
9.1: Moléculas de sinalização e receptores celulares
- Page ID
- 182185

Habilidades para desenvolver
- Descreva quatro tipos de sinalização encontrados em organismos multicelulares
- Compare receptores internos com receptores de superfície celular
- Reconhecer a relação entre a estrutura de um ligante e seu mecanismo de ação
Existem dois tipos de comunicação no mundo das células vivas. A comunicação entre células é chamada de sinalização intercelular e a comunicação dentro de uma célula é chamada de sinalização intracelular. Uma maneira fácil de lembrar a distinção é entender a origem latina dos prefixos: inter- significa “entre” (por exemplo, linhas que se cruzam são aquelas que se cruzam) e intra- significa “dentro” (como intravenoso).
Os sinais químicos são liberados pelas células sinalizadoras na forma de moléculas pequenas, geralmente voláteis ou solúveis, chamadas ligantes. Um ligante é uma molécula que se liga a outra molécula específica, em alguns casos, emitindo um sinal no processo. Os ligantes podem, portanto, ser considerados moléculas sinalizadoras. Ligantes interagem com proteínas nas células-alvo, que são células afetadas por sinais químicos; essas proteínas também são chamadas de receptores. Ligantes e receptores existem em várias variedades; no entanto, um ligante específico terá um receptor específico que normalmente se liga apenas a esse ligante.
Formas de sinalização
Existem quatro categorias de sinalização química encontradas em organismos multicelulares: sinalização parácrina, sinalização endócrina, sinalização autócrina e sinalização direta através de junções gap (Figura\(\PageIndex{1}\)). A principal diferença entre as diferentes categorias de sinalização é a distância que o sinal percorre pelo organismo para alcançar a célula-alvo. Nem todas as células são afetadas pelos mesmos sinais.
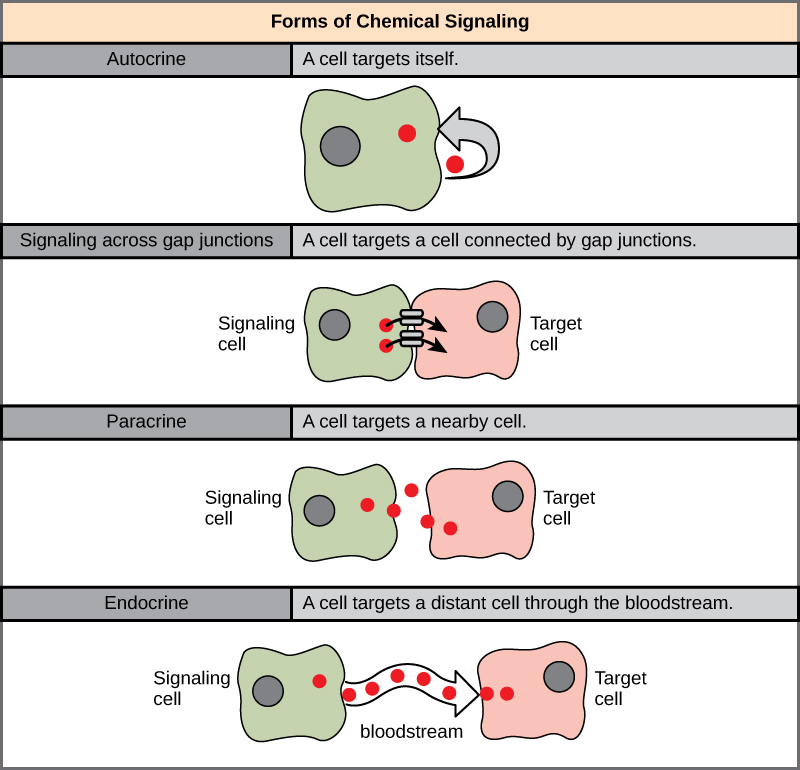
Sinalização parácrina
Sinais que agem localmente entre células próximas são chamados de sinais parácrinos. Sinais parácrinos se movem por difusão através da matriz extracelular. Esses tipos de sinais geralmente geram respostas rápidas que duram apenas um curto período de tempo. Para manter a resposta localizada, as moléculas de ligantes parácrinos normalmente são rapidamente degradadas pelas enzimas ou removidas pelas células vizinhas. A remoção dos sinais restabelecerá o gradiente de concentração do sinal, permitindo que eles se difundam rapidamente pelo espaço intracelular se liberados novamente.
