
8.6: Fotossíntese e a importância da luz
- Page ID
- 181458

Objetivos de
- Descreva a função e a localização dos pigmentos fotossintéticos em eucariotos e procariontes
- Descreva os principais produtos das reações dependentes da luz e independentes da luz
- Descreva as reações que produzem glicose em uma célula fotossintética
- Compare e contraste a fotofosforilação cíclica e não cíclica
Organismos heterotróficos, desde E. coli até humanos, dependem da energia química encontrada principalmente nas moléculas de carboidratos. Muitos desses carboidratos são produzidos pela fotossíntese, o processo bioquímico pelo qual organismos fototróficos convertem energia solar (luz solar) em energia química. Embora a fotossíntese seja mais comumente associada às plantas, a fotossíntese microbiana também é um fornecedor significativo de energia química, alimentando diversos ecossistemas. Nesta seção, vamos nos concentrar na fotossíntese microbiana.
A fotossíntese ocorre em dois estágios sequenciais: as reações dependentes da luz e as reações independentes da luz (Figura\(\PageIndex{1}\)). Nas reações dependentes da luz, a energia da luz solar é absorvida pelas moléculas de pigmento nas membranas fotossintéticas e convertida em energia química armazenada. Nas reações independentes da luz, a energia química produzida pelas reações dependentes da luz é usada para conduzir a montagem de moléculas de açúcar usando CO 2; no entanto, essas reações ainda dependem da luz porque os produtos das reações dependentes da luz necessárias para conduzi-las são curtos -viveu. As reações dependentes da luz produzem ATP e NADPH ou NADH para armazenar energia temporariamente. Esses transportadores de energia são usados nas reações independentes da luz para impulsionar o processo energeticamente desfavorável de “fixar” o CO 2 inorgânico em uma forma orgânica, o açúcar.
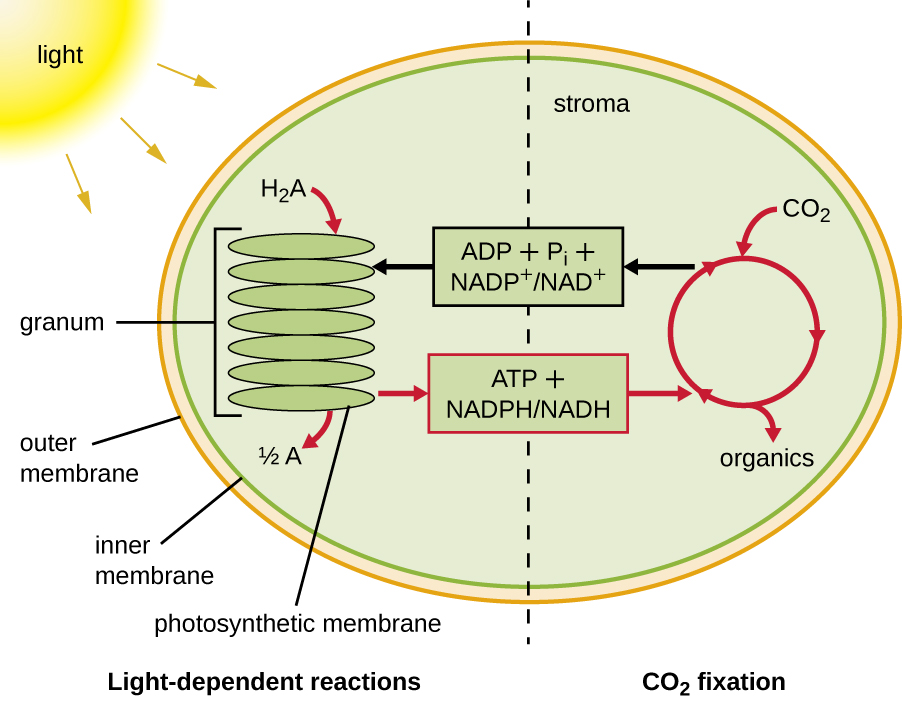
Estruturas fotossintéticas em eucariotos e procariontes
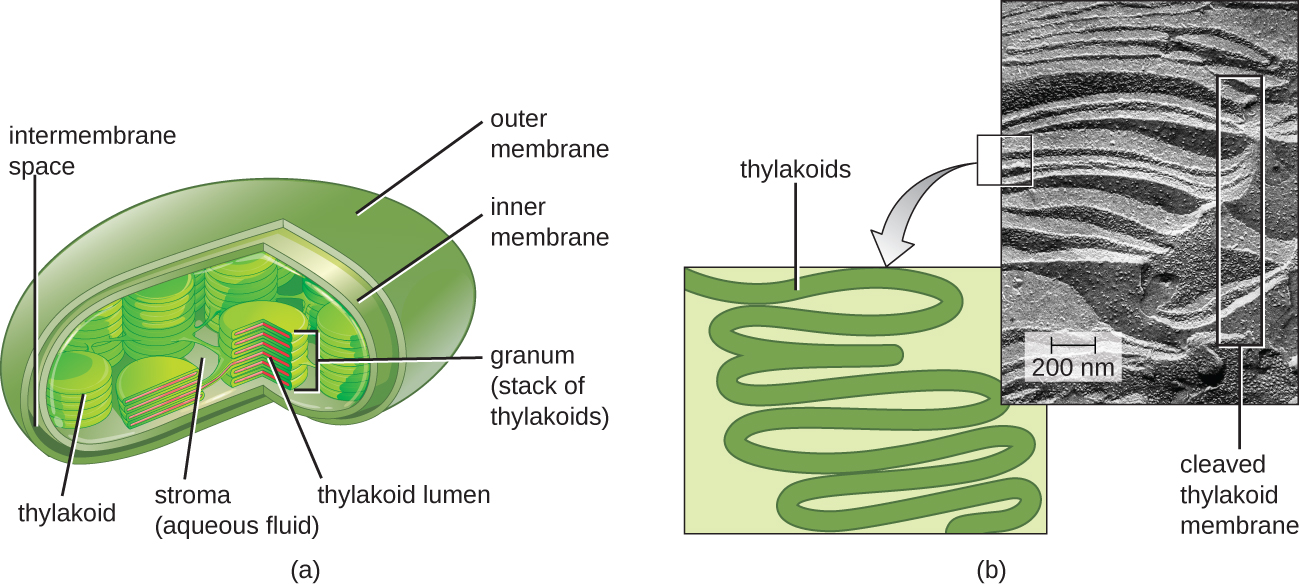
Em todos os eucariotos fototróficos, a fotossíntese ocorre dentro de um cloroplasto, uma organela que surgiu nos eucariotos pela endossimbiose de uma bactéria fotossintética (veja Características únicas das células eucarióticas). Esses cloroplastos são cercados por uma membrana dupla com camadas interna e externa. Dentro do cloroplasto há uma terceira membrana que forma estruturas fotossintéticas empilhadas em forma de disco chamadas tilacóides (Figura\(\PageIndex{2}\)). Uma pilha de tilacóides é chamada de grão, e o espaço ao redor do grão dentro do cloroplasto é chamado de estroma.
As membranas fotossintéticas em procariontes, por outro lado, não estão organizadas em organelas distintas fechadas por membrana; em vez disso, são regiões desdobradas da membrana plasmática. Nas cianobactérias, por exemplo, essas regiões desdobradas também são chamadas de tilacóides. Em ambos os casos, embutidas nas membranas tilacóides ou em outras membranas bacterianas fotossintéticas estão moléculas de pigmento fotossintético organizadas em um ou mais fotossistemas, onde a energia da luz é realmente convertida em energia química.
