4.5: Evolução em ação - passado e presente
- Page ID
- 185688

Ao final desta seção, você poderá:
Os antropólogos biológicos contemporâneos utilizam uma perspectiva evolutiva. Isso significa que os princípios da evolução são usados para entender como e por que os organismos vivos, incluindo pessoas, prosperam em quase todos os ambientes da Terra. Mais especificamente, a seleção natural é aceita como a força orientadora que molda por que os seres vivos são do jeito que são. De todas as variações possíveis de seres competindo pelos mesmos recursos na Terra, aqueles que prosperaram foram os mais adequados a seus ambientes do que todos os outros concorrentes. Os princípios da evolução e da seleção natural serão discutidos com alguns detalhes nas próximas seções, mas é importante estabelecer, neste ponto inicial, que este capítulo se baseia na suposição fundamental de que as forças naturais são as únicas forças que direcionam o desenvolvimento da vida na Terra.
Os primeiros evolucionistas e a fixidade das espécies
A evolução é definida como uma mudança na frequência alélica dentro de um pool genético que pode levar a mudanças na morfologia (forma e estrutura) de um organismo ao longo do tempo. A evolução envolve os processos de mutação, seleção natural e especiação, que serão introduzidos nas próximas seções. Antes do século XIX, a ideia predominante no pensamento ocidental era que a natureza era fixa e estática; era feita por um ser supremo na forma em que aparecia atualmente e não mudou. Dentro desse sistema natural fixo, as criaturas vivas foram organizadas dentro de uma ordem definida que foi considerada decretada por Deus, conhecida como a grande cadeia do ser. Essa ordem apresentava Deus no topo, anjos abaixo de Deus e depois humanos. Abaixo dos humanos havia vários tipos de animais, seguidos por plantas e minerais. Essa hierarquia era significativa porque colocava algumas criaturas acima de outras quanto porque separava claramente os humanos do resto do mundo animal.
Durante um período que vai do século XIV ao XVIII, algumas pessoas começaram a questionar se o mundo natural era tão estático quanto era tradicionalmente percebido. O cientista e arquiteto britânico Robert Hooke é lembrado como a primeira pessoa no mundo ocidental a afirmar não apenas que a natureza mudou com o tempo, mas também que as evidências dessas mudanças permanecem. Ele levantou a hipótese de que os fósseis são restos de plantas e animais reais que já estiveram vivos. Essa conclusão foi contrária à conclusão anteriormente aceita de que fósseis nada mais eram do que imagens de pedra. Hooke também observou que muitos fósseis marinhos estavam localizados longe de qualquer oceano existente, e ele chegou à conclusão radical de que a geografia e as características físicas da Terra haviam sofrido mudanças dramáticas.
A primeira pessoa a propor um mecanismo pelo qual as espécies poderiam mudar foi o naturalista francês Jean-Baptiste Lamarck, mais conhecido por ter desenvolvido a primeira teoria da macroevolução, uma hipótese sobre como a transformação real de uma espécie em outra poderia ocorrer. A teoria de Lamarck se baseou na ideia agora extinta da herança das características adquiridas.
Lamarck argumentou que a utilidade de uma característica ou órgão poderia ser determinada com base em sua complexidade ou tamanho. Em particular, ele acreditava que a utilidade de um órgão poderia ser avaliada por seu tamanho e a utilidade de uma característica por sua complexidade. Ele especulou que órgãos e características que ajudam uma criatura a sobreviver se tornarão maiores e mais complexos com o tempo, enquanto aqueles que são de pouca utilidade se tornarão menores e mais simples e eventualmente desaparecerão. Seu exemplo clássico dessa teoria em ação é o pescoço comprido de uma girafa. Lamarck especulou que, à medida que as girafas estendiam o pescoço para alcançar as folhas no topo das árvores, seus pescoços cresceriam mais e, além disso, esses pescoços mais longos seriam herdados pelas gerações subsequentes. Essa teoria da herança das características adquiridas também é conhecida como herança lamarckiana. Uma das coisas interessantes sobre a teoria de Lamarck é que ele acreditava que desejos, desejos, vontades e necessidades eram suficientes para motivar a mudança. Ou seja, desejar ou desejar uma mudança nas características físicas de uma pessoa pode fazer com que essa mudança aconteça.
Existem dois problemas principais com a herança lamarckiana. Primeiro, desejos, desejos e necessidades não mudam as características físicas sem uma mudança deliberada no comportamento. Alguém pode desejar cabelos azuis, mas a cor do cabelo não mudará sem tinta. O segundo problema é que a herança das características adquiridas não é possível. Se alguém pintasse o cabelo de azul, seus filhos não herdariam o cabelo azul. As características que são adquiridas durante a vida não são transmitidas às gerações subsequentes.
Só porque a teoria da macroevolução de Lamarck não está correta não significa que ela seja insignificante. Lamarck reconheceu a importância das interações entre organismos e seus ambientes no processo evolutivo e foi o primeiro a propor um mecanismo pelo qual a mudança evolutiva de uma espécie para outra poderia realmente ocorrer.