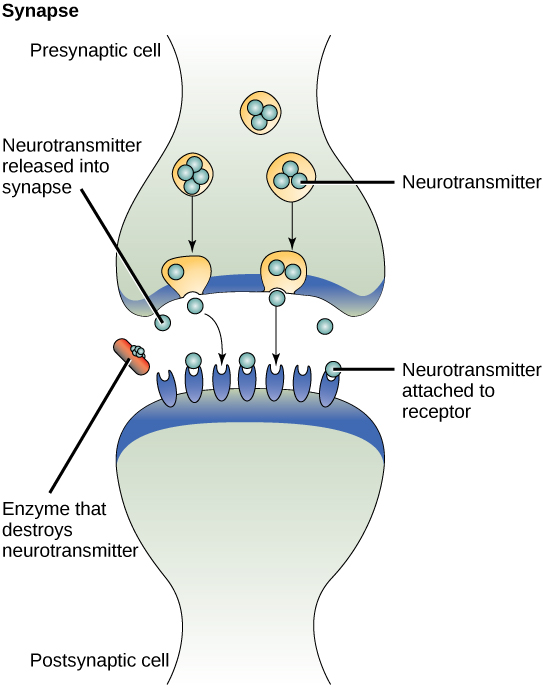
Um exemplo de sinalização parácrina é a transferência de sinais através de sinapses entre células nervosas. Uma célula nervosa consiste em um corpo celular, várias extensões curtas e ramificadas chamadas dendritos que recebem estímulos e uma extensão longa chamada axônio, que transmite sinais para outras células nervosas ou células musculares. A junção entre as células nervosas onde ocorre a transmissão do sinal é chamada de sinapse. Um sinal sináptico é um sinal químico que viaja entre as células nervosas. Os sinais dentro das células nervosas são propagados por impulsos elétricos em movimento rápido. Quando esses impulsos atingem o final do axônio, o sinal continua até um dendrito da próxima célula pela liberação de ligantes químicos chamados neurotransmissores pela célula pré-sináptica (a célula que emite o sinal). Os neurotransmissores são transportados por distâncias muito pequenas entre as células nervosas, chamadas sinapses químicas (Figura\(\PageIndex{2}\)). A pequena distância entre as células nervosas permite que o sinal viaje rapidamente; isso permite uma resposta imediata, como: Tire a mão do fogão!
Quando o neurotransmissor se liga ao receptor na superfície da célula pós-sináptica, o potencial eletroquímico da célula-alvo muda e o próximo impulso elétrico é lançado. Os neurotransmissores que são liberados na sinapse química são degradados rapidamente ou são reabsorvidos pela célula pré-sináptica para que a célula nervosa receptora possa se recuperar rapidamente e estar preparada para responder rapidamente ao próximo sinal sináptico.
Sinalização endócrina
Os sinais de células distantes são chamados de sinais endócrinos e se originam de células endócrinas. (No corpo, muitas células endócrinas estão localizadas nas glândulas endócrinas, como a glândula tireoidea, o hipotálamo e a glândula pituitária.) Esses tipos de sinais geralmente produzem uma resposta mais lenta, mas têm um efeito mais duradouro. Os ligantes liberados na sinalização endócrina são chamados de hormônios, moléculas sinalizadoras que são produzidas em uma parte do corpo, mas afetam outras regiões do corpo a alguma distância.
Os hormônios percorrem grandes distâncias entre as células endócrinas e suas células-alvo através da corrente sanguínea, que é uma forma relativamente lenta de se mover por todo o corpo. Por causa de sua forma de transporte, os hormônios se diluem e estão presentes em baixas concentrações quando atuam em suas células-alvo. Isso é diferente da sinalização parácrina, na qual as concentrações locais de ligantes podem ser muito altas.
