema modelo porque ela se autofertiliza naturalmente e é altamente endogânica, produzindo linhagens de plantas de ervilha “verdadeiras” - plantas que sempre produzem filhotes que se parecem com a mãe. Ao fazer experiências com plantas de ervilha verdadeiras, Mendel evitou o aparecimento de características inesperadas na prole que poderiam ocorrer se ele usasse plantas que não fossem verdadeiras. Mendel realizou hibridizações, que envolvem o acasalamento de dois indivíduos reprodutores verdadeiros (geração P) com características diferentes, e examinou as características de seus filhotes (primeira geração filial, F 1), bem como os filhos de autofertilização da geração F 1 (segunda geração filial, F (2) (Figura 1\(\PageIndex{1}\)).
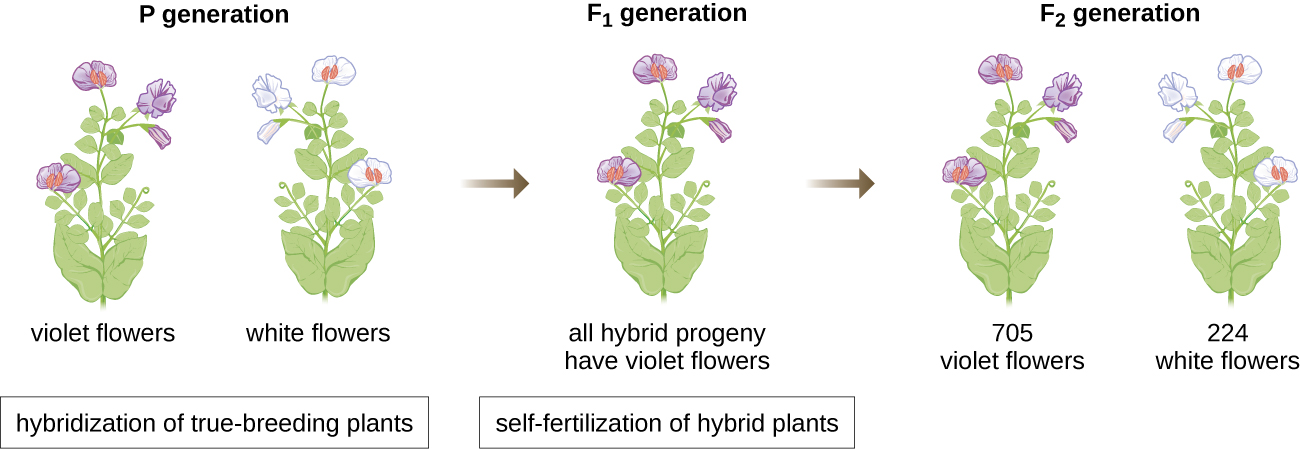
Em 1865, Mendel apresentou os resultados de seus experimentos com quase 30.000 plantas de ervilha à sociedade local de história natural. Ele demonstrou que as características são transmitidas fielmente de pais para filhos, independentemente de outras características. Em 1866, ele publicou seu trabalho, “Experiments in Plant Hybridization”, 1 no Proceedings of the Natural History Society of Brünn. O trabalho de Mendel passou praticamente despercebido pela comunidade científica, que acreditava, incorretamente, na teoria da mistura de traços em variação contínua.
Ele não foi reconhecido por suas contribuições científicas extraordinárias durante sua vida. Na verdade, foi somente em 1900 que seu trabalho foi redescoberto, reproduzido e revitalizado por cientistas à beira de descobrir a base cromossômica da hereditariedade.
A teoria cromossômica da herança
Mendel realizou seus experimentos muito antes de os cromossomos serem visualizados ao microscópio. No entanto, com o aprimoramento das técnicas microscópicas durante o final do século XIX, os biólogos celulares puderam manchar e visualizar estruturas subcelulares com corantes e observar suas ações durante a meiose. Eles foram capazes de observar cromossomos se replicando, condensando-se de uma massa nuclear amorfa em corpos distintos em forma de X e migrando para pólos celulares separados. A especulação de que os cromossomos poderiam ser a chave para entender a hereditariedade levou vários cientistas a examinar as publicações de Mendel e reavaliar seu modelo em termos do comportamento dos cromossomos durante a mitose e a meiose.
Em 1902, Theodor Boveri (1862-1915) observou que nos ouriços-do-mar, os componentes nucleares (cromossomos) determinavam o desenvolvimento embrionário adequado. Nesse mesmo ano, Walter Sutton (1877—1916) observou a separação dos cromossomos em células-filhas durante a meiose. Juntas, essas observações levaram ao desenvolvimento da Teoria Cromossômica da Herança, que identificou os cromossomos como o material genético responsável pela herança mendeliana.
Apesar das correlações convincentes entre o comportamento dos cromossomos durante a meiose e as observações de Mendel, a Teoria Cromossômica da Herança foi proposta muito antes de haver qualquer evidência direta de que as características eram transmitidas nos cromossomos. Thomas Hunt Morgan (1866-1945) e seus colegas passaram vários anos realizando cruzamentos com a mosca-das-frutas, Drosophila melanogaster. Eles realizaram observações microscópicas meticulosas dos cromossomos da mosca e correlacionaram essas observações com as características da mosca resultantes. Seu trabalho forneceu a primeira evidência experimental para apoiar a Teoria Cromossômica da Herança no início dos anos 1900. Em 1915, Morgan e seus colegas do “Fly Room” publicaram O Mecanismo da Hereditariedade Mendeliana, que identificou os cromossomos como as estruturas celulares responsáveis pela hereditariedade. Por suas muitas contribuições significativas à genética, Morgan recebeu o Prêmio Nobel de Fisiologia ou Medicina em 1933.
