
11.6 : Comment les procaryotes asexués atteignent la diversité génétique
- Page ID
- 187675

Objectifs d'apprentissage
- Comparez les processus de transformation, de transduction et de conjugaison
- Expliquer comment le transfert de gènes asexué entraîne la diversité génétique des procaryotes
- Expliquer la structure et les conséquences des transposons sur la diversité génétique bactérienne
Généralement, lorsque nous considérons le transfert génétique, nous pensons au transfert vertical de gènes, à la transmission de l'information génétique d'une génération à l'autre. Le transfert vertical de gènes est de loin le principal mode de transmission de l'information génétique dans toutes les cellules. Dans les organismes qui se reproduisent sexuellement, les croisements et l'assortiment indépendant de chromosomes individuels au cours de la méiose contribuent à la diversité génétique de la population. La diversité génétique est également introduite lors de la reproduction sexuée, lorsque les informations génétiques de deux parents, chacun avec des compléments d'informations génétiques différents, sont combinées, produisant de nouvelles combinaisons de génotypes parentaux chez la progéniture diploïde. La survenue de mutations contribue également à la diversité génétique d'une population. La diversité génétique de la progéniture est utile dans des environnements changeants ou incohérents et peut être l'une des raisons du succès évolutif de la reproduction sexuée.
Lorsque les procaryotes et les eucaryotes se reproduisent de manière asexuée, ils transfèrent une copie presque identique de leur matériel génétique à leur progéniture par transfert génique vertical. Bien que la reproduction asexuée produise plus de descendants plus rapidement, tous les avantages de la diversité chez ces enfants sont perdus. Comment alors les organismes dont le mode de reproduction dominant est asexué créent-ils de la diversité génétique ? Chez les procaryotes, le transfert horizontal de gènes (HGT), c'est-à-dire l'introduction de matériel génétique d'un organisme à un autre au sein de la même génération, est un moyen important d'introduire la diversité génétique. La HGT permet même à des espèces apparentées de partager des gènes, influençant ainsi leur phénotype. On pense que le HGT est plus répandu chez les procaryotes, mais que seule une petite fraction du génome procaryote peut être transférée par ce type de transfert à tout moment. Au fur et à mesure que le phénomène est étudié de manière plus approfondie, il peut s'avérer encore plus fréquent. De nombreux scientifiques pensent que la HGT et la mutation sont des sources importantes de variation génétique, matière première du processus de sélection naturelle, chez les procaryotes. Bien que le HGT soit plus fréquent chez les organismes liés à l'évolution, il peut se produire entre deux espèces qui vivent ensemble au sein d'une communauté naturelle.
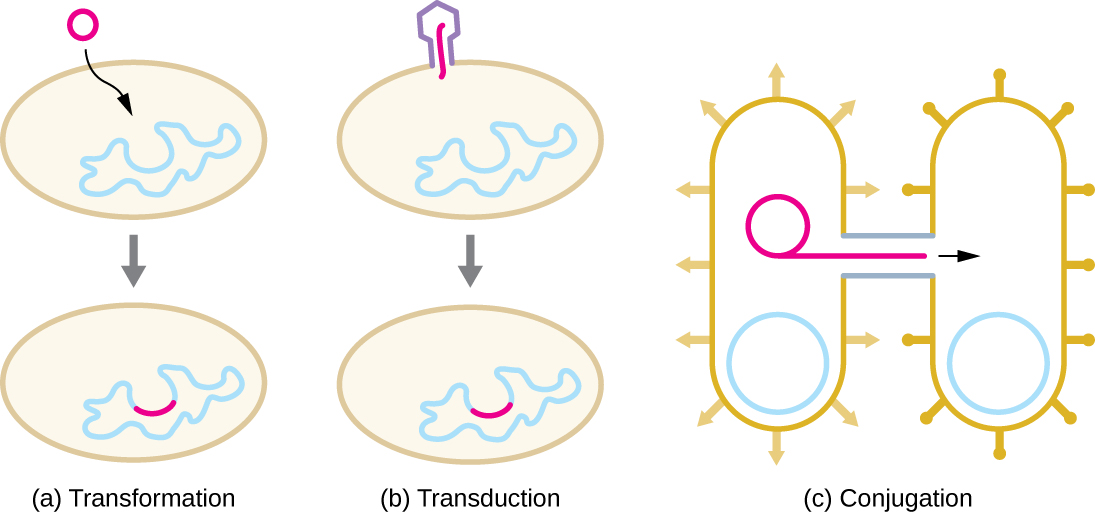
La HGT chez les procaryotes est connue pour se produire par les trois principaux mécanismes illustrés dans la figure\(\PageIndex{1}\) :
- Transformation : l'ADN nu est extrait de l'environnement
- Transduction : les gènes sont transférés entre les cellules d'un virus (voir Le cycle de vie viral)
- Conjugaison : utilisation d'un tube creux appelé pilus de conjugaison pour transférer des gènes entre les cellules
Exercice\(\PageIndex{1}\)
- Quelles sont les trois manières dont la reproduction sexuée introduit une variation génétique chez la progéniture ?