Georges Cuvier, outro francês e cientista líder no início do século XIX, fez inúmeras contribuições ao pensamento evolutivo. Ele é mais conhecido por sua teoria do catastrofismo, que ele desenvolveu para explicar o número crescente de fósseis que estavam sendo encontrados, alguns mostrando impressões de criaturas que não eram mais encontradas em nenhum lugar da Terra. O catastrofismo propõe que inundações, terremotos e outros desastres naturais — entendidos na teoria como atos de Deus — foram responsáveis por matar todos os animais vivos em certos lugares em determinados momentos. De acordo com Cuvier, novos animais foram criados ou as áreas foram repovoadas por animais de áreas vizinhas. Para ser consistente com as evidências fósseis emergentes indicando que os organismos se tornaram mais complexos com o tempo, Cuvier propôs que novos organismos com uma aparência mais moderna fossem o resultado de um evento de criação mais recente. Embora os cientistas não adiram mais ao catastrofismo como uma teoria viável, a ideia de extinção de Cuvier continua sendo um componente importante do pensamento evolutivo hoje.
Outro grande contribuinte para o pensamento evolutivo foi o geólogo escocês Charles Lyell, conhecido como o pai da geologia moderna. Ele escreveu um tratado de três volumes, Princípios de Geologia (1830—1833), no qual argumentou que os processos geológicos contemporâneos eram os mesmos que ocorreram no passado. Esses processos, como vento e chuva, produziram a paisagem geológica contemporânea. Montanhas, lagos e rios foram todos criados por esses processos geológicos, muitos deles de movimento lento. Essa teoria ficou conhecida como o princípio do uniformitarismo. Lyell sugeriu que, para que essas forças de ação lenta produzam mudanças significativas, a Terra deve ser muito mais antiga do que se suspeitava. Antes da publicação de Lyell, a maioria dos historiadores naturais acreditava que a Terra tinha menos de 6.000 anos, um número obtido por meio de cálculos feitos com base no Antigo Testamento. Ao alterar a idade suspeita da Terra de vários milhares de anos para milhões de anos, Lyell mudou a estrutura dentro da qual os cientistas viam o passado geológico.
O papel de Charles Darwin na mudança de visões do mundo natural
Charles Darwin introduziu uma nova forma de ver o mundo que foi altamente criticada e aclamada na comunidade científica de sua época. Apesar da resistência de vários segmentos da sociedade, suas teorias da seleção natural se tornaram a base da ciência biológica. Novos conhecimentos relacionados à genética e à ciência molecular fortaleceram as teorias de Darwin em vez de enfraquecê-las.
Darwin, o Aprendiz
Quando tinha 17 anos, bem antes de ganhar a reputação de naturalista, acadêmico e cientista, Darwin estava estudando para ser médico na Universidade de Edimburgo. Como muitos jovens, ele começou a questionar sua escolha original de estudos e decidiu, em vez disso, aprender taxidermia com John Edmonstone. John Edmonstone nasceu escravizado e cresceu em uma plantação de propriedade de um político escocês no que hoje é a Guiana na América do Sul. Charles Waterton, genro do proprietário da plantação e renomado naturalista, visitava a plantação com frequência. Ele começou a convidar Edmonstone para acompanhá-lo em suas viagens frequentes pela floresta tropical. Em suas viagens, Edmonstone adquiriu um conhecimento considerável sobre a flora e a fauna da América do Sul, juntamente com impressionantes habilidades de taxidermia.
Depois de ganhar sua liberdade em 1817, John Edmonstone ensinou taxidermia na Universidade de Edimburgo, onde atuou como mentor de Darwin por um período de vários meses. Acredita-se que a relação de Darwin com Edmonstone possa ter influenciado suas visões abolicionistas, que mais tarde foram fortalecidas por relatos em primeira mão sobre escravidão enquanto Darwin estava em sua infame viagem às Ilhas Galápagos, na costa do Equador.

Darwin, o Explorador e Estudioso
Charles Darwin deixou a Universidade de Edimburgo e decidiu estudar teologia no Christ's College, Cambridge. Seus estudos lá levaram à sua nomeação em 1831 como naturalista no HMS Beagle para uma expedição científica de cinco anos ao redor do mundo. Durante essa viagem, Darwin coletou, dissecou e organizou vários espécimes, especialmente nas Ilhas Galápagos, uma cadeia de ilhas na costa oeste da América do Sul. Suas observações nas Galápagos marcaram um ponto crucial em seu pensamento sobre a evolução. Ele observou que a fauna e a flora da costa oeste da América do Sul eram semelhantes às observadas nas Galápagos, mas ainda assim distintas o suficiente para serem consideradas espécies diferentes. Mais surpreendentemente, os animais de cada uma das várias ilhas da cadeia de Galápagos diferiam um pouco uns dos outros. Darwin observou 13 tipos diferentes de tentilhões em 13 pequenas ilhas diferentes. Os pássaros de cada ilha diferiam na estrutura do bico, na forma corporal e na cor das penas. Cada espécie foi adaptada especificamente aos habitats específicos de cada uma das ilhas. Darwin usou as técnicas que Edmonstone lhe ensinou para preservar os tentilhões de Galápagos, que se tornaram peças-chave de evidência que apóiam a teoria da seleção natural de Darwin.