No final da década de 1920, Barbara McClintock (1902—1992) desenvolveu técnicas de coloração cromossômica para visualizar e diferenciar os diferentes cromossomos do milho (milho). Nas décadas de 1940 e 1950, ela identificou um evento de quebra no cromossomo 9, que ela chamou de locus de dissociação (Ds). Ds podem mudar de posição dentro do cromossomo. Ela também identificou um locus ativador (Ac). A quebra do cromossomo Ds pode ser ativada por um elemento Ac (enzima transposase). No início, a descoberta de McClintock desses genes saltadores, que agora chamamos de transposons, não foi aceita pela comunidade científica. Foi só na década de 1960 e mais tarde que os transposons foram descobertos em bacteriófagos, bactérias e drosófilas. Hoje, sabemos que os transposons são segmentos móveis de DNA que podem se mover dentro do genoma de um organismo. Eles podem regular a expressão gênica, a expressão de proteínas e a virulência (capacidade de causar doenças).
Micróbios e vírus na pesquisa genética
Os microbiologistas também desempenharam um papel crucial em nossa compreensão da genética. Organismos experimentais, como ervilhas de jardim de Mendel, moscas-das-frutas de Morgan e milho de McClintock, já haviam sido usados com sucesso para preparar o caminho para uma compreensão da genética. No entanto, micróbios e vírus foram (e ainda são) excelentes sistemas modelo para o estudo da genética porque, diferentemente da ervilha, da mosca-da-fruta e do milho, eles se propagam mais facilmente em laboratório, crescendo até altas densidades populacionais em uma pequena quantidade de espaço e em pouco tempo. Além disso, devido à sua simplicidade estrutural, micróbios e vírus são mais facilmente manipulados geneticamente.
Felizmente, apesar das diferenças significativas em tamanho, estrutura, estratégias de reprodução e outras características biológicas, há unidade bioquímica entre todos os organismos; eles têm em comum as mesmas moléculas subjacentes responsáveis pela hereditariedade e pelo uso de material genético para dar às células sua variação características. Nas palavras do cientista francês Jacques Monod, “O que é verdade para a E. coli também é verdade para o elefante”, o que significa que a bioquímica da vida foi mantida ao longo da evolução e é compartilhada em todas as formas de vida, desde organismos unicelulares simples até organismos grandes e complexos. Essa continuidade bioquímica torna os micróbios excelentes modelos para uso em estudos genéticos.
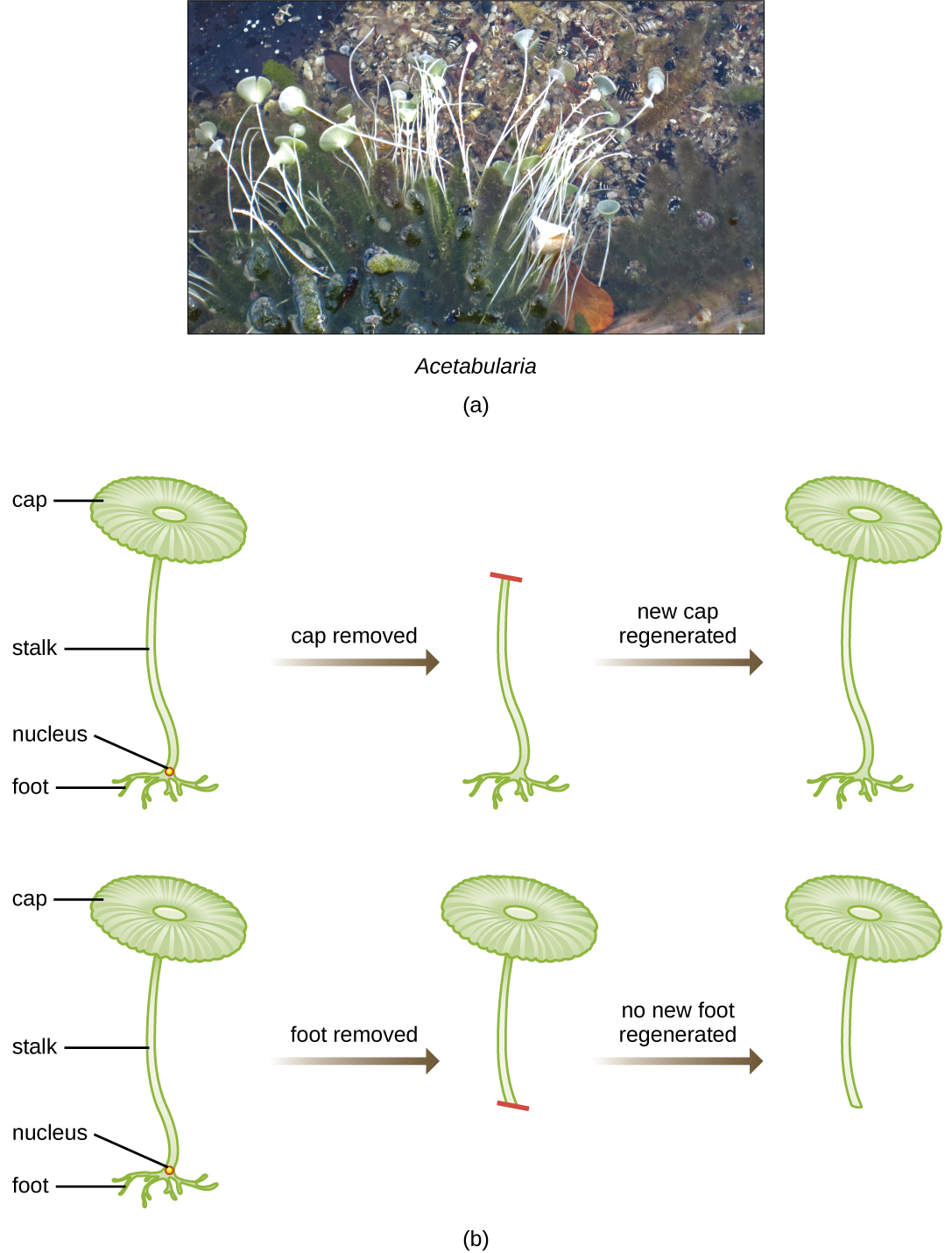
Em um conjunto inteligente de experimentos nas décadas de 1930 e 1940, o cientista alemão Joachim Hämmerling (1901—1980), usando a alga unicelular Acetabularia como modelo microbiano, estabeleceu que a informação genética em uma célula eucariótica está alojada dentro do núcleo. Acetabularia spp. são células algais invulgarmente grandes que crescem assimetricamente, formando um “pé” contendo o núcleo, que é usado para fixação do substrato; um caule; e uma tampa semelhante a um guarda-chuva - estruturas que podem ser facilmente vistas a olho nu. Em um conjunto inicial de experimentos, Hämmerling removeu a tampa ou o pé das células e observou se novas tampas ou pés estavam regenerados (Figura\(\PageIndex{2}\)). Ele descobriu que quando o pé dessas células foi removido, novos pés não cresceram; no entanto, quando as tampas foram removidas das células, novas capas foram regeneradas. Isso sugeriu que a informação hereditária estava localizada no pé contendo o núcleo de cada célula.