Os pigmentos fotossintéticos dentro das membranas fotossintéticas são organizados em fotossistemas, cada um dos quais é composto por um complexo de captação de luz (antenas) e um centro de reação. O complexo de coleta de luz consiste em várias proteínas e pigmentos associados, cada um deles pode absorver a energia da luz e, assim, ficar excitado. Essa energia é transferida de uma molécula de pigmento para outra até que, eventualmente (após cerca de um milionésimo de segundo), é entregue ao centro de reação. Até esse ponto, somente energia — não elétrons — foi transferida entre as moléculas. O centro de reação contém uma molécula de pigmento que pode sofrer oxidação após a excitação, liberando na verdade um elétron. É nessa etapa da fotossíntese que a energia da luz é convertida em um elétron excitado.
Diferentes tipos de pigmentos que captam luz absorvem padrões únicos de comprimentos de onda (cores) da luz visível. Os pigmentos refletem ou transmitem os comprimentos de onda que não conseguem absorver, fazendo com que pareçam com a cor correspondente. Exemplos de pigmentos fotossintéticos (moléculas usadas para absorver energia solar) são bacterioclorofilas (verdes, roxas ou vermelhas), carotenóides (laranja, vermelho ou amarelo), clorofilas (verdes), ficocianinas (azuis) e ficoeritrinas (vermelhas). Por ter misturas de pigmentos, um organismo pode absorver energia de mais comprimentos de onda. Como as bactérias fotossintéticas geralmente crescem em competição pela luz solar, cada tipo de bactéria fotossintética é otimizada para capturar os comprimentos de onda da luz aos quais é comumente exposta, levando à estratificação das comunidades microbianas nos ecossistemas aquáticos e do solo pela qualidade e penetração da luz.
Uma vez que o complexo de captação de luz transfere a energia para o centro de reação, o centro de reação entrega seus elétrons de alta energia, um por um, para um transportador de elétrons em um sistema de transporte de elétrons, e a transferência de elétrons através do ETS é iniciada. O ETS é semelhante ao usado na respiração celular e está embutido na membrana fotossintética. Em última análise, o elétron é usado para produzir NADH ou NADPH. O gradiente eletroquímico que se forma através da membrana fotossintética é usado para gerar ATP por quimiosmose por meio do processo de fotofosforilação, outro exemplo de fosforilação oxidativa (Figura\(\PageIndex{3}\)).
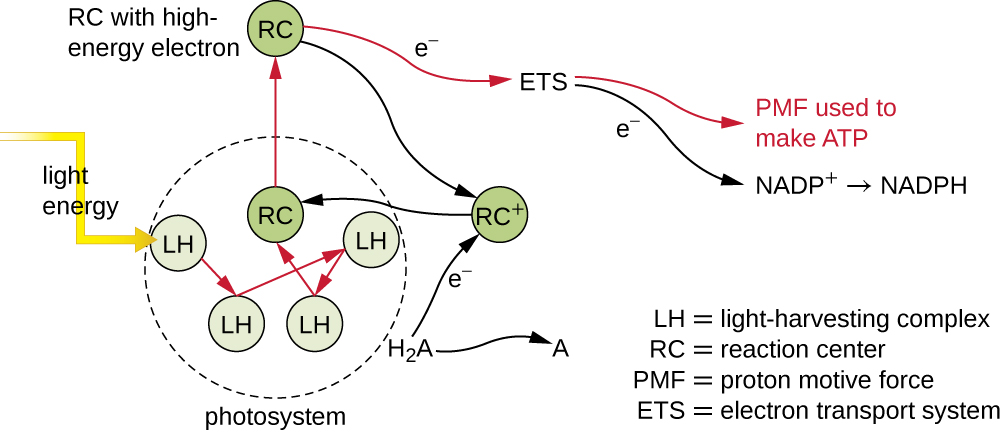
Exercício\(\PageIndex{1}\)
Em um eucarioto fototrófico, onde ocorre a fotossíntese?
