
9.2: Propagação do sinal
- Page ID
- 182184

Habilidades para desenvolver
- Explicar como a ligação de um ligante inicia a transdução do sinal por toda a célula
- Reconhecer o papel da fosforilação na transmissão de sinais intracelulares
- Avalie o papel dos segundos mensageiros na transmissão do sinal
Uma vez que um ligante se liga a um receptor, o sinal é transmitido através da membrana e para o citoplasma. A continuação de um sinal dessa maneira é chamada de transdução de sinal. A transdução de sinal ocorre apenas com receptores de superfície celular porque os receptores internos são capazes de interagir diretamente com o DNA no núcleo para iniciar a síntese de proteínas.
Quando um ligante se liga ao seu receptor, ocorrem mudanças conformacionais que afetam o domínio intracelular do receptor. Alterações conformacionais do domínio extracelular após a ligação do ligante podem se propagar pela região da membrana do receptor e levar à ativação do domínio intracelular ou de suas proteínas associadas. Em alguns casos, a ligação do ligante causa dimerização do receptor, o que significa que dois receptores se ligam um ao outro para formar um complexo estável chamado dímero. Um dímero é um composto químico formado quando duas moléculas (geralmente idênticas) se unem. A ligação dos receptores dessa maneira permite que seus domínios intracelulares entrem em contato próximo e se ativem mutuamente.
A vinculação inicia uma via de sinalização
Depois que o ligante se liga ao receptor da superfície celular, a ativação dos componentes intracelulares do receptor desencadeia uma cadeia de eventos chamada via de sinalização ou cascata de sinalização. Em uma via de sinalização, segundos mensageiros, enzimas e proteínas ativadas interagem com proteínas específicas, que por sua vez são ativadas em uma reação em cadeia que eventualmente leva a uma mudança no ambiente da célula (Figura\(\PageIndex{1}\)). Os eventos na cascata ocorrem em uma série, assim como uma corrente flui em um rio. As interações que ocorrem antes de um determinado ponto são definidas como eventos upstream, e os eventos posteriores a esse ponto são chamados de eventos downstream.
Conexão artística
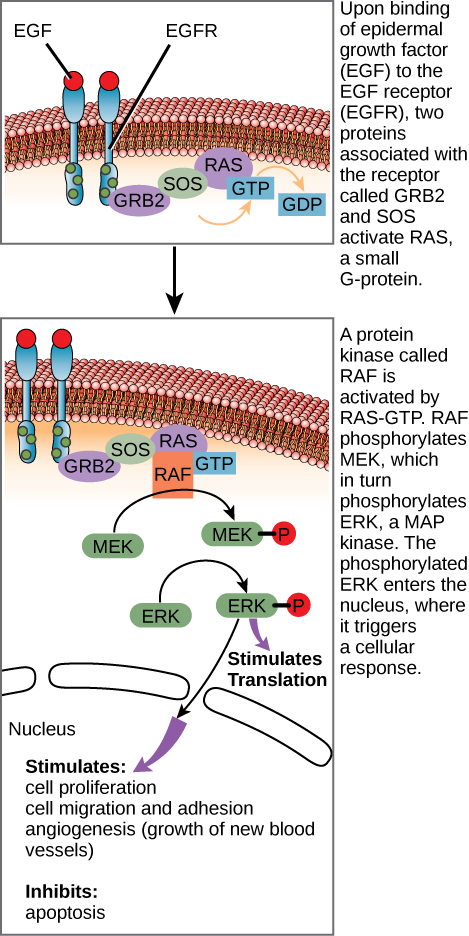
Em certos tipos de câncer, a atividade da GTPase da proteína G RAS é inibida. Isso significa que a proteína RAS não pode mais hidrolisar o GTP no PIB. Que efeito isso teria nos eventos celulares posteriores?