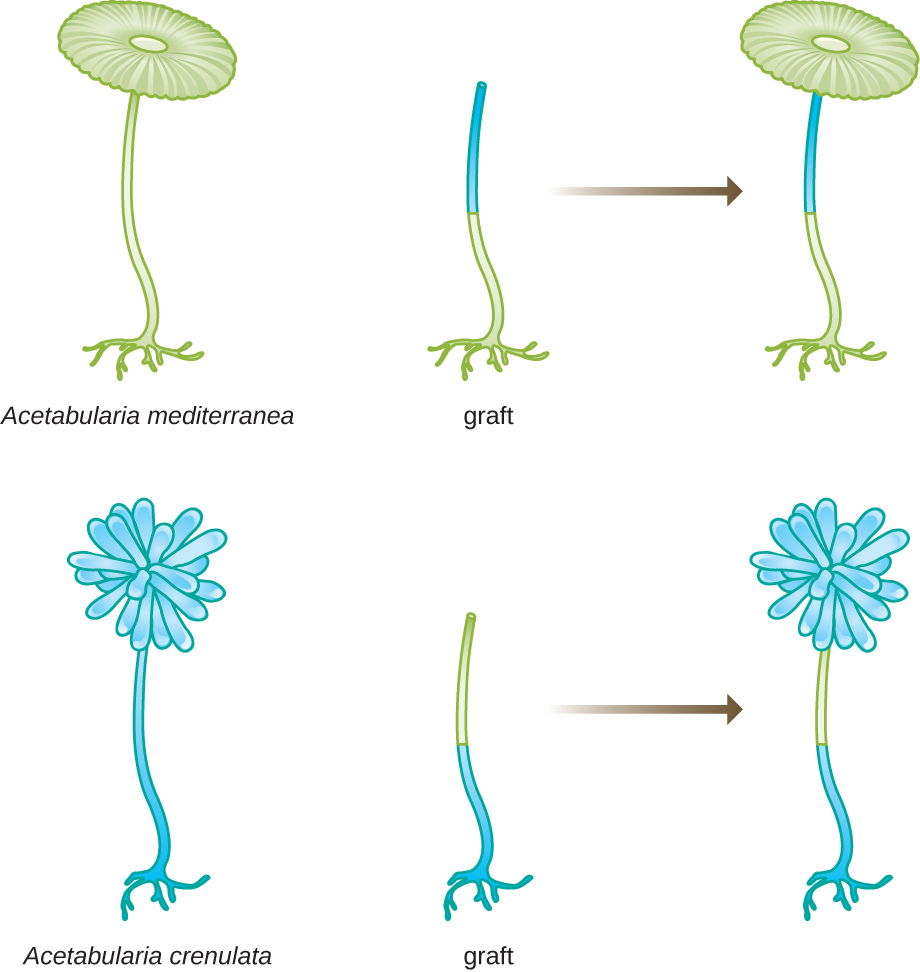
Em outro conjunto de experimentos, Hämmerling usou duas espécies de Acetabularia que têm diferentes morfologias de capa, A. crenulata e A. mediterranea (Figura\(\PageIndex{3}\)). Ele cortou as tampas de ambos os tipos de células e depois enxertou o caule de uma A. crenulata em um pé de A. mediterranea e vice-versa. Com o tempo, ele observou que a célula enxertada com o pé de A. crenulata e o caule de A. mediterranea desenvolveu uma capa com a morfologia de A. crenulata. Por outro lado, a célula enxertada com o pé de A. mediterranea e o caule de A. crenulata desenvolveu uma capa com a morfologia de A. mediterranea. Ele confirmou microscopicamente a presença de núcleos nos pés dessas células e atribuiu o desenvolvimento dessas morfologias do capô ao núcleo de cada célula enxertada. Assim, ele mostrou experimentalmente que o núcleo era a localização do material genético que ditava as propriedades de uma célula.
