
30.6: Sistemas sensoriais e respostas vegetais
- Page ID
- 182003

Habilidades para desenvolver
- Descreva como a luz vermelha e azul afetam o crescimento das plantas e as atividades metabólicas
- Discuta o gravitropismo
- Entenda como os hormônios afetam o crescimento e desenvolvimento das plantas
- Descreva o tigmotropismo, o tigmonastismo e a tigmogênese
- Explique como as plantas se defendem dos predadores e respondem às feridas
Os animais podem responder a fatores ambientais ao se mudarem para um novo local. As plantas, no entanto, estão enraizadas no local e devem responder aos fatores ambientais circundantes. As plantas têm sistemas sofisticados para detectar e responder à luz, gravidade, temperatura e toque físico. Os receptores detectam fatores ambientais e transmitem as informações aos sistemas efetores — geralmente por meio de mensageiros químicos intermediários — para gerar respostas da planta.
Respostas das plantas à luz
As plantas têm uma série de usos sofisticados da luz que vão muito além de sua capacidade de fotossintetizar açúcares de baixo peso molecular usando apenas dióxido de carbono, luz e água. A fotomorfogênese é o crescimento e desenvolvimento das plantas em resposta à luz. Ele permite que as plantas otimizem o uso da luz e do espaço. O fotoperiodismo é a capacidade de usar a luz para rastrear o tempo. As plantas podem saber a hora do dia e a época do ano detectando e usando vários comprimentos de onda da luz solar. O fototropismo é uma resposta direcional que permite que as plantas cresçam em direção ou mesmo longe da luz.
A detecção da luz no ambiente é importante para as plantas; pode ser crucial para a competição e a sobrevivência. A resposta das plantas à luz é mediada por diferentes fotorreceptores, que são compostos por uma proteína ligada covalentemente a um pigmento absorvente de luz chamado cromóforo. Juntos, os dois são chamados de cromoproteína.
As regiões vermelho/vermelho-distante e azul-violeta do espectro de luz visível desencadeiam o desenvolvimento estrutural nas plantas. Os fotorreceptores sensoriais absorvem a luz nessas regiões específicas do espectro de luz visível devido à qualidade da luz disponível no espectro diurno. Em habitats terrestres, a absorção de luz pelas clorofilas atinge o pico nas regiões azul e vermelha do espectro. À medida que a luz se filtra pelo dossel e os comprimentos de onda azul e vermelho são absorvidos, o espectro muda para a extremidade vermelha, mudando a comunidade vegetal para aquelas plantas mais bem adaptadas para responder à luz vermelha distante. Os receptores de luz azul permitem que as plantas avaliem a direção e a abundância da luz solar, que é rica em emissões azul-esverdeadas. A água absorve a luz vermelha, o que torna a detecção da luz azul essencial para algas e plantas aquáticas.
O sistema fitocromático e a resposta vermelho/vermelho distante
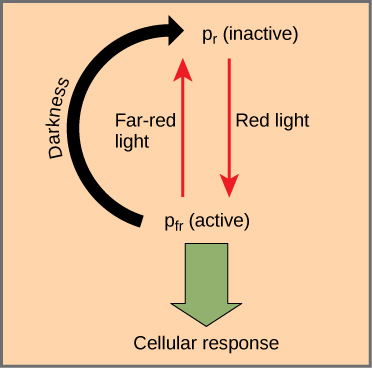
Os fitocromos são uma família de cromoproteínas com um cromóforo linear de tetrapirrol, semelhante ao grupo principal de clorofila absorvente de luz de tetrapirrol anelado. Os fitocromos têm duas formas foto-interconversíveis: Pr e Pfr. Pr absorve a luz vermelha (~ 667 nm) e é imediatamente convertido em Pfr. O Pfr absorve a luz vermelha distante (~ 730 nm) e é rapidamente convertido de volta para Pr. A absorção de luz vermelha ou vermelha distante causa uma mudança maciça na forma do cromóforo, alterando a conformação e a atividade da proteína fitocromática à qual ele está ligado. O Pfr é a forma fisiologicamente ativa da proteína; portanto, a exposição à luz vermelha produz atividade fisiológica. A exposição à luz vermelha distante inibe a atividade do fitocromo. Juntas, as duas formas representam o sistema fitocromático (Figura\(\PageIndex{1}\)).