Durante suas viagens no Beagle, Darwin estava pensando na seleção artificial — a criação seletiva de animais para produzir características que os humanos consideram úteis, comumente associadas ao processo de domesticação. Darwin entendeu que a seleção artificial fornecia pistas importantes sobre a evolução natural das espécies.
Enquanto estava a bordo do HMS Beagle, Darwin leu um livro do economista inglês Thomas Robert Malthus intitulado An Essay on the Principle of Population (1798). Darwin obteve dois pontos importantes desse livro. A primeira foi que as populações humanas, se não forem restritas, crescerão exponencialmente. Isso significa que eles dobrarão a cada geração. O segundo ponto foi que os recursos alimentares aumentam muito mais lentamente do que a população. Malthus observou que o crescimento das populações humanas é controlado por um limite de recursos alimentares, o que cria uma luta pela existência. A luta pela existência não se trata apenas de conseguir comida suficiente, mas também de sobreviver. Em outras palavras, trata-se da capacidade de um indivíduo de encontrar comida suficiente e não se tornar o alimento de outro organismo. Esse conceito simples, a luta pela existência, forneceu a Darwin um mecanismo de como a evolução poderia ocorrer. Darwin percebeu que indivíduos com características favoráveis para viver em um ambiente são aqueles que sobreviverão até a idade em que se reproduzem, enquanto aqueles com variações menos favoráveis não. Esse mecanismo para “selecionar” certas características e características é conhecido como a teoria da seleção natural.
Darwin concluiu, a partir de suas observações, que quando um grupo de animais da mesma espécie é separado geologicamente, eles se transformam em espécies separadas. Esse processo evolutivo é comumente chamado de especiação alopátrica (ou especiação geográfica) e é baseado nos princípios de que espécies relacionadas compartilham um ancestral comum e que as espécies mudam com o tempo.
Darwin não deu origem à ideia de evolução. Muitas das ideias usadas por Darwin em sua teoria da seleção natural foram desenvolvidas por outros pensadores. Darwin também não era a única pessoa que pensava na seleção natural. Outro historiador natural britânico, Alfred Russel Wallace, desenvolveu a mesma ideia mais ou menos na mesma época, totalmente independente de Darwin. Enquanto Darwin desenvolveu suas ideias com base em suas viagens às Galápagos, o pensamento de Wallace foi influenciado por suas próprias viagens pelo arquipélago malaio entre a Indochina e a Austrália. Wallace delineou sua teoria da evolução por seleção natural em uma carta escrita para Darwin enquanto ele estava na Malásia. Como Darwin ainda não havia publicado seu próprio trabalho, Wallace e Darwin apresentaram em conjunto artigos que introduzem a teoria da seleção natural. Em 1859, Darwin finalmente publicou seu livro Sobre a Origem das Espécies, cerca de 20 anos após sua viagem no HMS Beagle.
Compreendendo a Teoria da Seleção Natural de Darwin
A teoria da seleção natural tem cinco componentes principais:
Um conceito popular, mas muitas vezes mal compreendido, relacionado à seleção natural é o termo sobrevivência do mais apto. A sobrevivência do mais apto não significa necessariamente que os maiores e mais rápidos sobrevivam; em vez disso, refere-se àqueles que estão evolutivamente mais aptos. Isso significa que um organismo tem características que são suficientes para sobreviver e serão transmitidas às gerações futuras. O termo sobrevivência do mais apto nem sequer foi introduzido por Darwin; ao contrário, foi usado pela primeira vez pelo filósofo, antropólogo e sociólogo inglês Herbert Spencer, que promoveu a ideologia agora desacreditada do darwinismo social. O darwinismo social aplicou o conceito da evolução biológica de Darwin às sociedades humanas, propondo que a cultura humana estava progredindo em direção ao “humano perfeito”. Os escritos de Spencer se tornaram integralmente relacionados à ascensão do racismo científico e do colonialismo europeu no século XIX.

Exemplos da teoria da seleção natural de Darwin podem ser encontrados em todo o mundo natural. Talvez uma das mais conhecidas seja a mudança de cor observada nas mariposas apimentadas na Inglaterra durante o século XIX. Antes da Revolução Industrial, as mariposas apimentadas na Inglaterra eram de cor cinza claro, bem camufladas nos galhos das árvores e menos propensas a serem comidas pelos pássaros. Ocasionalmente, por meio do processo de mutação, mariposas negras apareciam na população, mas geralmente eram comidas rapidamente porque eram mais visíveis contra cascas de cor clara. Quando a fuligem das fábricas de carvão começou a cobrir a casca das árvores, as mariposas negras ficaram mais camufladas e as brancas ficaram mais visíveis. Consequentemente, as mariposas negras sobreviveram para se reproduzir, enquanto as brancas foram comidas. Em algumas décadas, todas as mariposas apimentadas das cidades eram pretas. O processo foi denominado melanismo industrial. À medida que o uso de carvão diminuiu e a casca das árvores mais uma vez ficou mais clara, as mariposas brancas novamente dominaram as áreas urbanas.