Sinalização autócrina
Os sinais autócrinos são produzidos por células de sinalização que também podem se ligar ao ligante liberado. Isso significa que a célula de sinalização e a célula alvo podem ser iguais ou similares (o prefixo auto- significa self, um lembrete de que a célula de sinalização envia um sinal para si mesma). Esse tipo de sinalização geralmente ocorre durante o desenvolvimento inicial de um organismo para garantir que as células se desenvolvam nos tecidos corretos e assumam a função adequada. A sinalização autócrina também regula a sensação de dor e as respostas inflamatórias. Além disso, se uma célula estiver infectada com um vírus, ela pode sinalizar que sofre a morte celular programada, matando o vírus no processo. Em alguns casos, células vizinhas do mesmo tipo também são influenciadas pelo ligante liberado. No desenvolvimento embriológico, esse processo de estimulação de um grupo de células vizinhas pode ajudar a direcionar a diferenciação de células idênticas para o mesmo tipo celular, garantindo assim o resultado adequado do desenvolvimento.
Sinalização direta em junções de lacuna
As junções lacunas em animais e plasmodesmas em plantas são conexões entre as membranas plasmáticas das células vizinhas. Esses canais cheios de água permitem que pequenas moléculas sinalizadoras, chamadas mediadores intracelulares, se difundam entre as duas células. Moléculas pequenas, como íons de cálcio (Ca 2+), são capazes de se mover entre as células, mas moléculas grandes, como proteínas e DNA, não podem passar pelos canais. A especificidade dos canais garante que as células permaneçam independentes, mas possam transmitir sinais de forma rápida e fácil. A transferência de moléculas sinalizadoras comunica o estado atual da célula que está diretamente próxima à célula-alvo; isso permite que um grupo de células coordene sua resposta a um sinal que apenas uma delas pode ter recebido. Nas plantas, os plasmodesmas são onipresentes, transformando toda a planta em uma gigantesca rede de comunicação.
Tipos de receptores
Receptores são moléculas de proteína na célula alvo ou em sua superfície que se ligam ao ligante. Existem dois tipos de receptores, receptores internos e receptores de superfície celular.
Receptores internos
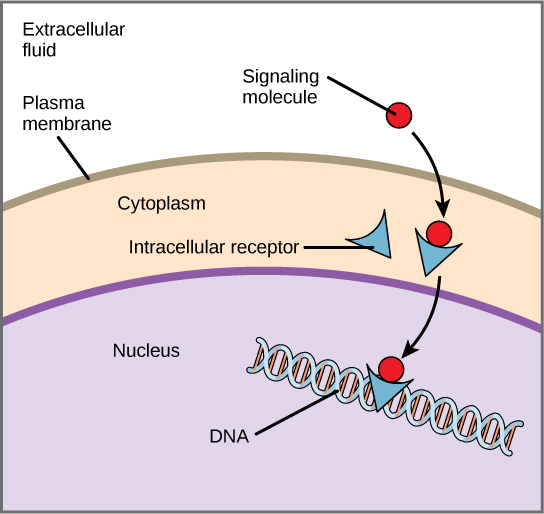
Os receptores internos, também conhecidos como receptores intracelulares ou citoplasmáticos, são encontrados no citoplasma da célula e respondem às moléculas de ligantes hidrofóbicos que são capazes de viajar pela membrana plasmática. Uma vez dentro da célula, muitas dessas moléculas se ligam a proteínas que atuam como reguladoras da síntese de mRNA (transcrição) para mediar a expressão gênica. A expressão gênica é o processo celular de transformar a informação no DNA de uma célula em uma sequência de aminoácidos, que, em última instância, forma uma proteína. Quando o ligante se liga ao receptor interno, uma mudança conformacional é acionada que expõe um sítio de ligação ao DNA na proteína. O complexo ligante-receptor se move para o núcleo, depois se liga a regiões reguladoras específicas do DNA cromossômico e promove o início da transcrição (Figura\(\PageIndex{3}\)). A transcrição é o processo de copiar as informações do DNA de uma célula em uma forma especial de RNA chamada RNA mensageiro (mRNA); a célula usa informações no mRNA (que se move para o citoplasma e se associa aos ribossomos) para ligar aminoácidos específicos na ordem correta, produzindo uma proteína. Os receptores internos podem influenciar diretamente a expressão gênica sem precisar passar o sinal para outros receptores ou mensageiros.
