
30.5: Transporte de água e solutos em plantas
- Page ID
- 182019

Habilidades para desenvolver
- Defina o potencial hídrico e explique como ele é influenciado por solutos, pressão, gravidade e potencial matricial
- Descreva como o potencial hídrico, a evapotranspiração e a regulação estomática influenciam a forma como a água é transportada nas plantas
- Explique como os fotossintatos são transportados nas plantas
A estrutura das raízes, caules e folhas das plantas facilita o transporte de água, nutrientes e fotossintatos por toda a planta. O floema e o xilema são os principais tecidos responsáveis por esse movimento. O potencial hídrico, a evapotranspiração e a regulação estomática influenciam a forma como a água e os nutrientes são transportados nas plantas. Para entender como esses processos funcionam, devemos primeiro entender a energética do potencial hídrico.
Potencial de
As plantas são engenheiros hidráulicos fenomenais. Usando apenas as leis básicas da física e a simples manipulação da energia potencial, as plantas podem mover a água até o topo de uma árvore de 116 metros de altura (Figura\(\PageIndex{1}\) a). As plantas também podem usar a hidráulica para gerar força suficiente para quebrar pedras e fechar calçadas (Figura\(\PageIndex{1}\) b). As plantas conseguem isso devido ao potencial hídrico.

O potencial hídrico é uma medida da energia potencial na água. Os fisiologistas vegetais não estão interessados na energia de nenhum sistema aquoso em particular, mas estão muito interessados no movimento da água entre dois sistemas. Em termos práticos, portanto, o potencial hídrico é a diferença na energia potencial entre uma determinada amostra de água e a água pura (à pressão atmosférica e à temperatura ambiente). O potencial hídrico é indicado pela letra grega (psi) e é expresso em unidades de pressão (a pressão é uma forma de energia) chamadas megapascais (MPa). O potencial da água pura (w H2O puro) é, por conveniência de definição, designado como um valor zero (mesmo que a água pura contenha bastante energia potencial, essa energia é ignorada). Os valores do potencial de água para a água na raiz, caule ou folha de uma planta são, portanto, expressos em relação a w H2O puro.
O potencial hídrico nas soluções vegetais é influenciado pela concentração do soluto, pressão, gravidade e fatores chamados efeitos da matriz. O potencial hídrico pode ser dividido em seus componentes individuais usando a seguinte equação:
onde s, p, g e m referem-se aos potenciais de soluto, pressão, gravidade e matriz, respectivamente. “Sistema” pode se referir ao potencial hídrico da água do solo (solo), da água da raiz (raiz), da água do caule (caule), da água da folha (folha) ou da água na atmosfera (atmosfera): qualquer sistema aquoso que esteja sendo considerado. Conforme os componentes individuais mudam, eles aumentam ou diminuem o potencial total de água de um sistema. Quando isso acontece, a água se move para se equilibrar, movendo-se do sistema ou compartimento com maior potencial hídrico para o sistema ou compartimento com menor potencial hídrico. Isso traz a diferença no potencial hídrico entre os dois sistemas (Δ) de volta a zero (Δ= 0). Portanto, para que a água se mova pela planta do solo para o ar (um processo chamado transpiração), o solo deve estar > raiz > caule > folha > atmosfera.
A água só se move em resposta a Δ, não em resposta aos componentes individuais. No entanto, como os componentes individuais influenciam o sistema total, manipulando os componentes individuais (especialmente s), uma planta pode controlar o movimento da água.
Potencial de soluto
O potencial de soluto (s), também chamado de potencial osmótico, é negativo em uma célula vegetal e zero em água destilada. Os valores típicos para o citoplasma celular são de —0,5 a —1,0 MPa. Os solutos reduzem o potencial hídrico (resultando em w negativo) consumindo parte da energia potencial disponível na água. As moléculas de soluto podem se dissolver na água porque as moléculas de água podem se ligar a elas por meio de ligações de hidrogênio; uma molécula hidrofóbica como o óleo, que não pode se ligar à água, não pode entrar em solução. A energia nas ligações de hidrogênio entre as moléculas de soluto e a água não está mais disponível para funcionar no sistema porque está ligada à ligação. Em outras palavras, a quantidade de energia potencial disponível é reduzida quando os solutos são adicionados a um sistema aquoso. Assim, s diminui com o aumento da concentração de soluto. Como s é um dos quatro componentes do sistema ou total, uma diminuição em s causará uma diminuição no total de. O potencial hídrico interno de uma célula vegetal é mais negativo do que a água pura devido ao alto teor de soluto do citoplasma (Figura\(\PageIndex{2}\)). Devido a essa diferença no potencial hídrico, a água se moverá do solo para as células radiculares de uma planta por meio do processo de osmose. É por isso que o potencial de soluto às vezes é chamado de potencial osmótico.