- Quel est l'avantage de la reproduction asexuée ?
- Quels sont les trois mécanismes du transfert horizontal de gènes chez les procaryotes ?
Transformation
Frederick Griffith a été le premier à démontrer le processus de transformation. En 1928, il a montré que des bactéries vivantes et non pathogènes de Streptococcus pneumoniae pouvaient être transformées en bactéries pathogènes par exposition à une souche pathogène tuée par la chaleur. Il a conclu qu'une sorte d'agent, qu'il a appelé le « principe de transformation », avait été transmise des bactéries pathogènes mortes aux bactéries vivantes non pathogènes. En 1944, Oswald Avery (1877-1955), Colin MacLeod (1909—1972) et Maclyn McCarty (1911-2005) ont démontré que le principe de transformation était l'ADN (voir Utiliser la microbiologie pour découvrir les secrets de la vie).
Lors de sa transformation, le procaryote absorbe de l'ADN nu présent dans son environnement et dérivé d'autres cellules qui se sont lysées à leur mort et ont libéré leur contenu, y compris leur génome, dans l'environnement. De nombreuses bactéries sont naturellement compétentes, c'est-à-dire qu'elles se lient activement à l'ADN environnemental, le transportent à travers leurs enveloppes cellulaires jusqu'à leur cytoplasme et le rendent monocaténaire. En général, l'ADN étranger bicaténaire contenu dans les cellules est détruit par les nucléases pour se défendre contre les infections virales. Cependant, ces nucléases sont généralement inefficaces contre l'ADN monocaténaire, de sorte que cet ADN monocaténaire de la cellule a la possibilité de se recombiner dans le génome bactérien. Une molécule d'ADN qui contient des fragments d'ADN provenant de différents organismes est appelée ADN recombinant. (L'ADN recombinant sera discuté plus en détail dans Microbes et outils du génie génétique.) Si la bactérie incorpore le nouvel ADN dans son propre génome par recombinaison, la cellule bactérienne peut acquérir de nouvelles propriétés phénotypiques. Par exemple, si une bactérie non pathogène absorbe l'ADN d'un gène de toxine provenant d'un agent pathogène et l'incorpore ensuite dans son chromosome, elle peut également devenir pathogène. L'ADN plasmidique peut également être absorbé par des bactéries compétentes et conférer de nouvelles propriétés à la cellule. Dans l'ensemble, la transformation dans la nature est un processus relativement inefficace car les niveaux d'ADN dans l'environnement sont faibles en raison de l'activité des nucléases qui sont également libérées lors de la lyse cellulaire. De plus, la recombinaison génétique est inefficace pour intégrer de nouvelles séquences d'ADN dans le génome.
Dans la nature, la transformation bactérienne est un mécanisme important pour l'acquisition d'éléments génétiques codant pour les facteurs de virulence et la résistance aux antibiotiques. Il a été démontré que les gènes codant pour la résistance aux composés antimicrobiens sont très répandus dans la nature, même dans des environnements non influencés par l'homme. Ces gènes, qui permettent aux microbes vivant dans des communautés mixtes de se disputer des ressources limitées, peuvent être transférés au sein d'une population par transformation, ainsi que par les autres processus de la HGT. En laboratoire, nous pouvons exploiter le processus naturel de transformation bactérienne pour le génie génétique afin de fabriquer une grande variété de médicaments, comme indiqué dans Microbes et outils du génie génétique.
Exercice\(\PageIndex{2}\)
Pourquoi une cellule bactérienne produit-elle de l'ADN environnemental introduit dans la cellule sous une forme monocaténaire ?