Receptores de superfície celular
Os receptores de superfície celular, também conhecidos como receptores transmembranares, são proteínas ancoradas na membrana (integrais) da superfície celular que se ligam a moléculas de ligantes externos. Esse tipo de receptor abrange a membrana plasmática e realiza a transdução de sinal, na qual um sinal extracelular é convertido em um sinal intercelular. Os ligantes que interagem com os receptores da superfície celular não precisam entrar na célula que afetam. Os receptores da superfície celular também são chamados de proteínas ou marcadores específicos da célula porque são específicos para tipos de células individuais.
Como as proteínas receptoras da superfície celular são fundamentais para o funcionamento normal da célula, não deve surpreender que um mau funcionamento de qualquer uma dessas proteínas possa ter consequências graves. Foi demonstrado que erros nas estruturas proteicas de certas moléculas receptoras desempenham um papel na hipertensão (pressão alta), asma, doenças cardíacas e câncer.
Cada receptor da superfície celular tem três componentes principais: um domínio externo de ligação ao ligante, uma região hidrofóbica que abrange a membrana e um domínio intracelular dentro da célula. O domínio de ligação ao ligante também é chamado de domínio extracelular. O tamanho e a extensão de cada um desses domínios variam muito, dependendo do tipo de receptor.
Evolution Connection: como vírus reconhecem um host
Ao contrário das células vivas, muitos vírus não têm uma membrana plasmática ou nenhuma das estruturas necessárias para sustentar a vida. Alguns vírus são simplesmente compostos por uma camada de proteína inerte contendo DNA ou RNA. Para se reproduzir, os vírus devem invadir uma célula viva, que serve como hospedeira, e então assumir o controle do aparato celular do hospedeiro. Mas como um vírus reconhece seu hospedeiro?
Os vírus geralmente se ligam aos receptores da superfície celular na célula hospedeira. Por exemplo, o vírus que causa a gripe humana se liga especificamente aos receptores nas membranas das células do sistema respiratório. Diferenças químicas nos receptores da superfície celular entre os hospedeiros significam que um vírus que infecta uma espécie específica (por exemplo, humanos) não pode infectar outra espécie (por exemplo, galinhas).
No entanto, os vírus têm quantidades muito pequenas de DNA ou RNA em comparação com os humanos e, como resultado, a reprodução viral pode ocorrer rapidamente. A reprodução viral invariavelmente produz erros que podem levar a alterações nos vírus recém-produzidos; essas mudanças significam que as proteínas virais que interagem com os receptores da superfície celular podem evoluir de tal forma que podem se ligar aos receptores em um novo hospedeiro. Essas mudanças acontecem aleatoriamente e com bastante frequência no ciclo reprodutivo de um vírus, mas as mudanças só importam se um vírus com novas propriedades de ligação entrar em contato com um hospedeiro adequado. No caso da gripe, essa situação pode ocorrer em ambientes onde animais e pessoas estão em contato próximo, como fazendas de aves e suínos. 1 Quando um vírus passa para um novo hospedeiro, ele pode se espalhar rapidamente. Cientistas observam de perto os vírus recém-surgidos (chamados vírus emergentes) na esperança de que esse monitoramento possa reduzir a probabilidade de epidemias virais globais.
Os receptores da superfície celular estão envolvidos na maior parte da sinalização em organismos multicelulares. Existem três categorias gerais de receptores de superfície celular: receptores ligados a canais iônicos, receptores ligados à proteína G e receptores ligados a enzimas.
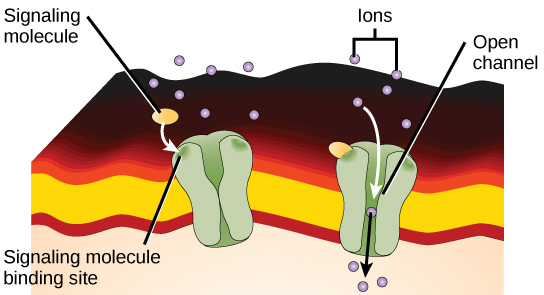
Os receptores ligados a canais iônicos se ligam a um ligante e abrem um canal através da membrana que permite a passagem de íons específicos. Para formar um canal, esse tipo de receptor de superfície celular tem uma extensa região de abrangência de membrana. Para interagir com as caudas de ácidos graxos fosfolipídicos que formam o centro da membrana plasmática, muitos dos aminoácidos na região que abrange a membrana são de natureza hidrofóbica. Por outro lado, os aminoácidos que revestem o interior do canal são hidrofílicos para permitir a passagem de água ou íons. Quando um ligante se liga à região extracelular do canal, há uma mudança conformacional na estrutura das proteínas que permite a passagem de íons como sódio, cálcio, magnésio e hidrogênio (Figura\(\PageIndex{4}\)).