As células vegetais podem manipular metabolicamente s (e, por extensão, total) adicionando ou removendo moléculas de soluto. Portanto, as plantas têm controle sobre total por meio de sua capacidade de exercer controle metabólico sobre s.
Conexão artística
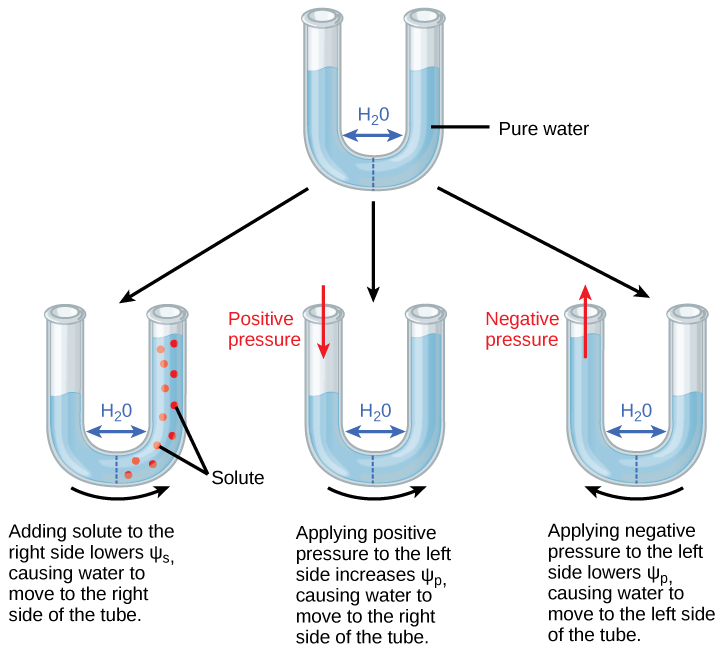
O potencial positivo de água é colocado no lado esquerdo do tubo aumentando p de forma que o nível da água suba no lado direito. Você poderia igualar o nível da água em cada lado do tubo adicionando soluto e, em caso afirmativo, como?
potencial de pressão
O potencial de pressão (p), também chamado de potencial de turgor, pode ser positivo ou negativo (Figura\(\PageIndex{3}\)). Como a pressão é uma expressão de energia, quanto maior a pressão, maior a energia potencial em um sistema e vice-versa. Portanto, um P positivo (compressão) aumenta total, e um p negativo (tensão) diminui total. A pressão positiva dentro das células é contida pela parede celular, produzindo pressão de turgor. Os potenciais de pressão normalmente estão em torno de 0,6—0,8 MPa, mas podem atingir até 1,5 MPa em uma planta bem regada. A p de 1,5 MPa equivale a 210 libras por polegada quadrada (1,5 MPa x 140 lb em -2 MPa -1 = 210 lb/in -2). Como comparação, a maioria dos pneus de automóveis são mantidos a uma pressão de 30 a 34 psi. Um exemplo do efeito da pressão do turgor é o murchamento das folhas e sua restauração após a rega da planta (Figura\(\PageIndex{3}\)). A água é perdida das folhas por transpiração (aproximando-se de p = 0 MPa no ponto de murchidão) e restaurada por absorção pelas raízes.
Uma planta pode manipular p por meio de sua capacidade de manipular s e pelo processo de osmose. Se uma célula vegetal aumentar a concentração de soluto citoplasmático, s diminuirá, total diminuirá, Δentre a célula e o tecido circundante diminuirá, a água entrará na célula por osmose e p aumentará. p também está sob controle indireto da planta por meio da abertura e fechamento dos estômatos. As aberturas estomáticas permitem que a água evapore da folha, reduzindo p e total da folha e aumentando ii entre a água na folha e o pecíolo, permitindo assim que a água flua do pecíolo para a folha.

