
7.6: Conexões das vias metabólicas de carboidratos, proteínas e lipídios
- Page ID
- 182085

Habilidades para desenvolver
- Discuta as maneiras pelas quais as vias metabólicas dos carboidratos, a glicólise e o ciclo do ácido cítrico se inter-relacionam com as vias metabólicas de proteínas e lipídios
- Explique por que as vias metabólicas não são consideradas sistemas fechados
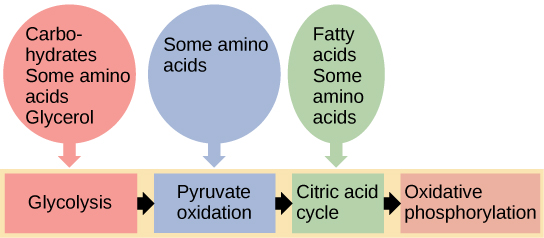
Você aprendeu sobre o catabolismo da glicose, que fornece energia às células vivas. Mas os seres vivos consomem mais do que glicose como alimento. Como um sanduíche de perú acaba como ATP em suas células? Isso acontece porque todas as vias catabólicas de carboidratos, proteínas e lipídios eventualmente se conectam à glicólise e às vias do ciclo do ácido cítrico (veja a Figura\(\PageIndex{2}\)). As vias metabólicas devem ser consideradas porosas, ou seja, as substâncias entram por outras vias e os intermediários partem para outras vias. Essas vias não são sistemas fechados. Muitos dos substratos, intermediários e produtos em uma via específica são reagentes em outras vias.
Conexões de outros açúcares com o metabolismo da glicose
O glicogênio, um polímero de glicose, é uma molécula de armazenamento de energia em animais. Quando há ATP adequado presente, o excesso de glicose é transferido para o glicogênio para armazenamento. O glicogênio é produzido e armazenado no fígado e no músculo. O glicogênio será hidrolisado em monômeros de glicose (G-1-P) se os níveis de açúcar no sangue caírem. A presença de glicogênio como fonte de glicose permite que o ATP seja produzido por um longo período de tempo durante o exercício. O glicogênio é decomposto em G-1-P e convertido em G-6-P nas células musculares e hepáticas, e esse produto entra na via glicolítica.
A sacarose é um dissacarídeo com uma molécula de glicose e uma molécula de frutose unidas com uma ligação glicosídica. A frutose é um dos três monossacarídeos dietéticos, junto com a glicose e a galactose (que faz parte do açúcar do leite, o dissacarídeo lactose), que são absorvidos diretamente na corrente sanguínea durante a digestão. O catabolismo da frutose e da galactose produz o mesmo número de moléculas de ATP que a glicose.
Conexões de proteínas ao metabolismo da glicose
As proteínas são hidrolisadas por uma variedade de enzimas nas células. Na maioria das vezes, os aminoácidos são reciclados para a síntese de novas proteínas. Se houver excesso de aminoácidos, no entanto, ou se o corpo estiver em estado de fome, alguns aminoácidos serão desviados para as vias do catabolismo da glicose (Figura\(\PageIndex{1}\)). Cada aminoácido deve ter seu grupo amino removido antes de entrar nessas vias. O grupo amino é convertido em amônia. Em mamíferos, o fígado sintetiza uréia a partir de duas moléculas de amônia e uma molécula de dióxido de carbono. Assim, a ureia é o principal produto residual dos mamíferos produzido a partir do nitrogênio originário dos aminoácidos e sai do corpo na urina.
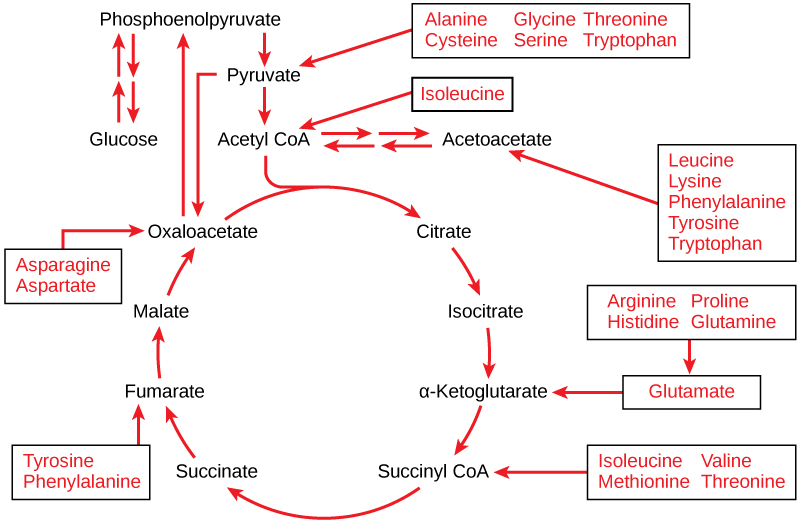
Conexões do metabolismo lipídico e da glicose
Os lipídios que estão conectados às vias da glicose são colesterol e triglicérides. O colesterol é um lipídio que contribui para a flexibilidade da membrana celular e é um precursor dos hormônios esteróides. A síntese do colesterol começa com grupos acetil e prossegue em apenas uma direção. O processo não pode ser revertido.