As vias de sinalização podem ficar muito complicadas muito rapidamente porque a maioria das proteínas celulares pode afetar diferentes eventos a jusante, dependendo das condições dentro da célula. Uma única via pode se ramificar em direção a diferentes endpoints com base na interação entre duas ou mais vias de sinalização, e os mesmos ligantes são frequentemente usados para iniciar sinais diferentes em diferentes tipos de células. Essa variação na resposta se deve às diferenças na expressão de proteínas em diferentes tipos de células. Outro elemento complicador é a integração de sinais das vias, na qual sinais de dois ou mais receptores diferentes da superfície celular se fundem para ativar a mesma resposta na célula. Esse processo pode garantir que vários requisitos externos sejam atendidos antes que uma célula se comprometa com uma resposta específica.
Os efeitos dos sinais extracelulares também podem ser amplificados por cascatas enzimáticas. No início do sinal, um único ligante se liga a um único receptor. No entanto, a ativação de uma enzima ligada ao receptor pode ativar muitas cópias de um componente da cascata de sinalização, que amplifica o sinal.
Métodos de sinalização intracelular
A indução de uma via de sinalização depende da modificação de um componente celular por uma enzima. Existem inúmeras modificações enzimáticas que podem ocorrer e elas são reconhecidas, por sua vez, pelo próximo componente a jusante. A seguir estão alguns dos eventos mais comuns na sinalização intracelular.
Fosforilação
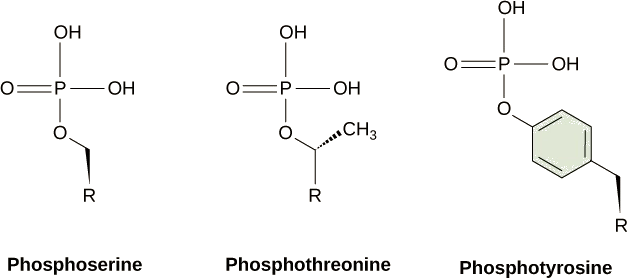
Uma das modificações químicas mais comuns que ocorrem nas vias de sinalização é a adição de um grupo fosfato (PO 4-3) a uma molécula como uma proteína em um processo chamado fosforilação. O fosfato pode ser adicionado a um nucleotídeo como o GMP para formar GDP ou GTP. Os fosfatos também são frequentemente adicionados aos resíduos de proteínas de serina, treonina e tirosina, onde substituem o grupo hidroxila do aminoácido (Figura\(\PageIndex{2}\)). A transferência do fosfato é catalisada por uma enzima chamada quinase. Várias quinases são nomeadas devido ao substrato que fosforilam. A fosforilação de resíduos de serina e treonina geralmente ativa enzimas. A fosforilação de resíduos de tirosina pode afetar a atividade de uma enzima ou criar um local de ligação que interage com componentes a jusante na cascata de sinalização. A fosforilação pode ativar ou inativar enzimas, e a reversão da fosforilação, a desfosforilação por uma fosfatase, reverterá o efeito.
Segundos mensageiros
Segundos mensageiros são pequenas moléculas que propagam um sinal após ele ter sido iniciado pela ligação da molécula sinalizadora ao receptor. Essas moléculas ajudam a espalhar um sinal pelo citoplasma, alterando o comportamento de certas proteínas celulares.
O íon cálcio é um segundo mensageiro amplamente utilizado. A concentração livre de íons de cálcio (Ca 2+) dentro de uma célula é muito baixa porque as bombas de íons na membrana plasmática usam continuamente adenosina-5'-trifosfato (ATP) para removê-la. Para fins de sinalização, o Ca 2+ é armazenado em vesículas citoplasmáticas, como o retículo endoplasmático, ou acessado de fora da célula. Quando a sinalização ocorre, os canais de íons de cálcio dependentes do ligante permitem que os níveis mais altos de Ca 2+ presentes fora da célula (ou nos compartimentos de armazenamento intracelular) fluam para o citoplasma, o que aumenta a concentração citoplasmática de Ca 2+. A resposta ao aumento do Ca 2+ varia, dependendo do tipo de célula envolvida. Por exemplo, nas células β do pâncreas, a sinalização de Ca 2+ leva à liberação de insulina e, nas células musculares, um aumento no Ca 2+ leva a contrações musculares.