Potencial de
O potencial gravitacional (g) é sempre negativo a zero em uma planta sem altura. Ele sempre remove ou consome energia potencial do sistema. A força da gravidade puxa a água para baixo até o solo, reduzindo a quantidade total de energia potencial na água da planta (total). Quanto mais alta a planta, mais alta a coluna de água e mais influente g se torna. Em escala celular e em plantas curtas, esse efeito é insignificante e facilmente ignorado. No entanto, acima da altura de uma árvore alta, como uma sequóia gigante costeira, a atração gravitacional de —0,1 MPa m -1 equivale a uma resistência extra de 1 MPa que deve ser superada para que a água alcance as folhas das árvores mais altas. As plantas são incapazes de manipular g.
Potencial matrício
O potencial matricial (m) é sempre negativo a zero. Em um sistema seco, pode ser tão baixo quanto —2 MPa em uma semente seca e zero em um sistema saturado de água. A ligação da água a uma matriz sempre remove ou consome energia potencial do sistema. m é semelhante ao potencial de soluto porque envolve a absorção da energia em um sistema aquoso formando ligações de hidrogênio entre a água e algum outro componente. No entanto, em potencial de soluto, os outros componentes são moléculas solúveis de soluto hidrofílico, enquanto em m, os outros componentes são moléculas hidrofílicas insolúveis da parede celular vegetal. Cada célula vegetal tem uma parede celular celulósica e a celulose nas paredes celulares é hidrofílica, produzindo uma matriz para adesão da água: daí o nome potencial matricial. m é muito grande (negativo) em tecidos secos, como sementes ou solos afetados pela seca. No entanto, ele rapidamente chega a zero quando a semente absorve água ou o solo se hidrata. m não pode ser manipulado pela planta e normalmente é ignorado em raízes, caules e folhas bem regados.
Movimento de água e minerais no Xilema
Solutos, pressão, gravidade e potencial matricial são importantes para o transporte de água nas plantas. A água se move de uma área de maior potencial total de água (maior energia livre de Gibbs) para uma área de menor potencial total de água. A energia livre de Gibbs é a energia associada a uma reação química que pode ser usada para trabalhar. Isso é expresso como Δ.
Transpiração é a perda de água da planta por evaporação na superfície da folha. É o principal motor do movimento da água no xilema. A transpiração é causada pela evaporação da água na interface folha-atmosfera; ela cria pressão negativa (tensão) equivalente a —2 MPa na superfície da folha. Esse valor varia muito dependendo do déficit de pressão de vapor, que pode ser insignificante em alta umidade relativa (UR) e substancial em baixa umidade relativa. A água das raízes é puxada para cima por essa tensão. À noite, quando os estômatos se fecham e a transpiração cessa, a água é retida no caule e na folha pela adesão da água às paredes celulares dos vasos do xilema e traqueídeos e pela coesão das moléculas de água umas às outras. Isso é chamado de teoria da coesão-tensão da ascensão da seiva.
Dentro da folha, no nível celular, a água na superfície das células mesofilas satura as microfibrilas de celulose da parede celular primária. A folha contém muitos grandes espaços de ar intercelulares para a troca de oxigênio por dióxido de carbono, que é necessário para a fotossíntese. A parede celular úmida é exposta ao espaço aéreo interno dessa folha, e a água na superfície das células evapora nos espaços aéreos, diminuindo a fina película na superfície das células do mesófilo. Essa diminuição cria uma maior tensão na água nas células do mesófilo (Figura\(\PageIndex{4}\)), aumentando assim a atração da água nos vasos do xilema. Os vasos do xilema e os traqueídeos são estruturalmente adaptados para lidar com grandes mudanças na pressão. Os anéis nos vasos mantêm sua forma tubular, assim como os anéis de uma mangueira de aspirador de pó mantêm a mangueira aberta enquanto está sob pressão. Pequenas perfurações entre os elementos do vaso reduzem o número e o tamanho das bolhas de gás que podem se formar por meio de um processo chamado cavitação. A formação de bolhas de gás no xilema interrompe o fluxo contínuo de água da base até o topo da planta, causando uma quebra denominada embolia no fluxo da seiva do xilema. Quanto mais alta a árvore, maiores as forças de tensão necessárias para puxar a água e mais eventos de cavitação. Em árvores maiores, as embolias resultantes podem obstruir os vasos do xilema, tornando-os não funcionais.