Os exemplos de seleção natural nos tempos modernos são numerosos. A resistência a pesticidas em insetos é um exemplo clássico. A resistência a pesticidas se refere à diminuição da suscetibilidade de uma população de pragas a um pesticida que antes era eficaz em controlá-la. As espécies de pragas desenvolvem resistência a pesticidas por meio da seleção natural, com os indivíduos mais resistentes sobrevivendo para transmitir sua capacidade de resistir ao pesticida para seus filhotes. Outro bom exemplo é o aumento das “superbactérias”, bactérias que se tornaram cada vez mais resistentes aos antibióticos.
Os processos da evolução
A mutação é a força criativa da evolução e representa o primeiro estágio do processo evolutivo. A mutação é definida como uma alteração em uma sequência genética que resulta em uma forma variante. Para que uma mutação tenha significado evolutivo, ela deve ocorrer nas células sexuais (espermatozóides e óvulos). Isso ocorre porque apenas a informação genética que está nas células sexuais é transmitida de geração em geração. Mutações em cromossomos não sexuais não serão transmitidas de uma geração para a outra. Enquanto outras forças evolutivas podem modificar o material genético existente, somente a mutação pode produzir novo material genético. Uma das coisas mais interessantes sobre as mutações é o fato de serem aleatórias. Não há como prever quando uma mutação específica ocorrerá; tudo o que os cientistas podem fazer é estimar a probabilidade de uma mutação ocorrer. As mutações não aparecem necessariamente quando são necessárias.
A visão convencional é que as mutações são prejudiciais, mas isso nem sempre é verdade. Algumas mutações são prejudiciais, outras são vantajosas e outras são neutras. Mutações vantajosas levam a mudanças que melhoram a sobrevivência e/ou as chances de reprodução de um indivíduo. A mutação que confere resistência ao inseticida nos mosquitos levou a mudanças que melhoraram sua sobrevivência. Da mesma forma, a mutação da coloração preta em mariposas apimentadas levou a uma maior sobrevivência durante a Revolução Industrial. Mutações neutras não têm efeito na sobrevivência ou reprodução. E algumas mutações são, de fato, bastante prejudiciais e afetam negativamente a sobrevivência e a reprodução de certos indivíduos.
As mutações geralmente ocorrem espontaneamente em resposta às condições do corpo ou do meio ambiente. A causa exata de uma mutação geralmente não pode ser determinada e a taxa de mutação é muito difícil de determinar. Isso ocorre porque mutações que são neutras ou que não levam a mudanças óbvias geralmente passam despercebidas. A probabilidade de uma mutação em qualquer gene está entre 1 em 10.000 e 1 em 100.000. Embora a probabilidade de um ponto específico no material genético de um indivíduo ter uma mutação seja claramente muito baixa, a probabilidade de que a totalidade do material genético de um indivíduo tenha pelo menos uma mutação é muito maior. A questão é que, embora rara, a mutação também é comum. Por exemplo, embora muitos mosquitos tenham se adaptado aos inseticidas por meio de uma mutação que confere alguma resistência aos produtos químicos, se a mutação ainda não estivesse presente na população, os mosquitos teriam morrido. A necessidade de uma mutação específica não teve efeito sobre se a mutação apareceu ou não.