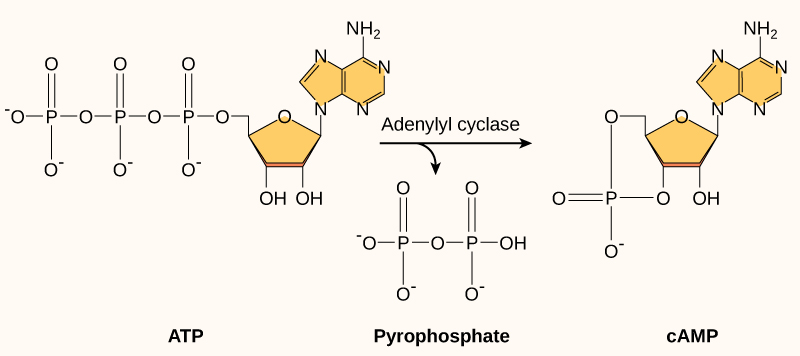
Outro segundo mensageiro utilizado em muitos tipos de células diferentes é o AMP cíclico (cAMP). O AMP cíclico é sintetizado pela enzima adenililciclase do ATP (Figura\(\PageIndex{3}\)). O principal papel do cAMP nas células é se ligar e ativar uma enzima chamada quinase dependente de CAMP (A-quinase). A A-quinase regula muitas vias metabólicas vitais: fosforila os resíduos de serina e treonina de suas proteínas-alvo, ativando-as no processo. A A-quinase é encontrada em muitos tipos diferentes de células, e as proteínas-alvo em cada tipo de célula são diferentes. As diferenças dão origem à variação das respostas ao cAMP em diferentes células.
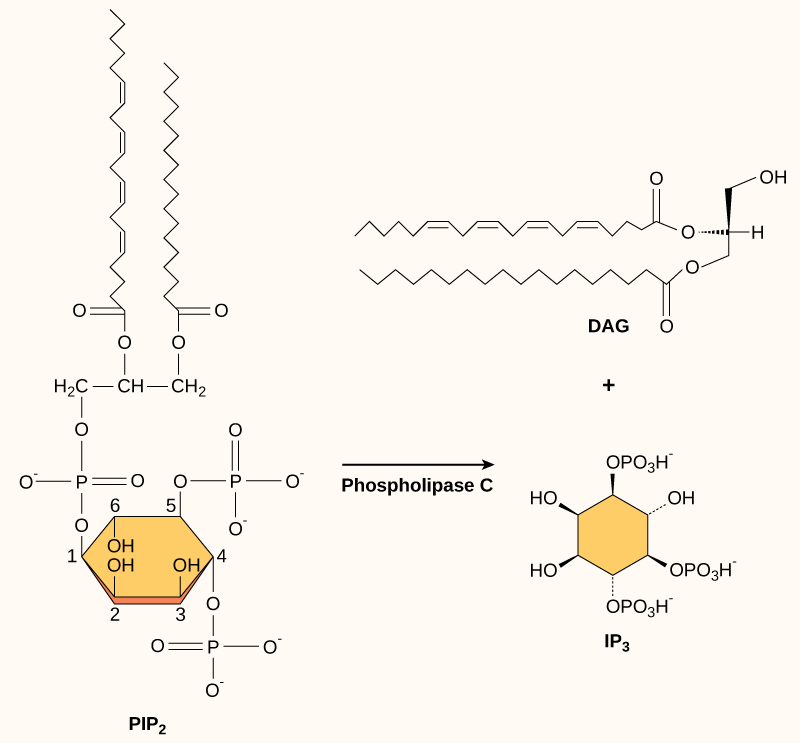
Presentes em pequenas concentrações na membrana plasmática, os fosfolipídios de inositol são lipídios que também podem ser convertidos em segundos mensageiros. Como essas moléculas são componentes da membrana, elas estão localizadas perto de receptores ligados à membrana e podem interagir facilmente com eles. O fosfatidilinositol (PI) é o principal fosfolipídio que desempenha um papel na sinalização celular. Enzimas conhecidas como fosforilato de quinases PI para formar fosfato de PI (PIP) e bisfosfato de PI (PIP 2).