Conexão artística
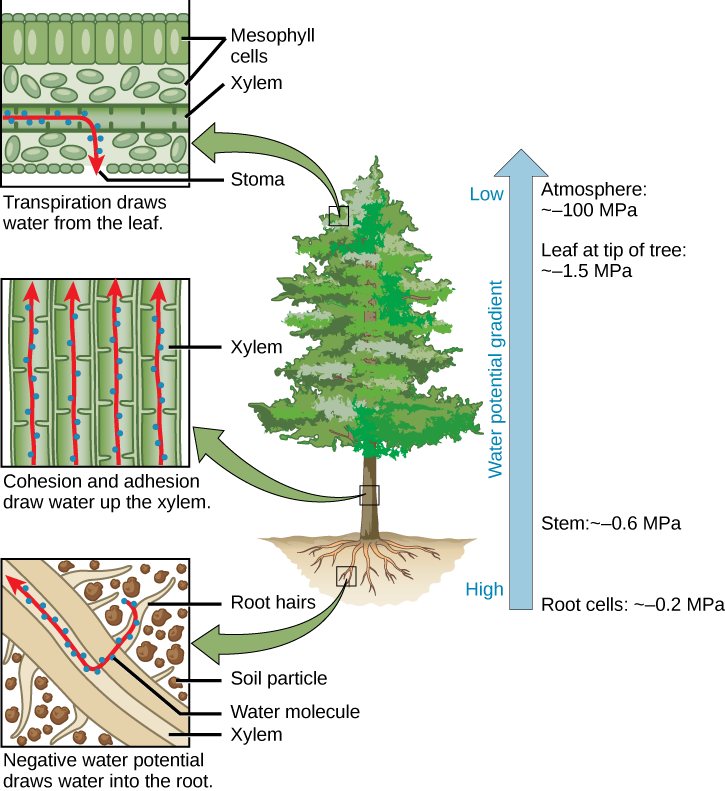
Qual das seguintes afirmações é falsa?
- O potencial negativo de água atrai a água para os pêlos das raízes. A coesão e a adesão aspiram água até o xilema. A transpiração retira água da folha.
- O potencial negativo de água atrai a água para os pêlos das raízes. A coesão e a adesão aspiram água até o floema. A transpiração retira água da folha.
- O potencial hídrico diminui das raízes até o topo da planta.
- A água entra nas plantas pelos pelos das raízes e sai pelo estoma.
A transpiração - a perda de vapor de água para a atmosfera por meio de estomas - é um processo passivo, o que significa que a energia metabólica na forma de ATP não é necessária para o movimento da água. A energia que impulsiona a transpiração é a diferença de energia entre a água no solo e a água na atmosfera. No entanto, a transpiração é rigidamente controlada.
Controle da transpiração
A atmosfera à qual a folha está exposta impulsiona a transpiração, mas também causa perda maciça de água da planta. Até 90 por cento da água absorvida pelas raízes pode ser perdida por meio da transpiração.
As folhas são cobertas por uma cutícula cerosa na superfície externa que evita a perda de água. A regulação da transpiração, portanto, é obtida principalmente por meio da abertura e fechamento dos estômatos na superfície da folha. Os estômatos são cercados por duas células especializadas chamadas células de guarda, que se abrem e fecham em resposta a sinais ambientais, como intensidade e qualidade da luz, estado da água nas folhas e concentrações de dióxido de carbono. Os estômatos devem se abrir para permitir que o ar contendo dióxido de carbono e oxigênio se difunda na folha para fotossíntese e respiração. Quando os estômatos estão abertos, no entanto, o vapor de água é perdido para o ambiente externo, aumentando a taxa de transpiração. Portanto, as plantas devem manter um equilíbrio entre a fotossíntese eficiente e a perda de água.
As plantas evoluíram ao longo do tempo para se adaptar ao ambiente local e reduzir a transpiração (Figura\(\PageIndex{5}\)). Plantas do deserto (xerófitas) e plantas que crescem em outras plantas (epífitas) têm acesso limitado à água. Essas plantas geralmente têm uma cutícula cerosa muito mais espessa do que aquelas que crescem em ambientes mais moderados e bem regados (mesófitos). As plantas aquáticas (hidrófitas) também têm seu próprio conjunto de adaptações anatômicas e morfológicas das folhas.