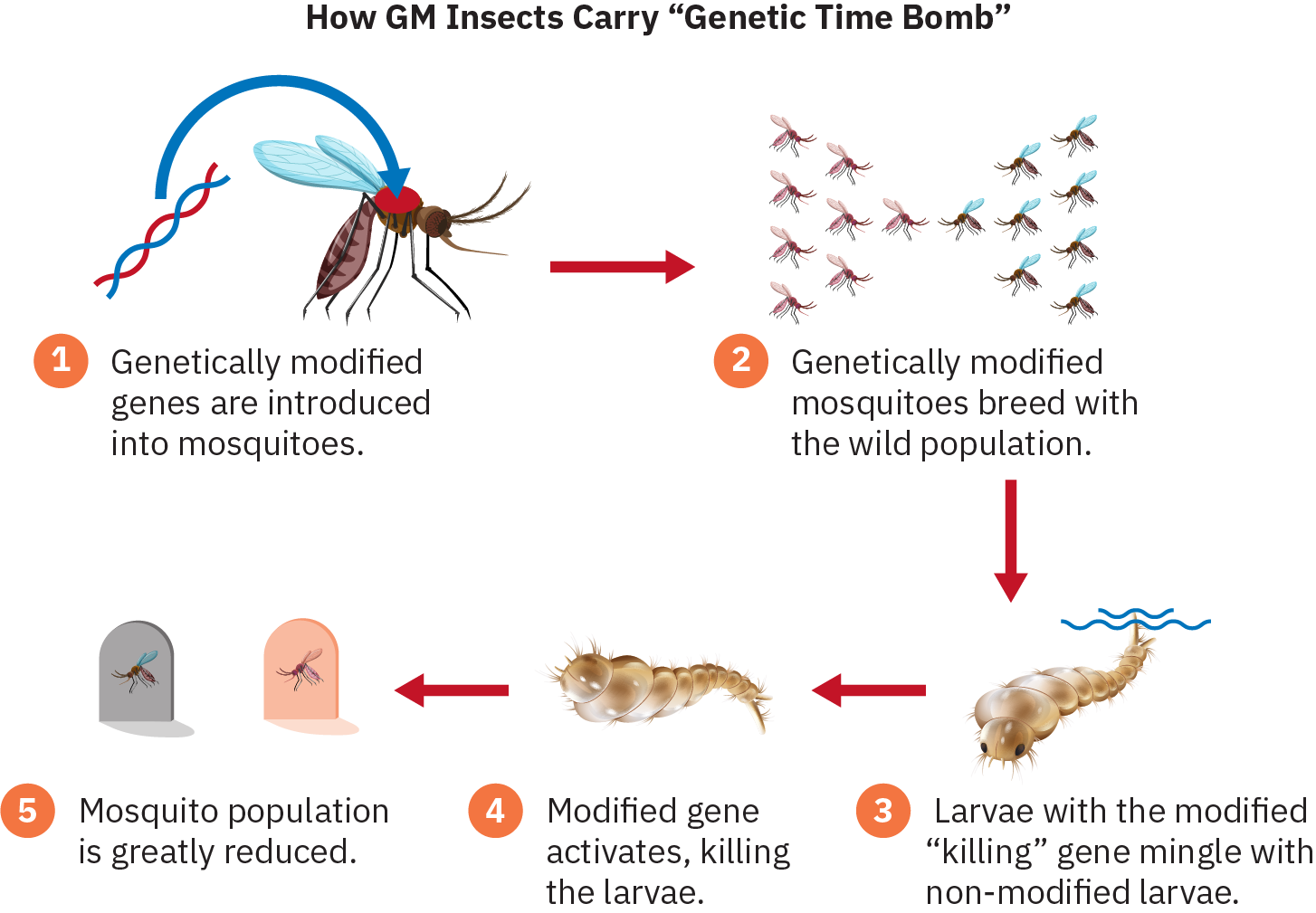
Atualmente, existe um polêmico programa piloto na Flórida com o objetivo de lidar com mosquitos contra os quais sprays de inseticidas estão se tornando cada vez mais ineficazes. Os primeiros mosquitos geneticamente modificados foram liberados em Florida Keys em maio de 2021. Os mosquitos geneticamente alterados produzem filhotes fêmeas que morrem na fase larval, impedindo que cresçam até a idade adulta, na qual podem picar e espalhar doenças. Atualmente, a ciência genética tem o poder de usar mutações para controlar ou até mesmo eliminar uma espécie inteira. A engenharia genética tem o potencial de beneficiar a humanidade, mas sem dúvida também levantará questões éticas e controvérsias.
Deriva genética
A deriva genética é definida como o efeito do acaso aleatório em uma população, principalmente a maneira pela qual ela determina se um indivíduo sobrevive, se reproduz ou morre. Imagine que você enfia a mão em um balde cheio de doces de Halloween. Qual é a probabilidade de você retirar uma barra de Snickers? A composição dos doces de Halloween em seu balde será afetada pela proporção de pessoas distribuindo barras de Snickers em comparação com outros doces. Se cada balde de doces de Halloween fosse uma população, pode-se dizer que a deriva genética - chance aleatória - estava afetando a composição do doce em seu balde de Halloween. Um ponto importante sobre a deriva genética é que ela está direta e inversamente relacionada ao tamanho da população. Quanto menor a população, maior a influência da deriva genética; quanto maior a população, menor a influência da deriva genética. Em uma grande população, digamos 100.000, a remoção de alguns indivíduos terá um efeito verdadeiramente minúsculo na população. Observe que, no início da evolução humana, no entanto, o tamanho da população era pequeno, então o efeito da deriva genética pode ter sido substancial.
Fluxo genético
O fluxo gênico é outra importante força evolutiva, envolvendo a troca de material genético entre populações e regiões geográficas. Sem o fluxo gênico, não haveria diversidade — e sem diversidade, uma espécie corre maior risco de extinção. O fluxo gênico pode ser visto no processo de polinização, no qual abelhas ou borboletas transportam e transferem pólen de uma área para outra. Sempre que um gene é introduzido em uma nova população onde ele não existia antes, isso é fluxo gênico.

Especiação
A especiação é o surgimento de uma nova espécie em resposta a uma mudança ou pressão ambiental. A especiação alopátrica, mencionada anteriormente, é a forma mais comum de evento de especiação. Durante a especiação alopátrica, uma espécie diverge quando duas populações se isolam uma da outra e continuam a evoluir. Esse isolamento é criado por barreiras geográficas, como montanhas, rios ou oceanos. Um bom exemplo de especiação alopátrica são as diferentes espécies de esquilo encontradas nos dois lados do Grand Canyon. Descendentes de um ancestral comum, os esquilos se isolaram reprodutivamente uns dos outros pelo Grand Canyon, resultando em espécies diferentes.