Fotossíntese oxigênica e anoxigênica
Para que a fotossíntese continue, o elétron perdido do pigmento do centro de reação deve ser substituído. A fonte desse elétron (H 2 A) diferencia a fotossíntese oxigênica de plantas e cianobactérias da fotossíntese anoxigênica realizada por outros tipos de fototróficos bacterianos (Figura\(\PageIndex{4}\)). Na fotossíntese oxigenada, o H 2 O é dividido e fornece o elétron ao centro da reação. Como o oxigênio é gerado como um subproduto e é liberado, esse tipo de fotossíntese é conhecido como fotossíntese oxigênica. No entanto, quando outros compostos reduzidos servem como doadores de elétrons, o oxigênio não é gerado; esses tipos de fotossíntese são chamados de fotossíntese anoxigênica. O sulfeto de hidrogênio (H 2 S) ou tiossulfato\(\ce{(S2O3^{2-})}\) pode servir como doador de elétrons, gerando como resultado\(\ce{(SO4^{2-})}\) íons elementares de enxofre e sulfato, respectivamente.
Os fotossistemas foram classificados em dois tipos: fotossistema I (PSI) e fotossistema II (PSII) (Figura\(\PageIndex{5}\)). As cianobactérias e os cloroplastos vegetais têm ambos os fotossistemas, enquanto as bactérias fotossintéticas anoxigênicas usam apenas um dos fotossistemas. Ambos os fotossistemas são excitados pela energia da luz simultaneamente. Se a célula precisar de ATP e NADPH para a biossíntese, ela realizará a fotofosforilação não cíclica. Ao passar