A enzima fosfolipase C cliva o PIP 2 para formar diacilglicerol (DAG) e trifosfato de inositol (IP 3) (Figura\(\PageIndex{4}\)). Esses produtos da clivagem do PIP 2 servem como segundos mensageiros. O diacilglicerol (DAG) permanece na membrana plasmática e ativa a proteína quinase C (PKC), que então fosforila os resíduos de serina e treonina em suas proteínas-alvo. O IP 3 se difunde no citoplasma e se liga aos canais de cálcio dependentes do ligante no retículo endoplasmático para liberar Ca 2+, que continua a cascata de sinal.
Resumo
A ligação do ligante ao receptor permite a transdução do sinal através da célula. A cadeia de eventos que transmite o sinal através da célula é chamada de via de sinalização ou cascata. As vias de sinalização costumam ser muito complexas devido à interação entre diferentes proteínas. Um componente importante das cascatas de sinalização celular é a fosforilação de moléculas por enzimas conhecidas como quinases. A fosforilação adiciona um grupo fosfato aos resíduos de serina, treonina e tirosina em uma proteína, alterando suas formas e ativando ou inativando a proteína. Moléculas pequenas, como nucleotídeos, também podem ser fosforiladas. Os segundos mensageiros são moléculas pequenas, não proteicas, usadas para transmitir um sinal dentro de uma célula. Alguns exemplos de segundos mensageiros são íons de cálcio (Ca 2+), AMP cíclico (cAMP), diacilglicerol (DAG) e trifosfato de inositol (IP 3).
Conexões artísticas
Figura\(\PageIndex{1}\): Em certos tipos de câncer, a atividade da GTPase da proteína G RAS é inibida. Isso significa que a proteína RAS não pode mais hidrolisar o GTP no PIB. Que efeito isso teria nos eventos celulares posteriores?
- Resposta
-
O ERK se tornaria ativado permanentemente, resultando em proliferação celular, migração, adesão e crescimento de novos vasos sanguíneos. A apoptose seria inibida.
Glossário
- AMP cíclico (cAMP)
- segundo mensageiro derivado do ATP
- quinase cíclica dependente de AMP
- (também, proteína quinase A ou PKA) quinase que é ativada pela ligação ao cAMP
- diacilglicerol (DAG)
- produto de clivagem do PIP 2 que é usado para sinalização dentro da membrana plasmática
- dímero
- composto químico formado quando duas moléculas se unem
- dimerização
- (de proteínas receptoras) interação de duas proteínas receptoras para formar um complexo funcional chamado dímero
- fosfolipídio de inositol
- lipídio presente em pequenas concentrações na membrana plasmática que é convertido em um segundo mensageiro; tem inositol (um carboidrato) como seu grupo principal hidrofílico
- trifosfato de inositol (IP 3)
- produto de clivagem do PIP 2 que é usado para sinalização dentro da célula
- quinase
- enzima que catalisa a transferência de um grupo fosfato do ATP para outra molécula
- segundo mensageiro
- pequena molécula não proteica que propaga um sinal dentro da célula após a ativação de um receptor causa sua liberação
- integração de sinal
- interação de sinais de dois ou mais receptores diferentes da superfície celular que se fundem para ativar a mesma resposta na célula
- transdução de sinal
- propagação do sinal através do citoplasma (e às vezes também do núcleo) da célula
- via de sinalização
- (também cascata de sinalização) cadeia de eventos que ocorre no citoplasma da célula para propagar o sinal da membrana plasmática para produzir uma resposta