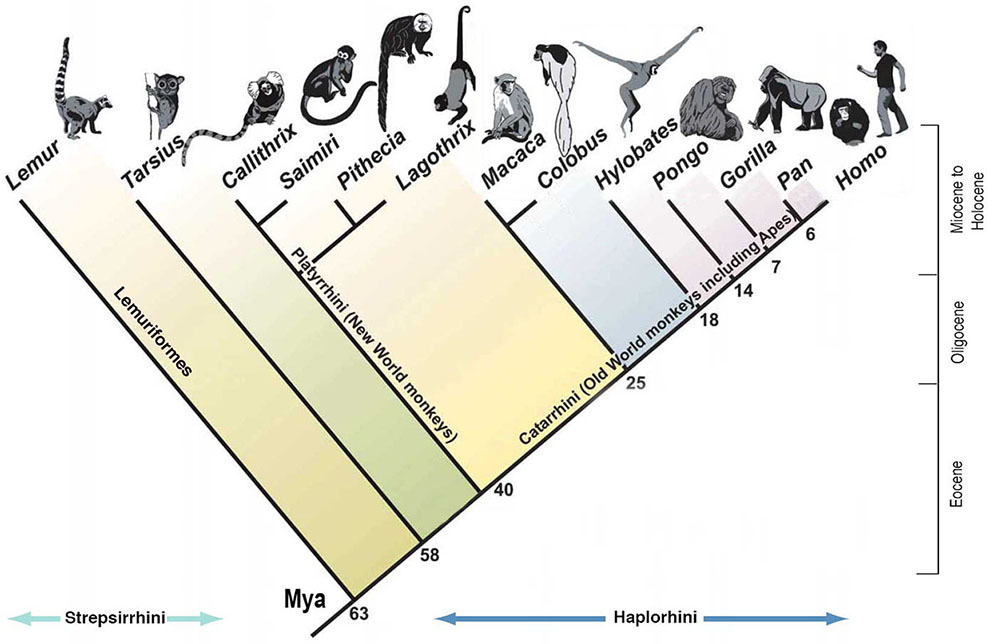
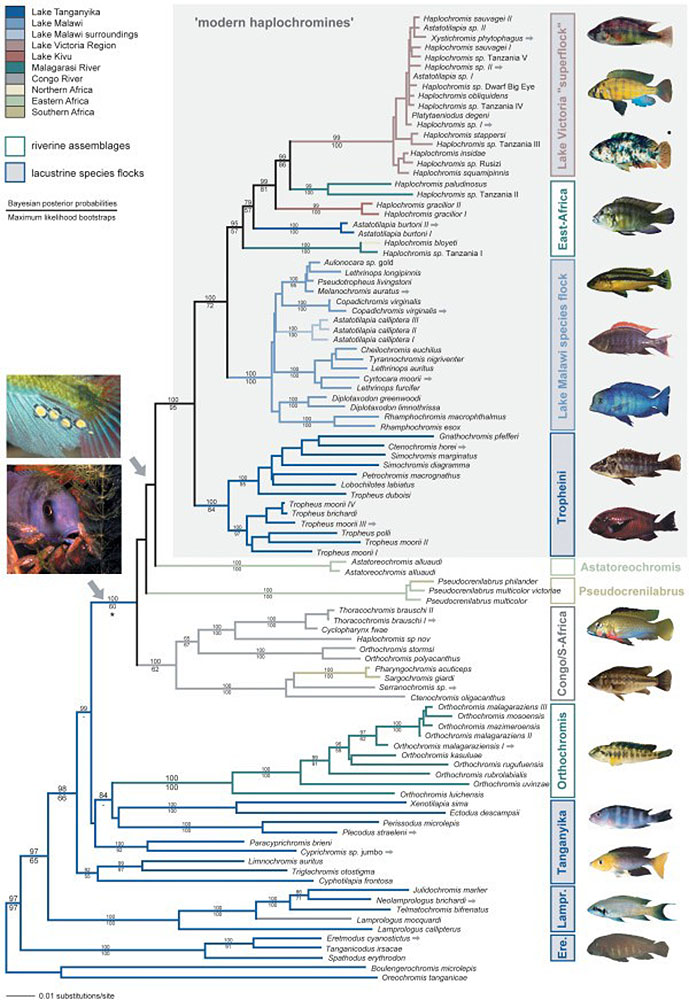
A especiação simpátrica envolve espécies que descendem de um ancestral comum e permanecem em um local sem uma barreira geográfica. Um bom exemplo são os peixes ciclídeos da África Oriental, que experimentam isolamento reprodutivo não devido a uma barreira física, mas à seleção de parceiros com certa coloração pelas fêmeas. A quantidade de luz que atinge diferentes níveis e profundidades do lago afeta a forma como as cores dos machos aparecem para as fêmeas. Os peixes ciclídeos da África Oriental também são um bom exemplo de radiação adaptativa. A radiação adaptativa é vista quando uma ou mais espécies dão origem a muitas novas espécies em um tempo relativamente curto. Pesquisas mostram que uma explosão de cerca de 250 espécies muito diversas de ciclídeos no Lago Tanganica ocorreu em menos de 10 milhões de anos (Takahashi e Koblmüller 2011). Outra pesquisa sugere que o ancestral comum foi o resultado de um enxame híbrido de dois locais diferentes, conforme visto na Figura 4.16. (Meier et al. 2017).