Os receptores ligados à proteína G se ligam a um ligante e ativam uma proteína de membrana chamada proteína G. A proteína G ativada então interage com um canal iônico ou uma enzima na membrana (Figura\(\PageIndex{5}\)). Todos os receptores ligados à proteína G têm sete domínios transmembranares, mas cada receptor tem seu próprio domínio extracelular específico e sítio de ligação à proteína G.
A sinalização celular usando receptores ligados à proteína G ocorre como uma série cíclica de eventos. Antes que o ligante se ligue, a proteína G inativa pode se ligar a um local recém-revelado no receptor específico para sua ligação. Uma vez que a proteína G se liga ao receptor, a mudança de forma resultante ativa a proteína G, que libera PIB e capta GTP. As subunidades da proteína G então se dividem na subunidade α e na subunidade βγ. Como resultado, um ou ambos os fragmentos de proteína G podem ativar outras proteínas. Depois de algum tempo, o GTP na subunidade α ativa da proteína G é hidrolisado em PIB e a subunidade βγ é desativada. As subunidades se reassociam para formar a proteína G inativa e o ciclo recomeça.
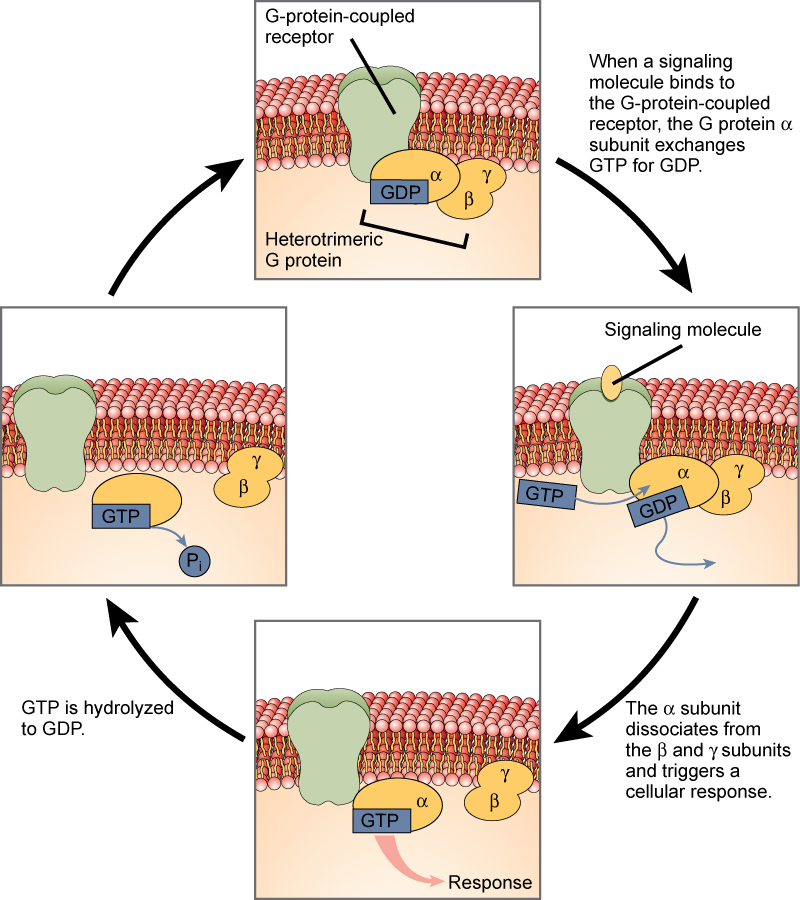
Os receptores ligados à proteína G foram extensivamente estudados e muito foi aprendido sobre seus papéis na manutenção da saúde. Bactérias patogênicas para humanos podem liberar venenos que interrompem a função específica do receptor ligado à proteína G, levando a doenças como coqueluche, botulismo e cólera. Na cólera (Figura\(\PageIndex{6}\)), por exemplo, a bactéria aquosa Vibrio cholerae produz uma toxina, o colerágeno, que se liga às células que revestem o intestino delgado. A toxina então entra nessas células intestinais, onde modifica uma proteína G que controla a abertura de um canal de cloreto e faz com que ela permaneça continuamente ativa, resultando em grandes perdas de fluidos do corpo e como resultado de uma desidratação potencialmente fatal.