O sistema fitocromático atua como um interruptor de luz biológico. Ele monitora o nível, intensidade, duração e cor da luz ambiental. O efeito da luz vermelha é reversível ao acender imediatamente uma luz vermelha distante na amostra, que converte a cromoproteína na forma inativa de Pr. Além disso, o Pfr pode reverter lentamente para Pr no escuro ou se decompor com o tempo. Em todos os casos, a resposta fisiológica induzida pela luz vermelha é revertida. A forma ativa do fitocromo (Pfr) pode ativar diretamente outras moléculas no citoplasma ou pode ser traficada para o núcleo, onde ativa ou reprime diretamente a expressão gênica específica.
Depois que o sistema fitocromático evoluiu, as plantas o adaptaram para atender a uma variedade de necessidades. A luz solar plena e não filtrada contém muito mais luz vermelha do que luz vermelha distante. Como a clorofila é absorvida fortemente na região vermelha do espectro visível, mas não na região do extremo vermelho, qualquer planta à sombra de outra planta no solo da floresta será exposta à luz vermelha esgotada e enriquecida com vermelho distante. A preponderância da luz vermelha distante converte o fitocromo nas folhas sombreadas na forma Pr (inativa), retardando o crescimento. As áreas não sombreadas (ou até menos sombreadas) mais próximas no solo da floresta têm mais luz vermelha; as folhas expostas a essas áreas sentem a luz vermelha, que ativa a forma Pfr e induz o crescimento. Resumindo, os brotos das plantas usam o sistema fitocromático para crescer longe da sombra e em direção à luz. Como a competição pela luz é tão acirrada em uma comunidade vegetal densa, as vantagens evolutivas do sistema fitocromático são óbvias.
Nas sementes, o sistema fitocromático não é usado para determinar a direção e a qualidade da luz (sombreada versus não sombreada). Em vez disso, é usado apenas para determinar se há alguma luz. Isso é especialmente importante em espécies com sementes muito pequenas, como a alface. Por causa de seu tamanho, as sementes de alface têm poucas reservas de alimentos. Suas mudas não podem crescer por muito tempo antes de ficarem sem combustível. Se germinassem até um centímetro abaixo da superfície do solo, a muda nunca chegaria à luz do sol e morreria. No escuro, o fitocromo está no Pr (forma inativa) e a semente não germinará; só germinará se exposta à luz na superfície do solo. Após a exposição à luz, o Pr é convertido em Pfr e a germinação prossegue.
As plantas também usam o sistema fitocromático para detectar a mudança de estação. O fotoperiodismo é uma resposta biológica ao horário e à duração do dia e da noite. Ele controla a floração, o aparecimento dos brotos de inverno e o crescimento vegetativo. A detecção de mudanças sazonais é crucial para a sobrevivência das plantas. Embora a temperatura e a intensidade da luz influenciem o crescimento das plantas, elas não são indicadores confiáveis da estação, pois podem variar de um ano para o outro. A duração do dia é um indicador melhor da época do ano.
Conforme mencionado acima, a luz solar não filtrada é rica em luz vermelha, mas deficiente em luz vermelha distante. Portanto, ao amanhecer, todas as moléculas de fitocromo em uma folha se convertem rapidamente na forma ativa de Pfr e permanecem nessa forma até o pôr do sol. No escuro, a forma Pfr leva horas para voltar lentamente à forma Pr. Se a noite for longa (como no inverno), toda a forma Pfr será revertida. Se a noite for curta (como no verão), uma quantidade considerável de Pfr pode permanecer ao nascer do sol. Ao detectar a relação Pr/Pfr ao amanhecer, uma planta pode determinar a duração do ciclo dia/noite. Além disso, as folhas retêm essas informações por vários dias, permitindo uma comparação entre a duração da noite anterior e as várias noites anteriores. Noites mais curtas indicam primavera para a planta; quando as noites ficam mais longas, o outono está se aproximando. Essas informações, juntamente com a detecção de temperatura e disponibilidade de água, permitem que as plantas determinem a época do ano e ajustem sua fisiologia de acordo. As plantas de dias curtos (noites longas) usam essas informações para florescer no final do verão e no início do outono, quando as noites excedem uma duração crítica (geralmente oito horas ou menos). As plantas de dias longos (noite curta) florescem durante a primavera, quando a escuridão é menor do que um comprimento crítico (geralmente de oito a 15 horas). Nem todas as plantas usam o sistema fitocromático dessa maneira. A floração em plantas neutras para o dia não é regulada pela duração do dia.