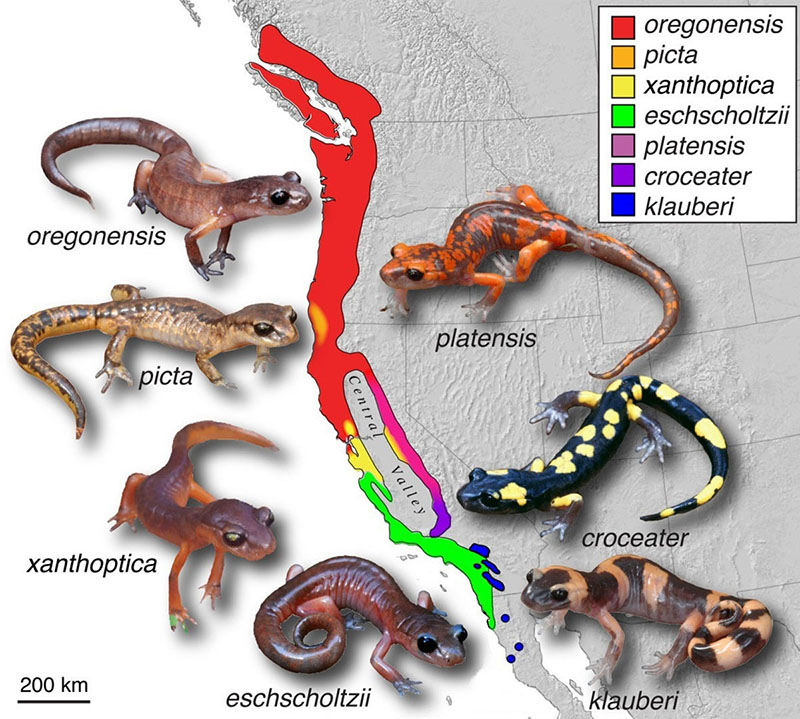
Na especiação peripátrica, membros da mesma população são separados e, com o tempo, evoluem como espécies separadas. A especiação em anel é considerada por alguns como um tipo de especiação peripátrica. A especiação em anéis ocorre quando várias espécies coexistem por um tempo em uma região próxima a uma extremidade de uma barreira geográfica. Quando parte da população migra da população original (ou pool genético) para o outro lado da barreira, ocorre o isolamento reprodutivo. O isolamento reprodutivo é mais forte para a parte da população que está mais distante da população original. Quando ocorre muita variação entre dois grupos, eles não se cruzam mais e, como resultado, a especiação - o desenvolvimento de duas espécies separadas - pode ocorrer. Embora seja bastante rara, acredita-se que a especiação em anéis explique as diferentes espécies do gênero de salamandra californiana Ensatina.
Gradualismo versus evolução pontuada
Os antropólogos biológicos estão interessados não apenas em como uma espécie é melhor definida, mas também em com que frequência e por que meios novas espécies são desenvolvidas. A visão tradicional da evolução pressupõe que as mudanças morfológicas, comportamentais e genéticas ocorrem gradualmente e se acumulam em uma única linha contínua e não ramificada; essa visão da evolução é conhecida como gradualismo. Se essa perspectiva estiver correta, os cientistas esperariam encontrar vários fósseis exibindo evidências de que eles estão lenta e gradualmente fazendo a transição para espécies novas e distintas. No entanto, embora os fósseis sejam raros, fósseis que mostram evidências de formas de transição são ainda mais raros. Embora a escassez de fósseis de transição seja frequentemente atribuída à incompletude do registro fóssil, isso fez com que alguns antropólogos biológicos questionassem se a evolução é realmente gradual.
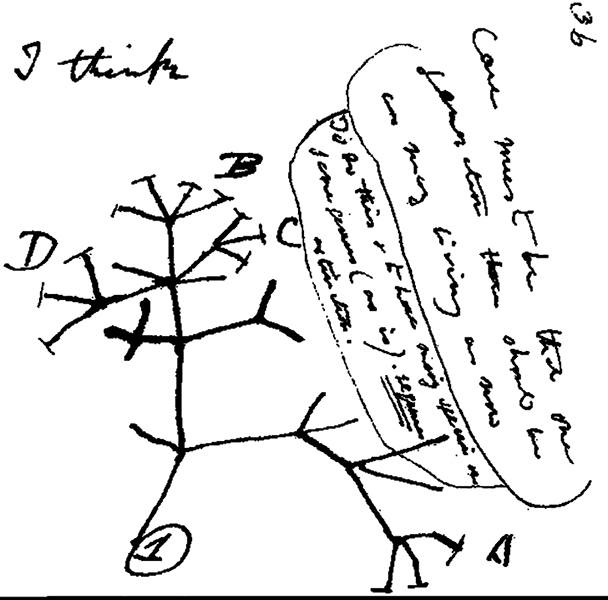
O que pode ser observado no registro fóssil são populações estáticas que são interrompidas por explosões repentinas de mudanças. Esse fenômeno de longos períodos de estase, ou nenhuma mudança, seguido por períodos rápidos de mudança é conhecido como equilíbrio pontuado. Em vez de um acúmulo gradual de pequenas mudanças, o equilíbrio pontuado sugere que mudanças rápidas devido a uma variedade de fatores ambientais, incluindo mudanças climáticas, são características da formação de novas espécies. Os dados fósseis de um grande número de organismos mostram exatamente isso: longos períodos de estase seguidos por mudanças rápidas e massivas. A escassez de formas intermediárias no registro fóssil levou alguns a concluir que o equilíbrio pontuado é a teoria dominante. No entanto, o fato de existirem formas intermediárias sugere que o gradualismo também é um fator importante no processo de evolução. Uma pesquisa descobriu que 30 a 35 por cento dos eventos de especiação ocorreram como resultado de um evento ou mudança repentina, enquanto o restante mostrou evidências de gradualismo (Phillips 2006). Tanto nos modelos graduais quanto nos pontuados, a especiação assume a forma de ramificações ao longo do tempo, em vez de uma progressão linear. A evolução não é linear nem progressiva, mas sim um processo de ramificação — uma árvore da vida contendo áreas de divergência e pontos de uma ancestralidade comum compartilhada.