Os triglicérides são uma forma de armazenamento de energia a longo prazo em animais. Os triglicerídeos são feitos de glicerol e três ácidos graxos. Os animais podem produzir a maioria dos ácidos graxos de que precisam. Os triglicérides podem ser produzidos e decompostos por partes das vias de catabolismo da glicose. O glicerol pode ser fosforilado em glicerol-3-fosfato, que continua por meio da glicólise. Os ácidos graxos são catabolizados em um processo chamado beta-oxidação que ocorre na matriz das mitocôndrias e converte suas cadeias de ácidos graxos em duas unidades de carbono dos grupos acetil. Os grupos acetil são captados pelo CoA para formar o acetil CoA que prossegue no ciclo do ácido cítrico.
Conexão Evolutiva: Vias da fotossíntese e do metabolismo celular
Os processos de fotossíntese e metabolismo celular consistem em várias vias muito complexas. Geralmente, pensa-se que as primeiras células surgiram em um ambiente aquoso - uma “sopa” de nutrientes - provavelmente na superfície de algumas argilas porosas. Se essas células se reproduzissem com sucesso e seu número aumentasse de forma constante, as células começariam a esgotar os nutrientes do meio em que viviam à medida que transferiam os nutrientes para os componentes de seus próprios corpos. Essa situação hipotética teria resultado na seleção natural favorecendo os organismos que poderiam existir usando os nutrientes que permaneceram em seu ambiente e manipulando esses nutrientes em materiais sobre os quais poderiam sobreviver. A seleção favoreceria os organismos que pudessem extrair o máximo valor dos nutrientes aos quais tinham acesso.
Desenvolveu-se uma forma inicial de fotossíntese que aproveitou a energia do sol usando a água como fonte de átomos de hidrogênio, mas essa via não produzia oxigênio livre (fotossíntese anoxigênica). (A fotossíntese precoce não produzia oxigênio livre porque não usava água como fonte de íons de hidrogênio; em vez disso, usava materiais como sulfeto de hidrogênio e, consequentemente, produzia enxofre). Pensa-se que a glicólise se desenvolveu nessa época e poderia aproveitar os açúcares simples produzidos, mas essas reações não conseguiram extrair totalmente a energia armazenada nos carboidratos. O desenvolvimento da glicólise provavelmente antecedeu a evolução da fotossíntese, pois era adequada para extrair energia de materiais que se acumulavam espontaneamente na “sopa primitiva”. Uma forma posterior de fotossíntese usou a água como fonte de elétrons e hidrogênio e gerou oxigênio livre. Com o tempo, a atmosfera ficou oxigenada, mas não antes que o oxigênio liberasse metais oxidados no oceano e criasse uma camada de “ferrugem” no sedimento, permitindo a datação da ascensão dos primeiros fotossintetizadores oxigênicos. Coisas vivas se adaptaram para explorar essa nova atmosfera que permitiu que a respiração aeróbica como a conhecemos evoluísse. Quando todo o processo de fotossíntese oxigênica se desenvolveu e a atmosfera ficou oxigenada, as células finalmente conseguiram usar o oxigênio expelido pela fotossíntese para extrair consideravelmente mais energia das moléculas de açúcar usando o ciclo do ácido cítrico e a fosforilação oxidativa.
Resumo
A decomposição e síntese de carboidratos, proteínas e lipídios se conectam às vias do catabolismo da glicose. Os açúcares simples são galactose, frutose, glicogênio e pentose. Eles são catabolizados durante a glicólise. Os aminoácidos das proteínas se conectam com o catabolismo da glicose por meio do piruvato, acetil CoA e componentes do ciclo do ácido cítrico. A síntese do colesterol começa com grupos acetil, e os componentes dos triglicérides vêm do glicerol-3-fosfato da glicólise e dos grupos acetil produzidos nas mitocôndrias a partir do piruvato.