Conexão de carreira: horticultor
A palavra “horticultor” vem das palavras latinas para jardim (hortus) e cultura (cultura). Essa carreira foi revolucionada pelo progresso feito na compreensão das respostas das plantas aos estímulos ambientais. Os produtores de safras, frutas, vegetais e flores eram anteriormente limitados por terem que cronometrar a semeadura e a colheita de acordo com a estação. Agora, os horticultores podem manipular as plantas para aumentar a produção de folhas, flores ou frutos, entendendo como os fatores ambientais afetam o crescimento e o desenvolvimento das plantas.
O manejo de estufas é um componente essencial da educação de um horticultor. Para prolongar a noite, as plantas são cobertas com um pano de sombra opaco. As plantas de longa duração são irradiadas com luz vermelha no inverno para promover a floração precoce. Por exemplo, a luz fluorescente (branca fria) com comprimentos de onda azuis estimula o crescimento das folhas e é excelente para iniciar mudas. As lâmpadas incandescentes (lâmpadas padrão) são ricas em luz vermelha e promovem a floração em algumas plantas. O tempo de amadurecimento dos frutos pode ser aumentado ou retardado pela aplicação de hormônios vegetais. Recentemente, houve um progresso considerável no desenvolvimento de raças de plantas adequadas a diferentes climas e resistentes a pragas e danos causados pelo transporte. Tanto o rendimento quanto a qualidade das culturas aumentaram como resultado das aplicações práticas do conhecimento das respostas das plantas a estímulos e hormônios externos.
Os horticultores encontram emprego em laboratórios privados e governamentais, estufas, jardins botânicos e nas áreas de produção ou pesquisa. Eles melhoram as safras aplicando seus conhecimentos de genética e fisiologia vegetal. Para se preparar para uma carreira na horticultura, os alunos têm aulas de botânica, fisiologia vegetal, fitopatologia, paisagismo e melhoramento de plantas. Para complementar esses cursos tradicionais, os cursos de horticultura adicionam estudos em economia, negócios, ciência da computação e comunicação.
As respostas da luz azul
O fototropismo - a curvatura direcional de uma planta em direção ou para longe de uma fonte de luz - é uma resposta aos comprimentos de onda azuis da luz. O fototropismo positivo é o crescimento em direção a uma fonte de luz (Figura\(\PageIndex{2}\)), enquanto o fototropismo negativo (também chamado de eskototropismo) é o crescimento longe da luz.
As fototropinas apropriadamente chamadas são receptores à base de proteínas responsáveis por mediar a resposta fototrópica. Como todos os fotorreceptores vegetais, as fototrofinas consistem em uma porção de proteína e uma porção absorvente de luz, chamada de cromóforo. Nas fototrofinas, o cromóforo é uma molécula de flavina ligada covalentemente; portanto, as fototropinas pertencem a uma classe de proteínas chamadas flavoproteínas.
Outras respostas sob o controle das fototrofinas são a abertura e o fechamento das folhas, o movimento do cloroplasto e a abertura dos estômatos. No entanto, de todas as respostas controladas pelas fototropinas, o fototropismo foi estudado por mais tempo e é o mais bem compreendido.
Em seu tratado de 1880 O Poder dos Movimentos nas Plantas, Charles Darwin e seu filho Francis descreveram pela primeira vez o fototropismo como a inclinação das mudas em direção à luz. Darwin observou que a luz era percebida pela ponta da planta (o meristema apical), mas que a resposta (flexão) ocorria em uma parte diferente da planta. Eles concluíram que o sinal tinha que viajar do meristema apical até a base da planta.