Receptores ligados a enzimas são receptores de superfície celular com domínios intracelulares associados a uma enzima. Em alguns casos, o domínio intracelular do próprio receptor é uma enzima. Outros receptores ligados a enzimas têm um pequeno domínio intracelular que interage diretamente com uma enzima. Os receptores ligados a enzimas normalmente têm grandes domínios extracelulares e intracelulares, mas a região que abrange a membrana consiste em uma única região alfa-helicoidal da fita peptídica. Quando um ligante se liga ao domínio extracelular, um sinal é transferido pela membrana, ativando a enzima. A ativação da enzima desencadeia uma cadeia de eventos dentro da célula que eventualmente leva a uma resposta. Um exemplo desse tipo de receptor ligado a enzimas é o receptor de tirosina quinase (Figura\(\PageIndex{7}\)). A quinase é uma enzima que transfere grupos fosfato do ATP para outra proteína. O receptor da tirosina quinase transfere grupos fosfato para moléculas de tirosina (resíduos de tirosina). Primeiro, as moléculas sinalizadoras se ligam ao domínio extracelular de dois receptores de tirosina quinase próximos. Os dois receptores vizinhos então se unem ou dimerizam. Os fosfatos são então adicionados aos resíduos de tirosina no domínio intracelular dos receptores (fosforilação). Os resíduos fosforilados podem então transmitir o sinal para o próximo mensageiro dentro do citoplasma.
Conexão artística
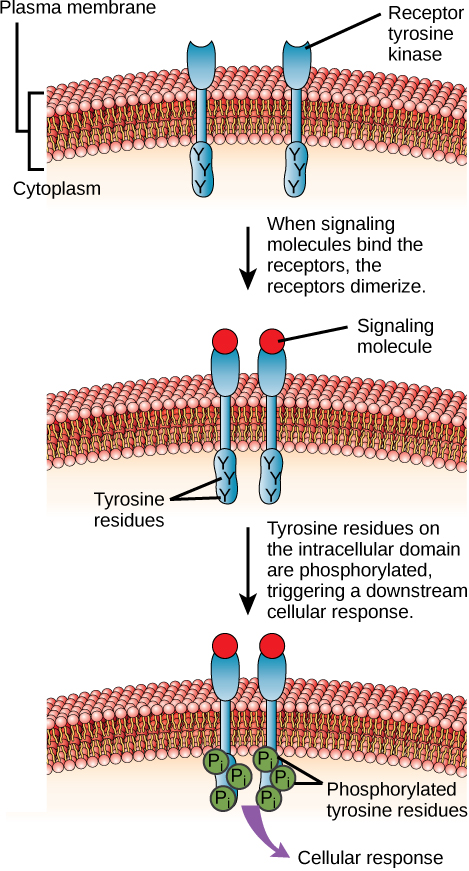
HER2 é um receptor de tirosina quinase. Em 30 por cento dos cânceres de mama humanos, o HER2 é ativado permanentemente, resultando em divisão celular não regulamentada. O lapatinibe, um medicamento usado para tratar o câncer de mama, inibe a autofosforilação da tirosina quinase do receptor HER2 (o processo pelo qual o receptor adiciona fosfatos a si mesmo), reduzindo assim o crescimento do tumor em 50 por cento. Além da autofosforilação, qual das seguintes etapas seria inibida pelo lapatinibe?
- Ligação de moléculas de sinalização, dimerização e resposta celular a jusante
- Dimerização e a resposta celular a jusante
- A resposta celular a jusante
- Atividade da fosfatase, dimerização e resposta celular a jusante
Moléculas de sinalização
Produzidos pelas células sinalizadoras e pela subsequente ligação aos receptores nas células-alvo, os ligantes atuam como sinais químicos que viajam até as células-alvo para coordenar as respostas. Os tipos de moléculas que servem como ligantes são incrivelmente variados e variam de pequenas proteínas a pequenos íons como cálcio (Ca 2+).