Xerófitas e epífitas geralmente têm uma espessa cobertura de tricomas ou de estômatos que estão afundados abaixo da superfície da folha. Os tricomas são células epidérmicas especializadas em forma de cabelo que secretam óleos e substâncias. Essas adaptações impedem o fluxo de ar pelo poro estomático e reduzem a transpiração. Várias camadas epidérmicas também são comumente encontradas nesses tipos de plantas.
Transporte de fotossintatos no floema
As plantas precisam de uma fonte de energia para crescer. Nas sementes e nos bulbos, os alimentos são armazenados em polímeros (como o amido) que são convertidos por processos metabólicos em sacarose para plantas recém-desenvolvidas. Uma vez que brotos e folhas verdes estão crescendo, as plantas são capazes de produzir seu próprio alimento por meio da fotossíntese. Os produtos da fotossíntese são chamados de fotossintatos, que geralmente estão na forma de açúcares simples, como a sacarose.
Estruturas que produzem fotossintatos para a planta em crescimento são chamadas de fontes. Os açúcares produzidos em fontes, como folhas, precisam ser entregues às partes em crescimento da planta por meio do floema em um processo chamado translocação. Os pontos de entrega de açúcar, como raízes, brotos jovens e sementes em desenvolvimento, são chamados de sumidouros. Sementes, tubérculos e bulbos podem ser uma fonte ou um sumidouro, dependendo do estágio de desenvolvimento da planta e da estação do ano.
Os produtos da fonte geralmente são translocados para a pia mais próxima através do floema. Por exemplo, as folhas mais altas enviarão fotossintatos para cima até a ponta do broto em crescimento, enquanto as folhas inferiores direcionarão os fotossintatos para baixo até as raízes. As folhas intermediárias enviarão produtos em ambas as direções, diferentemente do fluxo no xilema, que é sempre unidirecional (do solo para a folha para a atmosfera). O padrão do fluxo do fotossintato muda à medida que a planta cresce e se desenvolve. Os fotossintatos são direcionados principalmente às raízes desde o início, aos brotos e folhas durante o crescimento vegetativo e às sementes e frutos durante o desenvolvimento reprodutivo. Eles também são direcionados aos tubérculos para armazenamento.
Translocação: transporte da fonte para a pia
Os fotossintatos, como a sacarose, são produzidos nas células mesofilas das folhas fotossintetizadoras. A partir daí, eles são translocados pelo floema para onde são usados ou armazenados. As células mesofilas são conectadas por canais citoplasmáticos chamados plasmodesmas. Os fotossintatos se movem por esses canais para alcançar os elementos do tubo peneiro do floema (STEs) nos feixes vasculares. A partir das células do mesófilo, os fotossintatos são carregados no floema STeS. A sacarose é transportada ativamente contra seu gradiente de concentração (um processo que requer ATP) para as células do floema usando o potencial eletroquímico do gradiente de prótons. Isso é associado à absorção de sacarose com uma proteína transportadora chamada simportador de sacarose-H +.
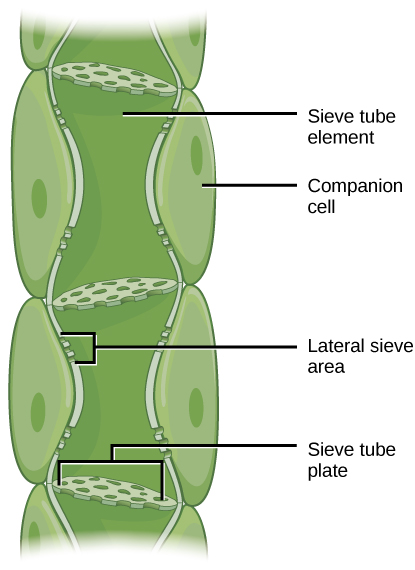
Os STEs de floema têm conteúdo citoplasmático reduzido e são conectados por uma placa de peneira com poros que permitem o fluxo a granel impulsionado por pressão, ou translocação, da seiva do floema. As células companheiras estão associadas aos STEs. Eles auxiliam nas atividades metabólicas e produzem energia para os STEs (Figura\(\PageIndex{6}\)).