Em 1913, Peter Boysen-Jensen demonstrou que um sinal químico produzido na ponta da planta era responsável pela flexão na base. Ele cortou a ponta de uma muda, cobriu a seção cortada com uma camada de gelatina e depois substituiu a ponta. A muda se inclinou em direção à luz quando iluminada. No entanto, quando flocos de mica impermeáveis foram inseridos entre a ponta e a base cortada, a plântula não se dobrou. Um refinamento do experimento mostrou que o sinal viajou pelo lado sombreado da muda. Quando a placa de mica foi inserida no lado iluminado, a planta se curvou em direção à luz. Portanto, o sinal químico era um estimulante de crescimento porque a resposta fototrópica envolvia um alongamento celular mais rápido no lado sombreado do que no lado iluminado. Agora sabemos que, à medida que a luz passa pelo caule de uma planta, ela é difratada e gera ativação de fototropina através do caule. A maioria das ativações ocorre no lado iluminado, fazendo com que o hormônio vegetal ácido indolacético (IAA) se acumule no lado sombreado. As células-tronco se alongam sob a influência do IAA.
Os criptocromos são outra classe de fotorreceptores absorvedores de luz azul que também contêm um cromóforo à base de flavina. Os criptocromos definem o ciclo de atividade de 24 horas da planta, também conhecido como ritmo circadiano, usando sinais de luz azul. Há algumas evidências de que os criptocromos trabalham em conjunto com as fototropinas para mediar a resposta fototrópica.
Link para o aprendizado
Use o menu de navegação no painel esquerdo deste site para ver imagens de plantas em movimento.
Respostas da planta à gravidade
Quer germinem ou não na luz ou na escuridão total, os brotos geralmente brotam do solo e as raízes crescem para baixo no solo. Uma planta colocada de lado no escuro enviará brotos para cima quando tiver tempo suficiente. O gravitropismo garante que as raízes cresçam no solo e que os brotos cresçam em direção à luz do sol. O crescimento da ponta apical da parte aérea para cima é chamado de gravitropismo negativo, enquanto o crescimento das raízes para baixo é chamado de gravitropismo positivo.
Os amiloplastos (também conhecidos como estatólitos) são plastídios especializados que contêm grânulos de amido e se depositam para baixo em resposta à gravidade. Os amiloplastos são encontrados em brotos e em células especializadas da capa radicular. Quando uma planta é inclinada, os estatólitos caem na nova parede celular inferior. Algumas horas depois, o broto ou a raiz mostrarão crescimento na nova direção vertical.
O mecanismo que medeia o gravitropismo é razoavelmente bem compreendido. Quando os amiloplastos se depositam no fundo das células sensíveis à gravidade na raiz ou na parte aérea, eles entram em contato físico com o retículo endoplasmático (ER), causando a liberação de íons de cálcio de dentro do pronto-socorro. Essa sinalização de cálcio nas células causa o transporte polar do hormônio vegetal IAA para o fundo da célula. Nas raízes, uma alta concentração de IAA inibe o alongamento celular. O efeito retarda o crescimento na parte inferior da raiz, enquanto as células se desenvolvem normalmente na parte superior. O IAA tem o efeito oposto nos brotos, onde uma maior concentração na parte inferior da parte aérea estimula a expansão celular, fazendo com que a parte aérea cresça. Depois que o broto ou a raiz começam a crescer verticalmente, os amiloplastos retornam à posição normal. Outras hipóteses - envolvendo a célula inteira no efeito do gravitropismo - foram propostas para explicar por que alguns mutantes que não têm amiloplastos ainda podem apresentar uma resposta gravitrópica fraca.
Respostas de crescimento
A resposta sensorial de uma planta a estímulos externos depende de mensageiros químicos (hormônios). Os hormônios vegetais afetam todos os aspectos da vida vegetal, desde a floração até a formação e maturação dos frutos, e do fototropismo à queda das folhas. Potencialmente, todas as células de uma planta podem produzir hormônios vegetais. Eles podem atuar em sua célula de origem ou ser transportados para outras partes do corpo vegetal, com muitas respostas vegetais envolvendo a interação sinérgica ou antagônica de dois ou mais hormônios. Em contraste, os hormônios animais são produzidos em glândulas específicas e transportados para um local distante para ação, e agem sozinhos.
Os hormônios vegetais são um grupo de substâncias químicas não relacionadas que afetam a morfogênese vegetal. Cinco hormônios vegetais principais são tradicionalmente descritos: auxinas (particularmente IAA), citocininas, giberelinas, etileno e ácido abscísico. Além disso, outros nutrientes e condições ambientais podem ser caracterizados como fatores de crescimento.