Ligantes hidrofóbicos pequenos
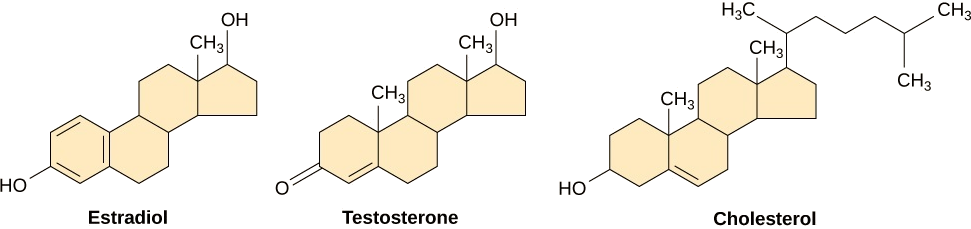
Pequenos ligantes hidrofóbicos podem se difundir diretamente pela membrana plasmática e interagir com receptores internos. Membros importantes dessa classe de ligantes são os hormônios esteróides. Os esteróides são lipídios que têm um esqueleto de hidrocarboneto com quatro anéis fundidos; diferentes esteróides têm diferentes grupos funcionais ligados ao esqueleto de carbono. Os hormônios esteróides incluem o hormônio sexual feminino, estradiol, que é um tipo de estrogênio; o hormônio sexual masculino, testosterona; e colesterol, que é um importante componente estrutural das membranas biológicas e um precursor dos hormônios esteróides (Figura\(\PageIndex{8}\)). Outros hormônios hidrofóbicos incluem hormônios tireoidianos e vitamina D. Para serem solúveis no sangue, os ligantes hidrofóbicos devem se ligar às proteínas transportadoras enquanto são transportados pela corrente sanguínea.
Ligantes solúveis em água
Ligantes solúveis em água são polares e, portanto, não podem passar pela membrana plasmática sem ajuda; às vezes, eles são muito grandes para passar pela membrana. Em vez disso, a maioria dos ligantes solúveis em água se liga ao domínio extracelular dos receptores da superfície celular. Esse grupo de ligantes é bastante diverso e inclui pequenas moléculas, peptídeos e proteínas.
Outros ligantes
O óxido nítrico (NO) é um gás que também atua como um ligante. É capaz de se difundir diretamente pela membrana plasmática e uma de suas funções é interagir com receptores no músculo liso e induzir o relaxamento do tecido. O NO tem uma meia-vida muito curta e, portanto, só funciona em curtas distâncias. A nitroglicerina, um tratamento para doenças cardíacas, atua desencadeando a liberação de NO, que faz com que os vasos sanguíneos se dilatem (expandam), restaurando assim o fluxo sanguíneo para o coração. O NO tornou-se mais conhecido recentemente porque a via que ele afeta é direcionada por medicamentos prescritos para disfunção erétil, como o Viagra (a ereção envolve vasos sanguíneos dilatados).
Resumo
As células se comunicam por sinalização inter e intracelular. As células sinalizadoras secretam ligantes que se ligam às células-alvo e iniciam uma cadeia de eventos dentro da célula-alvo. As quatro categorias de sinalização em organismos multicelulares são sinalização parácrina, sinalização endócrina, sinalização autócrina e sinalização direta nas junções gap. A sinalização parácrina ocorre em curtas distâncias. Os sinais endócrinos são transportados por longas distâncias pela corrente sanguínea pelos hormônios, e os sinais autócrinos são recebidos pela mesma célula que enviou o sinal ou por outras células próximas do mesmo tipo. As junções lacunas permitem que moléculas pequenas, incluindo moléculas sinalizadoras, fluam entre as células vizinhas.
Receptores internos são encontrados no citoplasma celular. Aqui, eles ligam moléculas de ligantes que atravessam a membrana plasmática; esses complexos receptor-ligante se movem para o núcleo e interagem diretamente com o DNA celular. Os receptores da superfície celular transmitem um sinal de fora da célula para o citoplasma. Os receptores ligados aos canais iônicos, quando ligados aos seus ligantes, formam um poro através da membrana plasmática por onde certos íons podem passar. Os receptores ligados à proteína G interagem com uma proteína G no lado citoplasmático da membrana plasmática, promovendo a troca de GDP ligado por GTP e interagindo com outras enzimas ou canais iônicos para transmitir um sinal. Receptores ligados a enzimas transmitem um sinal de fora da célula para um domínio intracelular de uma enzima ligada à membrana. A ligação ao ligante causa a ativação da enzima. Pequenos ligantes hidrofóbicos (como os esteróides) são capazes de penetrar na membrana plasmática e se ligar aos receptores internos. Ligantes hidrofílicos solúveis em água são incapazes de passar pela membrana; em vez disso, eles se ligam aos receptores da superfície celular, que transmitem o sinal para o interior da célula.