A Árvore da Vida: Mostrando relacionamentos evolutivos
A resposta, é claro, seria Limenitis archippus, a borboleta vice-rei, que é uma imitação da borboleta-monarca (Danaus plexippus). A primeira parte do nome do vice-rei, Limenitis, é o gênero. O fato de ser diferente dos outros mostra que está mais distante.

É importante observar que o sistema de classificação de Lineu tem limites. Às vezes, as espécies podem ser difíceis de identificar apenas pelas características físicas. Espécies que apresentam formas mímicas e larvais em diferentes estágios de desenvolvimento podem assumir a aparência de outros organismos, resultando em erros na classificação. Você pode dizer qual das borboletas na Figura 4.19 é a monarca? Um exame minucioso revela que as marcas nas asas são um pouco diferentes. O monarca está à esquerda e o monarca mímico, o vice-rei, está à direita. Da mesma forma, na Figura 4.20, você pode ver como pode ser difícil classificar corretamente cracas, caranguejos e lapas com base na aparência física. Pode-se ficar tentado a classificar a craca e a laca como intimamente relacionadas devido às conchas cônicas que elas compartilham, quando, na verdade, a craca está mais intimamente relacionada ao caranguejo, pois ambos são crustáceos. As conchas cônicas da craca e da lapa são adaptações semelhantes em resposta a pressões ambientais semelhantes, não evidências de que estejam intimamente relacionadas ou compartilhem um ancestral comum.

Morfologias estruturais como evidência de relacionamento
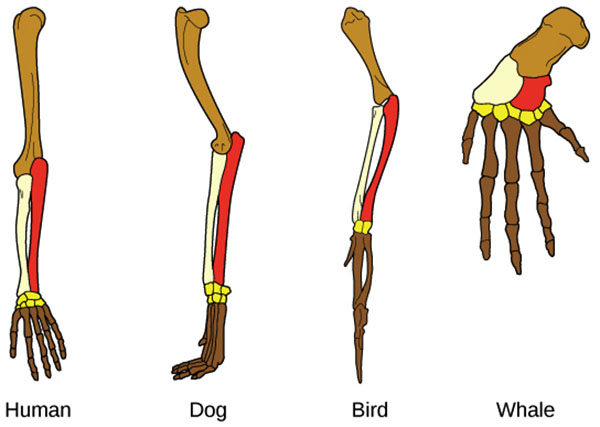
As semelhanças estruturais podem ser características derivadas (estruturas homólogas), herdadas de um ancestral comum ou podem ter se desenvolvido de forma independente (estruturas análogas). Um exemplo de estrutura homóloga é a mão agarrada encontrada em humanos e chimpanzés, o que sugere que humanos e chimpanzés compartilham um ancestral comum que também tinha uma mão agarrada. Estruturas análogas são vistas na asa de uma borboleta e na asa de um morcego. Embora ambas as asas tenham uma função semelhante, esses dois organismos provavelmente desenvolveram suas asas de forma independente e não necessariamente compartilham um ancestral comum. Identificar homologias é essencial para criar hierarquias de relações filogenéticas porque a homologia indica que características compartilhadas são devidas à descendência comum. No entanto, as homologias podem ser difíceis de identificar na natureza e são fáceis de confundir com características análogas.
A cladística, ou o uso de cladogramas, é um método de distinguir visualmente entre características ancestrais homólogas e derivadas. As características ancestrais são encontradas no ancestral comum da espécie que está sendo classificada, enquanto as características derivadas são encontradas apenas nos grupos em questão. Uma característica ancestral que os humanos compartilham com os ancestrais comuns são os polegares opostos. Em contraste, uma característica derivada que só é encontrada em humanos modernos é o queixo. Ao analisar exclusivamente as características derivadas, os antropólogos biológicos podem desenvolver uma compreensão mais clara das relações entre os grupos em estudo.
A Árvore Molecular da Vida e a Filogenética
O surgimento da ciência genética e molecular forneceu ferramentas e linhas de evidência adicionais para verificar as relações evolutivas. A árvore filogenética é um modelo usado pelos taxonomistas modernos para revelar a complexidade e diversidade da vida e seus muitos ramos. As árvores filogenéticas mostram como as espécies e outros grupos de táxons evoluíram a partir de uma série de ancestrais comuns. Eles são baseados em evidências físicas e genéticas.