Auxinas
O termo auxina é derivado da palavra grega auxeína, que significa “crescer”. As auxinas são os principais hormônios responsáveis pelo alongamento celular no fototropismo e no gravitropismo. Eles também controlam a diferenciação do meristema em tecido vascular e promovem o desenvolvimento e o arranjo das folhas. Embora muitas auxinas sintéticas sejam usadas como herbicidas, a IAA é a única auxina natural que mostra atividade fisiológica. A dominância apical - a inibição da formação lateral do broto - é desencadeada pelas auxinas produzidas no meristema apical. A floração, o amadurecimento e o amadurecimento dos frutos e a inibição da abscisão (queda das folhas) são outras respostas das plantas sob o controle direto ou indireto das auxinas. As auxinas também atuam como um relé para os efeitos das respostas da luz azul e vermelho/vermelho distante.
O uso comercial de auxinas é generalizado em viveiros de plantas e para produção agrícola. O IAA é usado como um hormônio de enraizamento para promover o crescimento de raízes adventícias em estacas e folhas soltas. A aplicação de auxinas sintéticas em plantas de tomate em estufas promove o desenvolvimento normal dos frutos. A aplicação externa de auxina promove a sincronização da fixação e queda dos frutos para coordenar a época de colheita. Frutas como pepinos sem sementes podem ser induzidas a frutificar tratando flores de plantas não fertilizadas com auxinas.
Citocininas
O efeito das citocininas foi relatado pela primeira vez quando se descobriu que a adição do endosperma líquido do coco aos embriões vegetais em desenvolvimento em cultura estimulou seu crescimento. Verificou-se que o fator de crescimento estimulante é a citocinina, um hormônio que promove a citocinese (divisão celular). Quase 200 citocininas naturais ou sintéticas são conhecidas até o momento. As citocininas são mais abundantes nos tecidos em crescimento, como raízes, embriões e frutas, onde a divisão celular está ocorrendo. Sabe-se que as citocininas retardam a senescência nos tecidos foliares, promovem a mitose e estimulam a diferenciação do meristema em brotos e raízes. Muitos efeitos no desenvolvimento das plantas estão sob a influência das citocininas, seja em conjunto com a auxina ou outro hormônio. Por exemplo, a dominância apical parece resultar de um equilíbrio entre as auxinas que inibem as gemas laterais e as citocininas que promovem um crescimento mais denso.
Gibberelinas
As giberelinas (GAs) são um grupo de cerca de 125 hormônios vegetais intimamente relacionados que estimulam o alongamento dos brotos, a germinação das sementes e a maturação dos frutos e das flores. Os GAs são sintetizados nos meristemas apicais da raiz e do caule, nas folhas jovens e nos embriões das sementes. Em áreas urbanas, os antagonistas do GA às vezes são aplicados em árvores sob linhas de energia para controlar o crescimento e reduzir a frequência da poda.
Os GAs quebram a dormência (um estado de crescimento e desenvolvimento inibidos) nas sementes de plantas que precisam ser expostas ao frio ou à luz para germinar. O ácido abscísico é um forte antagonista da ação do GA. Outros efeitos dos GAs incluem a expressão de gênero, o desenvolvimento de frutos sem sementes e o atraso da senescência nas folhas e frutos. As uvas sem sementes são obtidas por meio de métodos de criação padrão e contêm sementes imperceptíveis que não se desenvolvem. Como os GAs são produzidos pelas sementes e porque o desenvolvimento dos frutos e o alongamento do caule estão sob controle do GA, essas variedades de uvas normalmente produziriam frutos pequenos em cachos compactos. As uvas maduras são tratadas rotineiramente com GA para promover frutos maiores, bem como cachos mais soltos (caules mais longos), o que reduz a ocorrência de infecção por mofo (Figura\(\PageIndex{3}\)).

Ácido abscísico
O hormônio vegetal ácido abscísico (ABA) foi descoberto pela primeira vez como o agente que causa a abscisão ou queda de cápsulas de algodão. No entanto, estudos mais recentes indicam que a ABA desempenha apenas um papel menor no processo de abscisão. O ABA se acumula como uma resposta a condições ambientais estressantes, como desidratação, temperaturas baixas ou duração reduzida do dia. Sua atividade contraria muitos dos efeitos promotores de crescimento de GAs e auxinas. O ABA inibe o alongamento do caule e induz dormência nas gemas laterais.