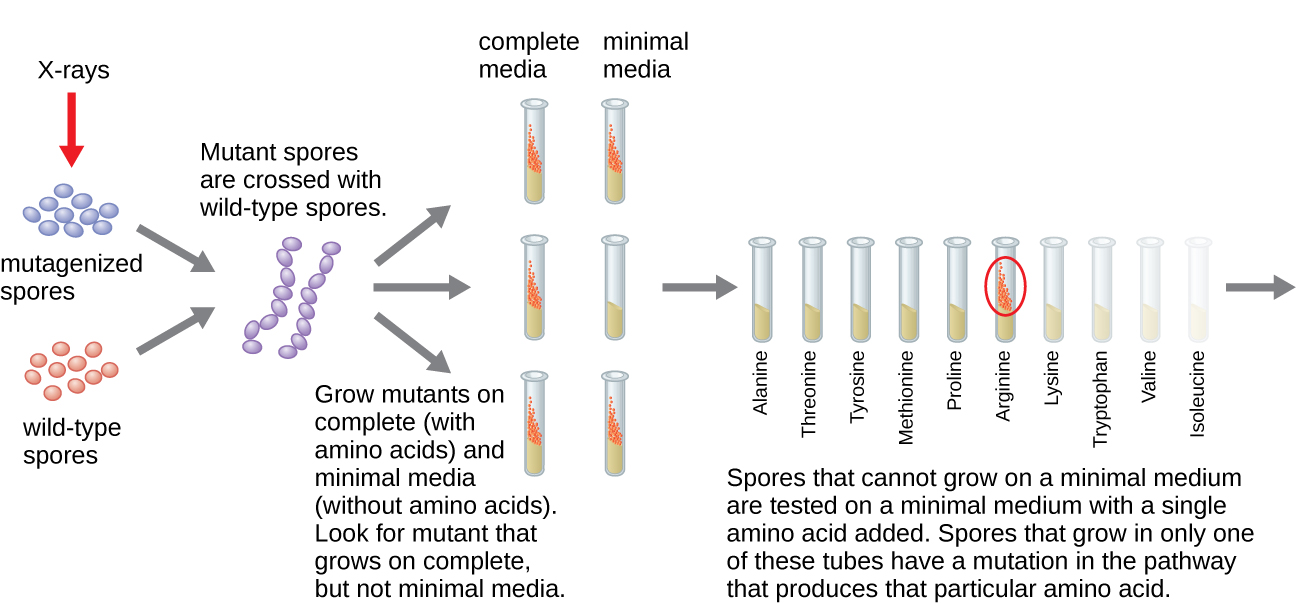
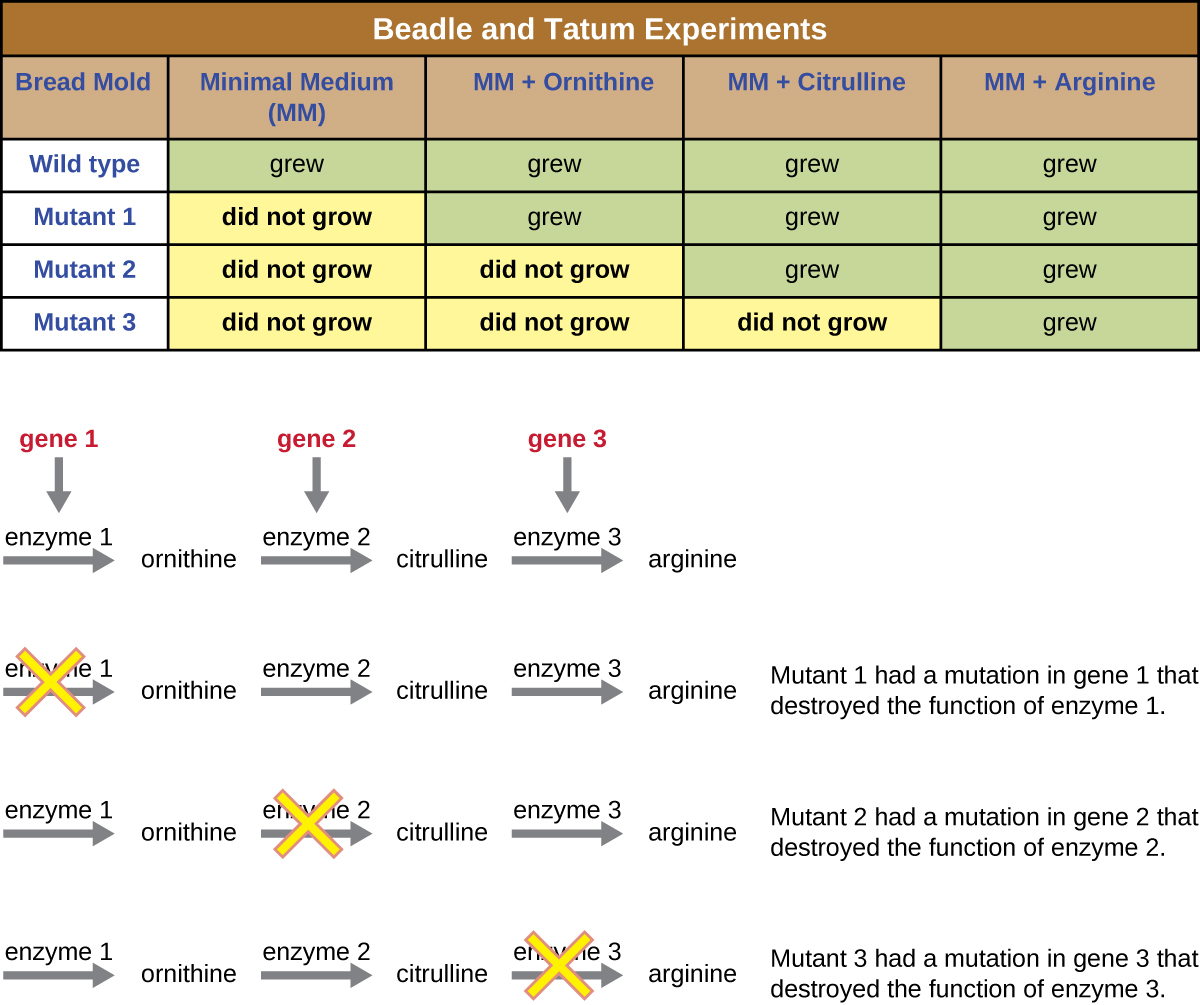
Outro modelo microbiano, o molde de pão vermelho Neurospora crassa, foi usado por George Beadle e Edward Tatum para demonstrar a relação entre os genes e as proteínas que eles codificam. Beadle havia trabalhado com moscas-das-frutas no laboratório de Morgan, mas as achou muito complexas para realizar certos tipos de experimentos. A N. crassa, por outro lado, é um organismo mais simples e tem a capacidade de crescer em um meio mínimo porque contém vias enzimáticas que lhe permitem usar o meio para produzir suas próprias vitaminas e aminoácidos.