Conexões artísticas
Figura\(\PageIndex{7}\): HER2 é um receptor de tirosina quinase. Em 30 por cento dos cânceres de mama humanos, o HER2 é ativado permanentemente, resultando em divisão celular não regulamentada. O lapatinibe, um medicamento usado para tratar o câncer de mama, inibe a autofosforilação da tirosina quinase do receptor HER2 (o processo pelo qual o receptor adiciona fosfatos a si mesmo), reduzindo assim o crescimento do tumor em 50 por cento. Além da autofosforilação, qual das seguintes etapas seria inibida pelo lapatinibe?
- Ligação de moléculas de sinalização, dimerização e resposta celular a jusante.
- Dimerização e a resposta celular a jusante.
- A resposta celular a jusante.
- Atividade da fosfatase, dimerização e resposta celular a jusante.
- Responda
-
C. A resposta celular a jusante seria inibida.
Notas de pé
- 1 A. B. Sigalov, A Escola da Natureza. IV. Aprendendo com os vírus, a si mesmo 1, nº 4 (2010): 282-298. Y. Cao, X. Koh, L. Dong, X. Du, A. Wu, X. Ding, H. Deng, Y. Shu, J. Chen, T. Jiang, Estimativa rápida da atividade de ligação da hemaglutinina do vírus da gripe aos receptores humanos e aviários, PLoS One 6, no. 4 (2011): e18664.
Glossário
- sinal autócrino
- sinal que é enviado e recebido pelas mesmas células próximas ou similares
- receptor de superfície celular
- proteína da superfície celular que transmite um sinal do exterior da célula para o interior, mesmo que o ligante não entre na célula
- sinapse química
- pequeno espaço entre os terminais axônicos e os dendritos das células nervosas onde os neurotransmissores funcionam
- célula endócrina
- célula que libera ligantes envolvidos na sinalização endócrina (hormônios)
- sinal endócrino
- sinal de longa distância que é entregue por ligantes (hormônios) que viajam pelo sistema circulatório de um organismo da célula sinalizadora até a célula-alvo
- receptor ligado a enzimas
- receptor de superfície celular com domínios intracelulares associados a enzimas ligadas à membrana
- domínio extracelular
- região de um receptor de superfície celular que está localizado na superfície celular
- Receptor ligado à proteína G
- receptor de superfície celular que ativa proteínas G ligadas à membrana para transmitir um sinal do receptor para componentes de membrana próximos
- sinalização intercelular
- comunicação entre células
- receptor interno
- (também, receptor intracelular) proteína receptora que está localizada no citosol de uma célula e se liga a ligantes que passam pela membrana plasmática
- mediador intracelular
- (também, segundo mensageiro) molécula pequena que transmite sinais dentro de uma célula
- sinalização intracelular
- comunicação dentro das células
- receptor ligado a canais iônicos
- receptor de superfície celular que forma um canal de membrana plasmática, que se abre quando um ligante se liga ao domínio extracelular (canais dependentes do ligante)
- ligando
- molécula produzida por uma célula sinalizadora que se liga a um receptor específico, entregando um sinal no processo
- neurotransmissor
- ligante químico que transporta um sinal de uma célula nervosa para a próxima
- sinal parácrino
- sinal entre células próximas que é entregue por ligantes que viajam no meio líquido no espaço entre as células
- receptor
- proteína em ou sobre uma célula-alvo que se liga a ligantes
- célula de sinalização
- célula que libera moléculas de sinal que permitem a comunicação com outra célula
- sinal sináptico
- sinal químico (neurotransmissor) que viaja entre as células nervosas
- célula-alvo
- célula que tem um receptor para um sinal ou ligante de uma célula sinalizadora