O ABA induz dormência nas sementes bloqueando a germinação e promovendo a síntese de proteínas de armazenamento. Plantas adaptadas a climas temperados requerem um longo período de temperatura fria antes que as sementes germinem. Esse mecanismo protege as plantas jovens de brotarem muito cedo durante o clima quente fora de época no inverno. À medida que o hormônio se decompõe gradualmente durante o inverno, a semente sai da dormência e germina quando as condições são favoráveis na primavera. Outro efeito do ABA é promover o desenvolvimento dos brotos de inverno; ele medeia a conversão do meristema apical em um broto adormecido. A baixa umidade do solo causa um aumento no ABA, o que faz com que os estômatos se fechem, reduzindo a perda de água nos brotos de inverno.
Etileno
O etileno está associado ao amadurecimento dos frutos, ao murchamento das flores e à queda das folhas. O etileno é incomum porque é um gás volátil (C 2 H 4). Centenas de anos atrás, quando lâmpadas de rua a gás foram instaladas nas ruas da cidade, as árvores que cresciam perto de postes de iluminação desenvolveram troncos torcidos e espessados e perderam suas folhas mais cedo do que o esperado. Esses efeitos foram causados pela volatilização do etileno das lâmpadas.
Tecidos envelhecidos (especialmente folhas senescentes) e nódulos de caules produzem etileno. O efeito mais conhecido do hormônio, entretanto, é a promoção do amadurecimento dos frutos. O etileno estimula a conversão de amido e ácidos em açúcares. Algumas pessoas armazenam frutas verdes, como abacates, em um saco de papel lacrado para acelerar o amadurecimento; o gás liberado pela primeira fruta a amadurecer acelerará a maturação da fruta restante. O etileno também desencadeia a abscisão das folhas e dos frutos, o desbotamento e a queda das flores e promove a germinação em alguns cereais e o surgimento de bulbos e batatas.
O etileno é amplamente utilizado na agricultura. Os fruticultores comerciais controlam o tempo de amadurecimento dos frutos com a aplicação do gás. Os horticultores inibem a queda de folhas em plantas ornamentais removendo o etileno das estufas usando ventiladores e ventilação.
Hormônios não tradicionais
Pesquisas recentes descobriram vários compostos que também influenciam o desenvolvimento das plantas. Seus papéis são menos compreendidos do que os efeitos dos principais hormônios descritos até agora.
Os jasmonatos desempenham um papel importante nas respostas de defesa à herbivoria. Seus níveis aumentam quando uma planta é ferida por um predador, resultando em um aumento nos metabólitos secundários tóxicos. Eles contribuem para a produção de compostos voláteis que atraem inimigos naturais dos predadores. Por exemplo, mastigar plantas de tomate por lagartas leva a um aumento nos níveis de ácido jasmônico, o que, por sua vez, desencadeia a liberação de compostos voláteis que atraem predadores da praga.
As oligossacarinas também desempenham um papel na defesa das plantas contra infecções bacterianas e fúngicas. Eles agem localmente no local da lesão e também podem ser transportados para outros tecidos. As estrigolactonas promovem a germinação das sementes em algumas espécies e inibem o desenvolvimento apical lateral na ausência de auxinas. As estrigolactonas também desempenham um papel no estabelecimento de micorrizas, uma associação mutualística de raízes de plantas e fungos. Os brassinoesteróides são importantes para muitos processos fisiológicos e de desenvolvimento. Sinais entre esses compostos e outros hormônios, principalmente auxina e GAs, amplificam seu efeito fisiológico. A dominância apical, a germinação das sementes, o gravitropismo e a resistência ao congelamento são todos influenciados positivamente pelos hormônios. O crescimento das raízes e a queda dos frutos são inibidos pelos esteróides.
Respostas da planta ao vento e ao toque
O broto de uma ervilha serpenteia em torno de uma treliça, enquanto uma árvore cresce em um ângulo em resposta aos fortes ventos predominantes. Esses são exemplos de como as plantas respondem ao toque ou ao vento.