Beadle e Tatum irradiaram o molde com raios-X para induzir mudanças em uma sequência de ácidos nucléicos, chamadas mutações. Eles acasalaram os esporos de fungos irradiados e tentaram cultivá-los em um meio completo e em um meio mínimo. Eles procuraram mutantes que crescessem em um meio completo, suplementados com vitaminas e aminoácidos, mas não cresceram no meio mínimo sem esses suplementos. Esses moldes teoricamente continham mutações nos genes que codificavam as vias biossintéticas. Ao encontrar esses mutantes, eles testaram sistematicamente cada um para determinar qual vitamina ou aminoácido não era capaz de produzir (Figura\(\PageIndex{4}\)) e publicaram esse trabalho em 1941.
Trabalhos subsequentes de Beadle, Tatum e colegas mostraram que eles poderiam isolar diferentes classes de mutantes que exigiam um suplemento específico, como o aminoácido arginina (Figura\(\PageIndex{5}\)). Com algum conhecimento da via de biossíntese da arginina, eles identificaram três classes de mutantes da arginina suplementando o meio mínimo com intermediários (citrulina ou ornitina) na via. Os três mutantes diferiram em suas habilidades de crescer em cada uma das mídias, o que levou o grupo de cientistas a propor, em 1945, que cada tipo de mutante tinha um defeito em um gene diferente na via de biossíntese da arginina. Isso levou à chamada hipótese de um gene - uma enzima, que sugeria que cada gene codifica uma enzima.
O conhecimento subsequente sobre os processos de transcrição e tradução levou os cientistas a revisá-lo para a hipótese “um gene - um polipeptídeo”. Embora existam alguns genes que não codificam polipeptídeos (mas codificam para RNAs de transferência [tRNAs] ou RNAs ribossômicos [rRNAs], que discutiremos mais adiante), a hipótese de um gene - uma enzima é verdadeira em muitos casos, especialmente em micróbios. A descoberta da ligação entre genes e características correspondentes por Beadle e Tatum rendeu-lhes o Prêmio Nobel de Fisiologia e Medicina de 1958 e, desde então, tornou-se a base da genética molecular moderna.
Para saber mais sobre os experimentos de Beadle e Tatum, visite este site do DNA Learning Center.
Exercício\(\PageIndex{2}\)
- Que organismo Morgan e seus colegas usaram para desenvolver a Teoria Cromossômica da Herança? Quais características eles rastrearam?
- O que Hämmerling provou com seus experimentos com acetabularia?
DNA como molécula responsável pela hereditariedade
No início do século XX, muito trabalho já havia sido feito na caracterização do DNA e no estabelecimento dos fundamentos da genética, incluindo a atribuição de hereditariedade aos cromossomos encontrados no núcleo. Apesar de todas essas pesquisas, foi somente no século 20 que essas linhas de pesquisa convergiram e os cientistas começaram a considerar que o DNA poderia ser o material genético que os filhos herdaram de seus pais. Acreditava-se que o DNA, contendo apenas quatro nucleotídeos diferentes, fosse estruturalmente muito simples para codificar informações genéticas tão complexas. Em vez disso, acreditava-se que a proteína tivesse a complexidade necessária para servir como informação genética celular porque é composta por 20 aminoácidos diferentes que podem ser combinados em uma grande variedade de combinações. Os microbiologistas desempenharam um papel fundamental na pesquisa que determinou que o DNA é a molécula responsável pela hereditariedade.