do elétron do centro de reação do PSII para o ETS que conecta PSII e PSI, o elétron perdido do centro de reação do PSII é substituído pela divisão da água. O elétron excitado do centro de reação PSI é usado para reduzir NADP + a NADPH e é substituído pelo elétron que sai do ETS. O fluxo de elétrons dessa maneira é chamado de esquema Z.
Se a necessidade de ATP de uma célula for significativamente maior do que a necessidade de NADPH, ela pode ignorar a produção de energia redutora por meio da fotofosforilação cíclica. Somente PSI é usado durante a fotofosforilação cíclica; o elétron de alta energia do centro de reação PSI é passado para um transportador de ETS e, em seguida, retorna ao pigmento oxidado do centro de reação PSI, reduzindo-o assim.
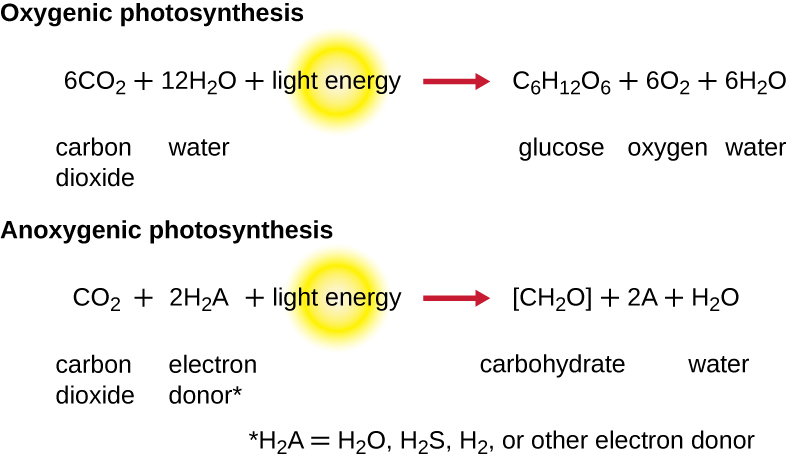
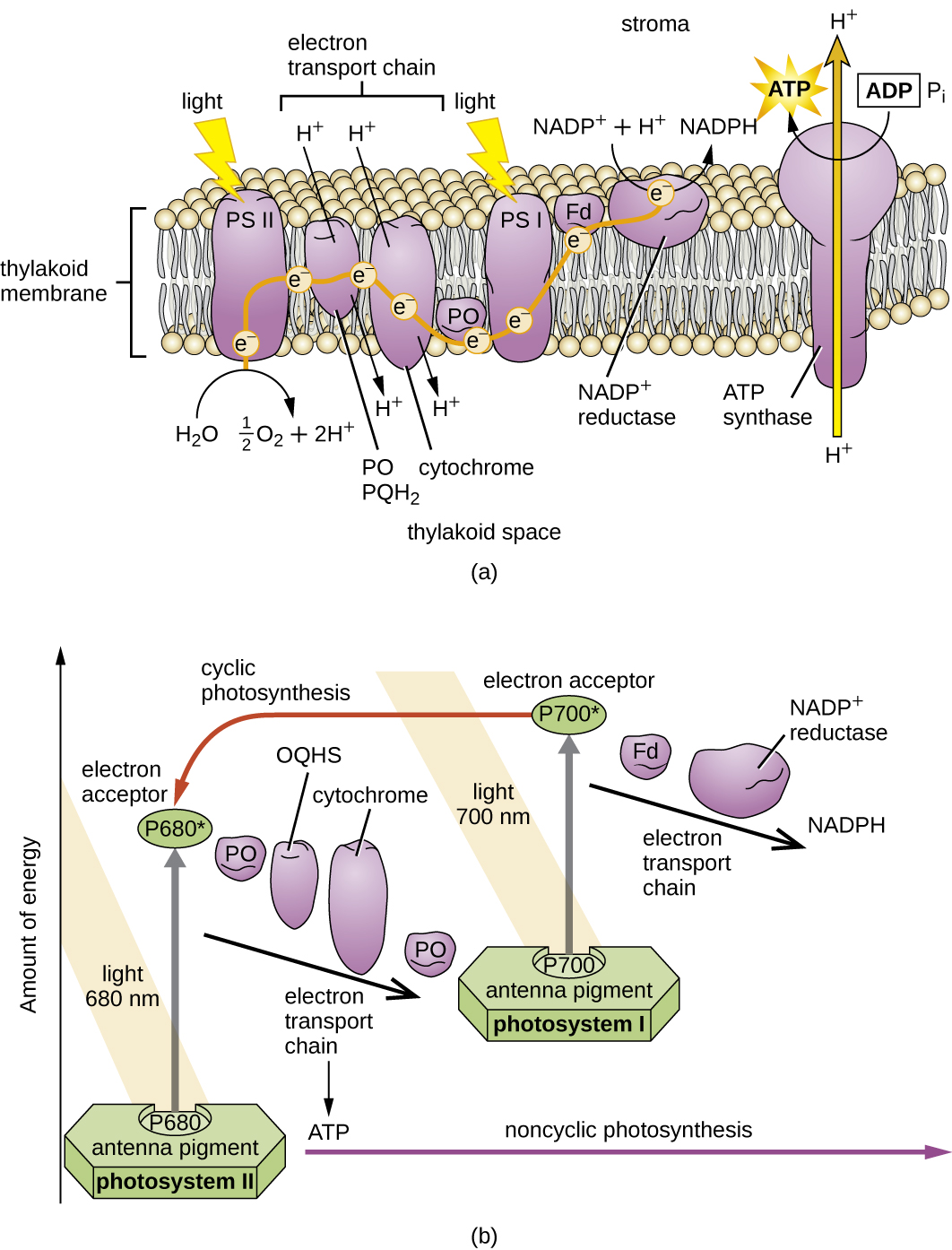
Exercício\(\PageIndex{2}\)
Por que uma bactéria fotossintética teria pigmentos diferentes?
Reações independentes da luz
Depois que a energia do sol é convertida em energia química e temporariamente armazenada nas moléculas de ATP e NADPH (com vida útil de milionésimos de segundo), os fotoautotróficos têm o combustível necessário para construir moléculas de carboidratos multicarbonetos, que podem sobreviver por centenas de milhões de anos, por longo prazo armazenamento de energia. O carbono vem do CO 2, o gás que é um produto residual da respiração celular.