O movimento de uma planta submetida à pressão direcional constante é chamado de tigmotropismo, das palavras gregas thigma, que significa “toque”, e tropismo, que significa “direção”. Gavinhas são um exemplo disso. A região meristemática das gavinhas é muito sensível ao toque; o toque leve evocará uma resposta rápida de enrolamento. As células em contato com uma superfície de suporte se contraem, enquanto as células do lado oposto do suporte se expandem. A aplicação de ácido jasmônico é suficiente para desencadear o enrolamento da gavinha sem um estímulo mecânico.
Uma resposta tigmonástica é uma resposta ao toque independente da direção do estímulo. Na armadilha de Vênus, duas folhas modificadas são unidas em uma dobradiça e forradas com dentes finos em forma de garfo ao longo das bordas externas. Pêlos minúsculos estão localizados dentro da armadilha. Quando um inseto escova esses pelos do gatilho, tocando dois ou mais deles em sucessão, as folhas se fecham rapidamente, prendendo a presa. As glândulas na superfície da folha secretam enzimas que digerem lentamente o inseto. Os nutrientes liberados são absorvidos pelas folhas, que reabrem para a próxima refeição.
A tigmomorfogênese é uma lenta mudança de desenvolvimento na forma de uma planta submetida a estresse mecânico contínuo. Quando as árvores se curvam com o vento, por exemplo, o crescimento geralmente é atrofiado e o tronco engrossa. O tecido fortalecedor, especialmente o xilema, é produzido para aumentar a rigidez e resistir à força do vento. Os pesquisadores levantam a hipótese de que a tensão mecânica induz crescimento e diferenciação para fortalecer os tecidos. O etileno e o jasmonato provavelmente estão envolvidos na tigmomorfogênese.
Link para o aprendizado

Use o menu à esquerda para navegar até três curtas-metragens: uma armadilha para mosca de Vênus capturando presas, o fechamento progressivo de folhetos de plantas sensíveis e o entrelaçamento de gavinhas.
Respostas de defesa contra herbívoros e patógenos
As plantas enfrentam dois tipos de inimigos: herbívoros e patógenos. Herbívoros, grandes e pequenos, usam plantas como alimento e as mastigam ativamente. Os patógenos são agentes da doença. Esses microrganismos infecciosos, como fungos, bactérias e nematóides, vivem da planta e danificam seus tecidos. As plantas desenvolveram uma variedade de estratégias para desencorajar ou matar atacantes.
A primeira linha de defesa nas plantas é uma barreira intacta e impenetrável. A casca e a cutícula cerosa podem proteger contra predadores. Outras adaptações contra a herbivoria incluem espinhos, que são galhos modificados, e espinhos, que são folhas modificadas. Eles desencorajam os animais, causando danos físicos e induzindo erupções cutâneas e reações alérgicas. A proteção externa de uma planta pode ser comprometida por danos mecânicos, que podem fornecer um ponto de entrada para patógenos. Se a primeira linha de defesa for violada, a planta deve recorrer a um conjunto diferente de mecanismos de defesa, como toxinas e enzimas.
Metabólitos secundários são compostos que não são derivados diretamente da fotossíntese e não são necessários para a respiração ou o crescimento e desenvolvimento das plantas. Muitos metabólitos são tóxicos e podem até ser letais para os animais que os ingerem. Alguns metabólitos são alcalóides, que desencorajam predadores com odores nocivos (como os óleos voláteis de menta e sálvia) ou sabores repelentes (como o amargor da quinina). Outros alcalóides afetam os herbívoros causando estimulação excessiva (a cafeína é um exemplo) ou a letargia associada aos opioides. Alguns compostos se tornam tóxicos após a ingestão; por exemplo, o cianeto de glicol na raiz da mandioca libera cianeto somente após a ingestão pelo herbívoro.
Ferimentos mecânicos e ataques de predadores ativam mecanismos de defesa e proteção tanto no tecido danificado quanto em locais mais distantes do local da lesão. Algumas reações de defesa ocorrem em minutos: outras em várias horas. As células infectadas e vizinhas podem morrer, impedindo assim a propagação da infecção.
A sinalização de longa distância provoca uma resposta sistêmica com o objetivo de dissuadir o predador. À medida que o tecido é danificado, os jasmonatos podem promover a síntese de compostos que são tóxicos para os predadores. Os jasmonatos também provocam a síntese de compostos voláteis que atraem parasitóides, que são insetos que passam seus estágios de desenvolvimento dentro ou sobre outro inseto e, eventualmente, matam seu hospedeiro. A planta pode ativar a abscisão do tecido lesionado se for danificada sem reparos.