Experimentos de transformação de Griffith
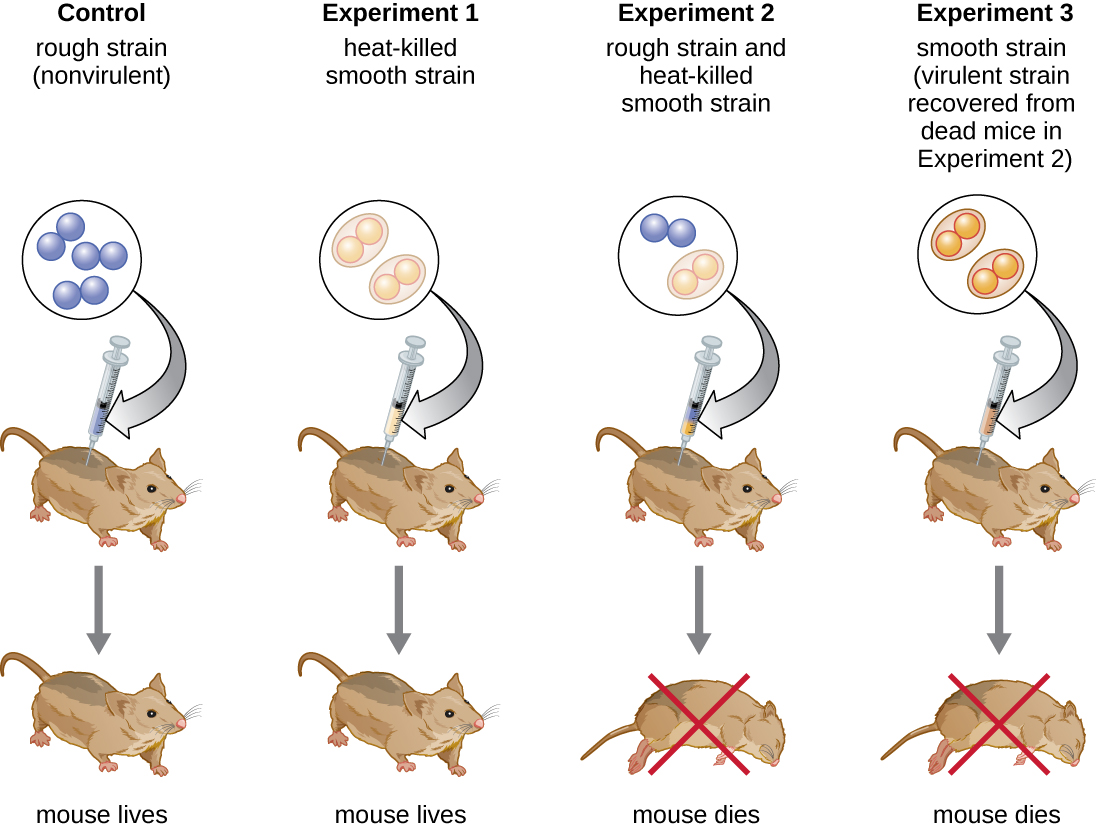
O bacteriologista britânico Frederick Griffith (1879-1941) foi talvez a primeira pessoa a mostrar que as informações hereditárias poderiam ser transferidas de uma célula para outra “horizontalmente” (entre membros da mesma geração), em vez de “verticalmente” (de pais para filhos). Em 1928, ele relatou a primeira demonstração de transformação bacteriana, um processo no qual o DNA externo é captado por uma célula, alterando assim suas características. 3 Ele estava trabalhando com duas cepas de Streptococcus pneumoniae, uma bactéria que causa pneumonia: uma cepa áspera (R) e uma cepa lisa (S). A cepa R não é patogênica e não possui uma cápsula em sua superfície externa; como resultado, as colônias da cepa R parecem ásperas quando cultivadas em placas. A cepa S é patogênica e tem uma cápsula fora de sua parede celular, permitindo que ela escape da fagocitose pelo sistema imunológico do hospedeiro. As cápsulas fazem com que as colônias da cepa S pareçam lisas quando cultivadas em placas.
Em uma série de experimentos, Griffith analisou os efeitos de cepas R vivas, S vivas e S mortas pelo calor de S. pneumoniae em camundongos vivos (Figura\(\PageIndex{6}\)). Quando os camundongos foram injetados com a cepa S viva, os camundongos morreram. Quando ele injetou nos camundongos a cepa R viva ou a cepa S, morta pelo calor, os camundongos sobreviveram. Mas quando ele injetou nos camundongos uma mistura de cepa R viva e cepa S morta pelo calor, os camundongos morreram. Ao isolar a bactéria viva do camundongo morto, ele recuperou apenas a cepa S da bactéria. Quando ele então injetou essa cepa S isolada em camundongos frescos, os camundongos morreram. Griffith concluiu que algo havia passado da cepa S morta pelo calor para a cepa R viva e a “transformou” na cepa S patogênica; ele chamou isso de “princípio transformador”. Esses experimentos agora são conhecidos como experimentos de transformação de Griffith.
Em 1944, Oswald Avery, Colin MacLeod e Maclyn McCarty estavam interessados em explorar ainda mais o princípio transformador de Griffith. Eles isolaram a cepa S de camundongos mortos infectados, a mataram termicamente e inativaram vários componentes do extrato S, conduzindo um estudo sistemático de eliminação (Figura\(\PageIndex{7}\)). Eles usaram enzimas que degradaram especificamente proteínas, RNA e DNA e misturaram o extrato de S com cada uma dessas enzimas individuais. Em seguida, eles testaram a capacidade resultante de cada combinação de extrato/enzima de transformar a cepa R, conforme observado pelo crescimento difuso da cepa S em meio de cultura e confirmado visualmente pelo crescimento em placas. Eles descobriram que quando o DNA era degradado, a mistura resultante não era mais capaz de transformar a bactéria da cepa R, enquanto nenhum outro tratamento enzimático foi capaz de impedir a transformação. Isso os levou a concluir que o DNA era o princípio transformador. Apesar dos resultados, muitos cientistas não aceitaram sua conclusão, acreditando que havia contaminantes proteicos em seus extratos.