O ciclo Calvin-Benson (nomeado em homenagem a Melvin Calvin [1911—1997] e Andrew Benson [1917—2015]), a via bioquímica usada para fixação de CO 2, está localizado no citoplasma de bactérias fotossintéticas e no estroma dos cloroplastos eucarióticos. As reações independentes da luz do ciclo de Calvin podem ser organizadas em três estágios básicos: fixação, redução e regeneração (consulte o Apêndice C para uma ilustração detalhada do ciclo de Calvin).
- Fixação: A enzima ribulose bisfosfato carboxilase (RuBisCO) catalisa a adição de CO 2 ao bisfosfato de ribulose (RuBP). Isso resulta na produção de 3-fosfoglicerato (3-PGA).
- Redução: Seis moléculas de ATP e NADPH (das reações dependentes da luz) são usadas para converter 3-PGA em gliceraldeído 3-fosfato (G3P). Algum G3P é então usado para construir glicose.
- Regeneração: O G3P restante não usado para sintetizar glicose é usado para regenerar o RuBP, permitindo que o sistema continue a fixação de CO 2. Mais três moléculas de ATP são usadas nessas reações de regeneração.
O ciclo de Calvin é usado extensivamente por plantas e bactérias fotoautotróficas, e a enzima RuBisCo é considerada a enzima mais abundante na Terra, compondo 30% a 50% da proteína solúvel total nos cloroplastos vegetais. 1 No entanto, além de seu uso predominante em fotoautotróficos, o ciclo de Calvin também é usado por muitos quimioautotróficos não fotossintéticos para fixar CO 2. Além disso, outras bactérias e arquéias usam sistemas alternativos para fixação de CO 2. Embora a maioria das bactérias que usam alternativas do ciclo de Calvin sejam quimioautotróficas, também foi demonstrado que certas bactérias fotoautotróficas de enxofre verde usam uma via alternativa de fixação de CO 2.
Exercício\(\PageIndex{3}\)
Descreva os três estágios do ciclo de Calvin.
Conceitos principais e resumo
- Os heterotróficos dependem dos carboidratos produzidos pelos autótrofos, muitos dos quais são fotossintéticos, convertendo a energia solar em energia química.
- Diferentes organismos fotossintéticos usam diferentes misturas de pigmentos fotossintéticos, que aumentam a faixa dos comprimentos de onda da luz que um organismo pode absorver.
- Cada fotossistema (PSI e PSII) contém um complexo de captação de luz, composto por várias proteínas e pigmentos associados que absorvem a energia da luz. As reações fotossintéticas dependentes da luz convertem energia solar em energia química, produzindo ATP e NADPH ou NADH para armazenar temporariamente essa energia.
- Na fotossíntese oxigenada, o H 2 O serve como doador de elétrons para substituir o elétron do centro de reação e o oxigênio é formado como um subproduto. Na fotossíntese anoxigênica, outras moléculas reduzidas, como H 2 S ou tiossulfato, podem ser usadas como doadoras de elétrons; como tal, o oxigênio não é formado como um subproduto.
- A fotofosforilação não cíclica é usada na fotossíntese oxigênica quando há necessidade de produção de ATP e NADPH. Se as necessidades de ATP de uma célula superarem suas necessidades de NADPH, ela poderá realizar a fotofosforilação cíclica, produzindo apenas ATP.
- As reações de fotossíntese independentes da luz usam o ATP e o NADPH das reações dependentes da luz para fixar CO 2 em moléculas de açúcar orgânico.
Notas de pé
- 1 A. Dhingra et al. “A tradução aprimorada de um gene Rbc S expresso em cloroplasto restaura pequenos níveis de subunidades e fotossíntese em plantas nucleares antisense Rbc S”. Anais da Academia Nacional de Ciências dos Estados Unidos da América 101 no. 16 (2004) :6315—6320.