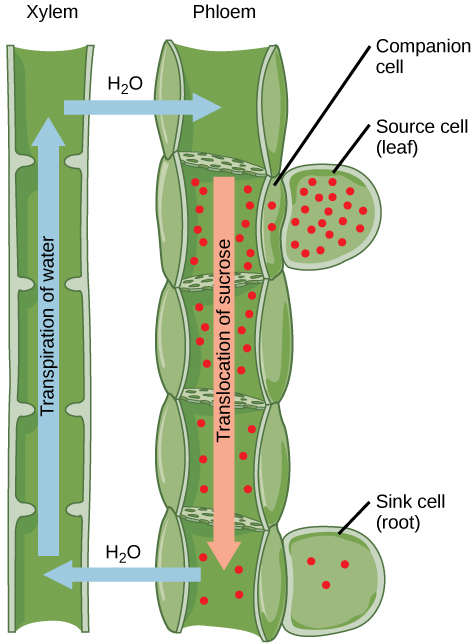
Uma vez no floema, os fotossintatos são translocados para a pia mais próxima. A seiva de floema é uma solução aquosa que contém até 30% de açúcar, minerais, aminoácidos e reguladores de crescimento vegetal. A alta porcentagem de açúcar diminui s, o que diminui o potencial total de água e faz com que a água se mova por osmose do xilema adjacente para os tubos do floema, aumentando assim a pressão. Esse aumento no potencial total de água causa o fluxo total do floema da fonte para o sumidouro (Figura\(\PageIndex{7}\)). A concentração de sacarose nas células coletoras é menor do que no floema STeS porque a sacarose do sumidouro foi metabolizada para crescimento ou convertida em amido para armazenamento ou outros polímeros, como celulose, para integridade estrutural. A descarga na extremidade do coletor do tubo floema ocorre por difusão ou transporte ativo de moléculas de sacarose de uma área de alta concentração para uma de baixa concentração. A água se difunde do floema por osmose e é então transpirada ou reciclada através do xilema de volta para a seiva do floema.
Resumo
O potencial hídrico () é uma medida da diferença de energia potencial entre uma amostra de água e água pura. O potencial hídrico em soluções vegetais é influenciado pela concentração de soluto, pressão, gravidade e potencial matricial. O potencial hídrico e a transpiração influenciam a forma como a água é transportada pelo xilema nas plantas. Esses processos são regulados pela abertura e fechamento estomáticos. Os fotossintatos (principalmente sacarose) se movem das fontes para os sumidouros através do floema da planta. A sacarose é carregada ativamente nos elementos do tubo de peneira do floema. O aumento da concentração de soluto faz com que a água se mova por osmose do xilema para o floema. A pressão positiva produzida empurra a água e os solutos para baixo no gradiente de pressão. A sacarose é descarregada na pia e a água retorna aos vasos do xilema.
Conexões artísticas
Figura\(\PageIndex{2}\): O potencial positivo de água é colocado no lado esquerdo do tubo aumentando p de forma que o nível da água suba no lado direito. Você poderia igualar o nível da água em cada lado do tubo adicionando soluto e, em caso afirmativo, como?
- Resposta
-
Sim, você pode equalizar o nível da água adicionando o soluto no lado esquerdo do tubo, de forma que a água se mova para a esquerda até que os níveis da água sejam iguais.
Figura\(\PageIndex{4}\): Qual das seguintes afirmações é falsa?
- O potencial negativo de água atrai a água para os pêlos das raízes. A coesão e a adesão aspiram água até o xilema. A transpiração retira água da folha.
- O potencial negativo de água atrai a água para os pêlos das raízes. A coesão e a adesão aspiram água até o floema. A transpiração retira água da folha.
- O potencial hídrico diminui das raízes até o topo da planta.
- A água entra nas plantas pelos pelos das raízes e sai pelo estoma.
- Resposta
-
B.
Glossário
- cutícula
- cobertura cerosa na parte externa da folha e do caule que evita a perda de água
- megapascal (MPa)
- unidades de pressão que medem o potencial da água
- pia
- partes em crescimento de uma planta, como raízes e folhas jovens, que requerem fotossintato
- fonte
- órgão que produz fotossintato para uma planta
- translocação
- transporte em massa de fotossintatos da fonte para o sumidouro em plantas vasculares
- transpiração
- perda de vapor de água para a atmosfera através de estômatos
- potencial hídrico (w)
- a energia potencial de uma solução de água por unidade de volume em relação à água pura à pressão atmosférica e à temperatura ambiente