Resumo
As plantas respondem à luz por meio de mudanças na morfologia e atividade. A irradiação por luz vermelha converte o fitocromo fotorreceptor em sua forma de absorção de luz vermelha distante - PFR. Essa forma controla a germinação e a floração em resposta à duração do dia, bem como desencadeia a fotossíntese em plantas dormentes ou que acabaram de emergir do solo. Receptores de luz azul, criptocromos e fototropinas são responsáveis pelo fototropismo. Os amiloplastos, que contêm grânulos pesados de amido, detectam a gravidade. Os brotos apresentam gravitropismo negativo, enquanto as raízes exibem gravitropismo positivo. Os hormônios vegetais - compostos naturais sintetizados em pequenas quantidades - podem atuar tanto nas células que os produzem quanto em tecidos e órgãos distantes. As auxinas são responsáveis pela dominância apical, crescimento radicular, crescimento direcional em direção à luz e muitas outras respostas de crescimento. As citocininas estimulam a divisão celular e combatem a dominância apical nas brotações. As giberelinas inibem a dormência das sementes e promovem o crescimento do caule. O ácido abscísico induz dormência em sementes e gemas e protege as plantas da perda excessiva de água ao promover o fechamento estomático. O gás etileno acelera o amadurecimento dos frutos e a queda das folhas. As plantas respondem ao toque por meio de movimentos rápidos (tigmotropia e timosteiro) e de crescimento diferencial lento (tigmomorfogênese). As plantas desenvolveram mecanismos de defesa contra predadores e patógenos. Barreiras físicas, como cascas e espinhos, protegem os tecidos sensíveis. As plantas também têm defesas químicas, incluindo metabólitos secundários tóxicos e hormônios, que provocam mecanismos adicionais de defesa.
Glossário
- ácido abscísico (ABA)
- hormônio vegetal que induz dormência em sementes e outros órgãos
- abscisão
- processo fisiológico que leva à queda de um órgão vegetal (como queda de folhas ou pétalas)
- auxina
- hormônio vegetal que influencia o alongamento celular (no fototropismo), o gravitropismo, a dominância apical e o crescimento radicular
- cromóforo
- molécula que absorve luz
- criptocromo
- proteína que absorve luz nas regiões azul e ultravioleta do espectro de luz
- citocinina
- hormônio vegetal que promove a divisão celular
- etileno
- hormônio vegetal volátil que está associado ao amadurecimento dos frutos, ao murchamento das flores e à queda das folhas
- giberelina (GA)
- hormônio vegetal que estimula o alongamento dos brotos, a germinação das sementes e a maturação e queda de frutas e flores
- jasmonatos
- pequena família de compostos derivados do ácido graxo ácido linoléico
- gravitropismo negativo
- crescimento longe da gravidade da Terra
- oligossacarina
- hormônio importante nas defesas das plantas contra infecções bacterianas e fúngicas
- fotomorfogênese
- crescimento e desenvolvimento de plantas em resposta à luz
- fotoperiodismo
- ocorrência de processos vegetais, como germinação e floração, de acordo com a época do ano
- fototropina
- receptor de luz azul que promove fototropismo, abertura e fechamento estomáticos e outras respostas que promovem a fotossíntese
- fototropismo
- flexão direcional de uma planta em direção a uma fonte de luz
- fitocromo
- proteína pigmentar vegetal que existe em duas formas reversíveis (Pr e Pfr) e medeia mudanças morfológicas em resposta à luz vermelha
- gravitropismo positivo
- crescimento em direção ao centro gravitacional da Terra
- estatólito
- (também, amiloplasto) organela vegetal que contém grânulos pesados de amido
- estrigolactona
- hormônio que promove a germinação de sementes em algumas espécies e inibe o desenvolvimento apical lateral na ausência de auxinas
- tigmomorfogênese
- resposta de desenvolvimento ao toque
- thigmonástico
- crescimento direcional de uma planta independente da direção na qual o contato é aplicado
- tigmotropismo
- crescimento direcional de uma planta em resposta ao contato constante