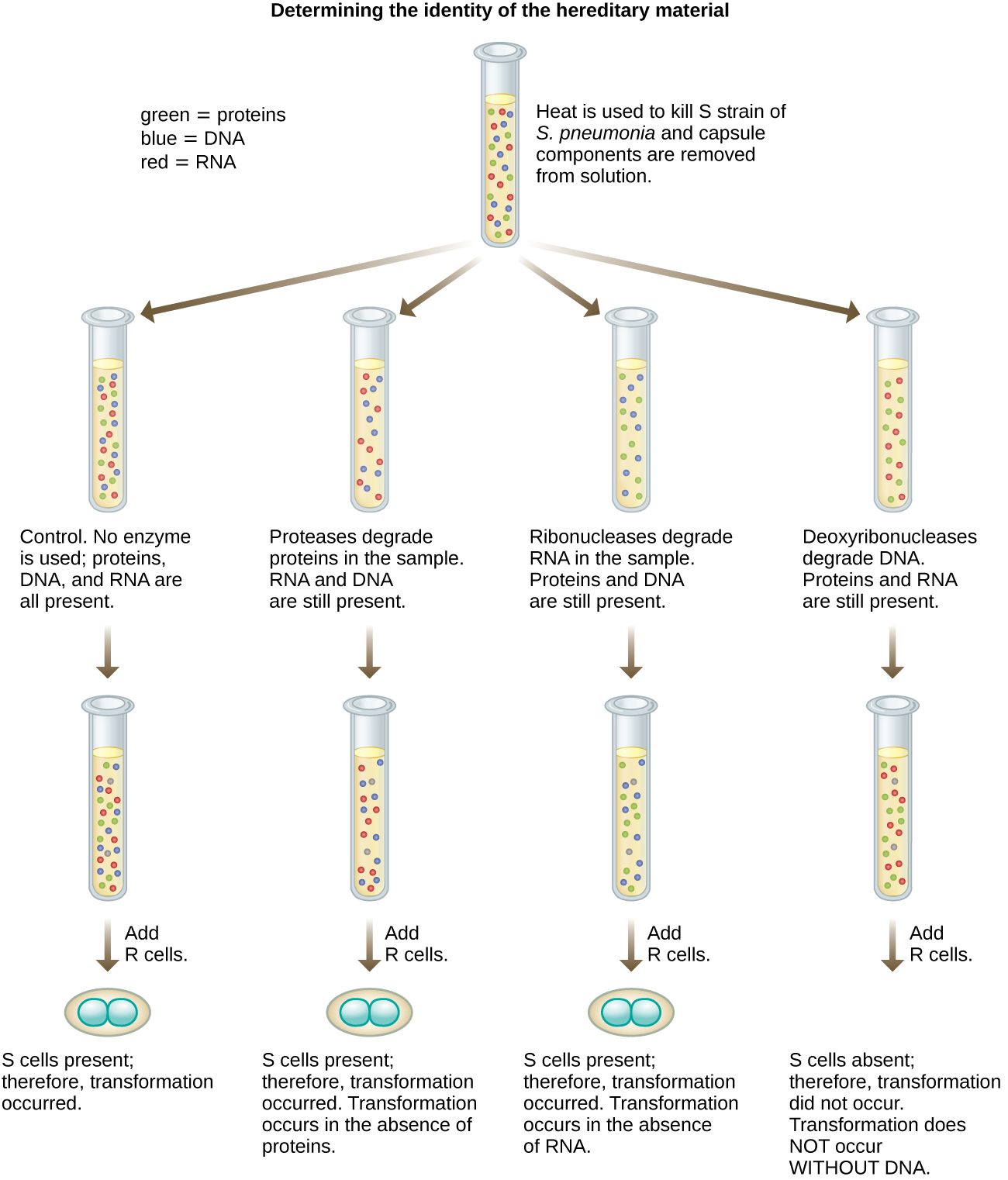
Exercício\(\PageIndex{3}\)
Como os experimentos de Avery, MacLeod e McCarty mostraram que o DNA foi o princípio transformador descrito pela primeira vez por Griffith?
Prova de DNA como material genético de Hershey e Chase
Alfred Hershey e Martha Chase realizaram seus próprios experimentos em 1952 e foram capazes de fornecer evidências confirmatórias de que o DNA, não a proteína, era o material genético (Figura\(\PageIndex{8}\)). 4 Hershey e Chase estavam estudando um bacteriófago, um vírus que infecta bactérias. Os vírus geralmente têm uma estrutura simples: um revestimento proteico, chamado capsídeo, e um núcleo de ácido nucléico que contém o material genético, seja DNA ou RNA (veja Vírus). O bacteriófago específico que eles estavam estudando era o bacteriófago T2, que infecta as células de E. coli. Como sabemos hoje, o T2 se liga à superfície da célula bacteriana e, em seguida, injeta seus ácidos nucléicos dentro da célula. O DNA do fago faz várias cópias de si mesmo usando a maquinaria hospedeira e, eventualmente, a célula hospedeira se rompe, liberando um grande número de bacteriófagos.
Hershey e Chase rotularam o revestimento proteico em um lote de fago usando enxofre radioativo, 35 S, porque o enxofre é encontrado nos aminoácidos metionina e cisteína, mas não nos ácidos nucléicos. Eles rotularam o DNA em outro lote usando fósforo radioativo, 32 P, porque o fósforo é encontrado no DNA e no RNA, mas não normalmente nas proteínas.
Cada lote de fago foi autorizado a infectar as células separadamente. Após a infecção, Hershey e Chase colocaram cada suspensão bacteriana do fago em um liquidificador, que separou as camadas do fago da célula hospedeira e girou a suspensão resultante em uma centrífuga. As células bacterianas mais pesadas se estabeleceram e formaram um pellet, enquanto as partículas mais leves do fago permaneceram no sobrenadante. No tubo com a proteína marcada, a radioatividade permaneceu apenas no sobrenadante. No tubo com o DNA marcado, a radioatividade foi detectada apenas nas células bacterianas. Hershey e Chase concluíram que foi o DNA do fago injetado na célula que transportou as informações para produzir mais partículas de fago, provando assim que o DNA, não as proteínas, era a fonte do material genético. Como resultado de seu trabalho, a comunidade científica aceitou mais amplamente o DNA como a molécula responsável pela hereditariedade.
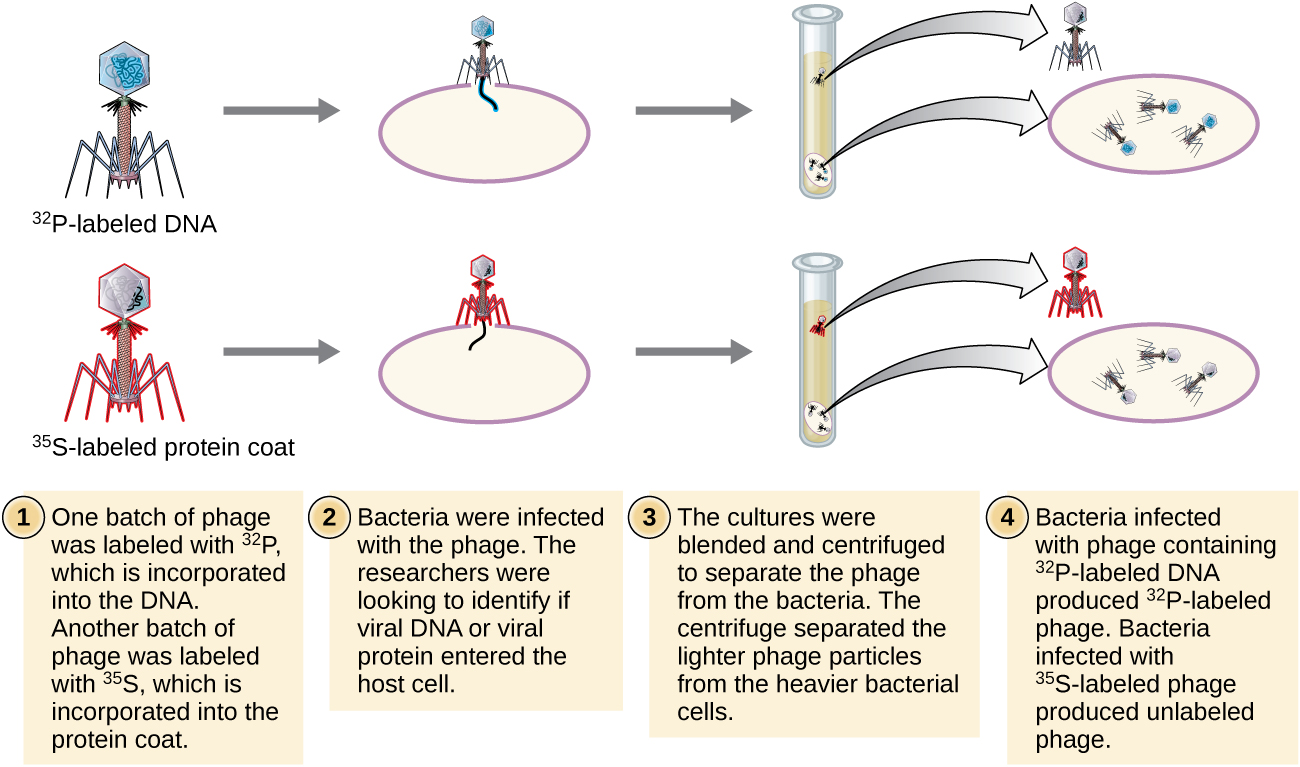
Quando Hershey e Chase publicaram seu experimento no início dos anos 1950, microbiologistas e outros cientistas estavam pesquisando a hereditariedade há mais de 80 anos. Com base nas pesquisas uns dos outros durante esse período, culminou no acordo geral de que o DNA era o material genético responsável pela hereditariedade (Figura\(\PageIndex{9}\)). Esse conhecimento preparou o terreno para a próxima era da biologia molecular e os avanços significativos em biotecnologia e biologia de sistemas que estamos experimentando hoje.
Para saber mais sobre os experimentos envolvidos na história da genética e a descoberta do DNA como material genético das células, visite este site do Centro de Aprendizagem de DNA.
Exercício\(\PageIndex{4}\)
Como Hershey e Chase usaram micróbios para provar que o DNA é material genético?
Conceitos principais e resumo
- O DNA foi descoberto e caracterizado muito antes de seu papel na hereditariedade ser compreendido. Os microbiologistas desempenharam um papel significativo na demonstração de que o DNA é a informação hereditária encontrada nas células.
- Nas décadas de 1850 e 1860, Gregor Mendel experimentou ervilhas de jardim verdadeiras para demonstrar a herdabilidade de características observáveis específicas.
- Em 1869, Friedrich Miescher isolou e purificou um composto rico em fósforo dos núcleos dos glóbulos brancos; ele chamou o composto de nucleína. O aluno de Miescher, Richard Altmann, descobriu sua natureza ácida, renomeando-a como ácido nucléico. Albrecht Kossell caracterizou as bases nucleotídicas encontradas nos ácidos nucléicos.
- Embora Walter Sutton e Theodor Boveri tenham proposto a Teoria Cromossômica da Herança em 1902, ela não foi demonstrada cientificamente até a publicação em 1915 do trabalho de Thomas Hunt Morgan e seus colegas.
- Usando Acetabularia, uma grande célula de algas, como seu sistema modelo, Joachim Hämmerling demonstrou nas décadas de 1930 e 1940 que o núcleo era a localização da informação hereditária nessas células.
- Na década de 1940, George Beadle e Edward Tatum usaram o molde Neurospora crassa para mostrar que a produção de cada proteína estava sob o controle de um único gene, demonstrando a hipótese “um gene — uma enzima”.
- Em 1928, Frederick Griffith mostrou que bactérias mortas encapsuladas podiam passar informações genéticas para bactérias vivas não encapsuladas e transformá-las em cepas nocivas. Em 1944, Oswald Avery, Colin McLeod e Maclyn McCarty identificaram o composto como DNA.
- A natureza do DNA como molécula que armazena informações genéticas foi demonstrada inequivocamente no experimento de Alfred Hershey e Martha Chase publicado em 1952. O DNA marcado de vírus bacterianos entrou e infectou as células bacterianas, dando origem a mais partículas virais. Os revestimentos proteicos marcados não participaram da transmissão da informação genética.
Notas de pé
- 1 J. G. Mendel. “Versuche über Pflanzenhybriden.” Verhandlungen des naturforschenden Vereines em Brünn, Bd. Abhandlungen 4 (1865) :3—7. (Para tradução em inglês, consulte http://www.mendelweb.org/Mendel.plain.html)
- 2 G.W. Beadle, E. L. Tatum. “Controle genético de reações bioquímicas em Neurospora.” Anais da Academia Nacional de Ciências 27 no. 11 (1941) :499—506.
- 3 F. Griffith. “A importância dos tipos pneumocócicas”. Journal of Hygiene 27 no. 2 (1928) :8—159.
- 4 A.D. Hershey, Sr. Chase. “Funções independentes da proteína viral e do ácido nucléico no crescimento do bacteriófago.” Jornal de Fisiologia Geral 36 no. 1 (1952) :39—56.