Transduction
Les virus qui infectent les bactéries (bactériophages) peuvent également déplacer de petits fragments d'ADN chromosomique d'une bactérie à l'autre dans le cadre d'un processus appelé transduction (voir Figure 6.2.3). Rappelons que lors de la transduction généralisée, tout fragment d'ADN chromosomique peut être transféré vers une nouvelle cellule hôte par empaquetage accidentel de l'ADN chromosomique dans une tête de phage lors de l'assemblage du phage. En revanche, la transduction spécialisée résulte de l'excision imprécise d'un prophage lysogénique du chromosome bactérien, de telle sorte qu'il transporte un morceau du chromosome bactérien de chaque côté du site d'intégration du phage vers une nouvelle cellule hôte. Par conséquent, l'hôte peut acquérir de nouvelles propriétés. Ce processus s'appelle la conversion lysogénique. D'un point de vue médical, un phage lysogénique peut être porteur d'un gène de virulence pour son nouvel hôte. Une fois inséré dans le chromosome du nouvel hôte, celui-ci peut acquérir de la pathogénicité. Plusieurs bactéries pathogènes, dont Corynebacterium diphtheriae (l'agent causal de la diphtérie) et Clostridium botulinum (l'agent causal du botulisme), sont virulentes en raison de l'introduction de gènes codant pour les toxines par des bactériophages lysogéniques, ce qui confirme la pertinence de la transduction dans l'échange de gènes impliqués dans les maladies infectieuses. Les archées possèdent leurs propres virus qui transloquent le matériel génétique d'un individu à un autre.
Exercice\(\PageIndex{3}\)
- Quel est l'agent de transduction des cellules procaryotes ?
- Dans la transduction spécialisée, d'où vient le fragment d'ADN transducteur ?
Les conséquences cliniques de la transduction
Paul, un travailleur humanitaire d'Atlanta âgé de 23 ans, s'est rendu en Haïti en 2011 pour apporter de l'aide à la suite du tremblement de terre de 2010. Après y avoir travaillé pendant plusieurs semaines, il a soudainement commencé à ressentir des troubles abdominaux, notamment de fortes crampes, des nausées, des vomissements et une diarrhée aqueuse. Il a également commencé à ressentir d'intenses crampes musculaires. Dans une clinique locale, le médecin soupçonnait que les symptômes de Paul étaient dus au choléra, car il y avait eu une épidémie de choléra après le tremblement de terre. Comme le choléra se transmet par voie fécale-orale, les défaillances des infrastructures sanitaires, comme celles qui se produisent souvent à la suite de catastrophes naturelles, peuvent provoquer des épidémies. Le médecin a confirmé le diagnostic présomptif à l'aide d'un test sur bandelette réactive pour le choléra. Il a ensuite prescrit à Paul une dose unique de doxycycline, ainsi que des sels de réhydratation orale, lui demandant de boire des quantités importantes d'eau propre.
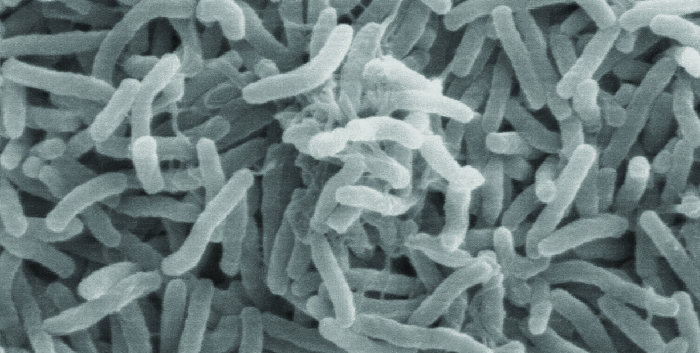
Le choléra est causé par la tige incurvée gram-négative Vibrio cholerae (Figure\(\PageIndex{2}\)). Ses symptômes résultent en grande partie de la production de la toxine cholérique (CT), qui active finalement un transporteur de chlorure pour pomper les ions chlorure des cellules épithéliales vers la lumière intestinale. L'eau suit ensuite les ions chlorure, provoquant la diarrhée aqueuse prolifique caractéristique du choléra. Le gène codant pour la toxine cholérique est incorporé dans le chromosome bactérien de V. choléra par infection de la bactérie par le phage filamenteux CTX lysogène, qui porte le gène CT et l'introduit dans le chromosome lors de l'intégration du prophage. Ainsi, les souches pathogènes de V. cholerae résultent d'un transfert horizontal de gènes par transduction spécialisée.
Exercice\(\PageIndex{4}\)
- Pourquoi les épidémies de choléra sont-elles plus fréquentes à la suite d'une catastrophe naturelle ?
- Pourquoi les crampes musculaires sont-elles un symptôme courant du choléra ? Pourquoi le traitement aux sels de réhydratation orale est-il si important pour le traitement du choléra ?
- Dans les zones touchées par le choléra, quelles stratégies les gens pourraient-ils utiliser pour prévenir la transmission de la maladie ?
Conjugaison
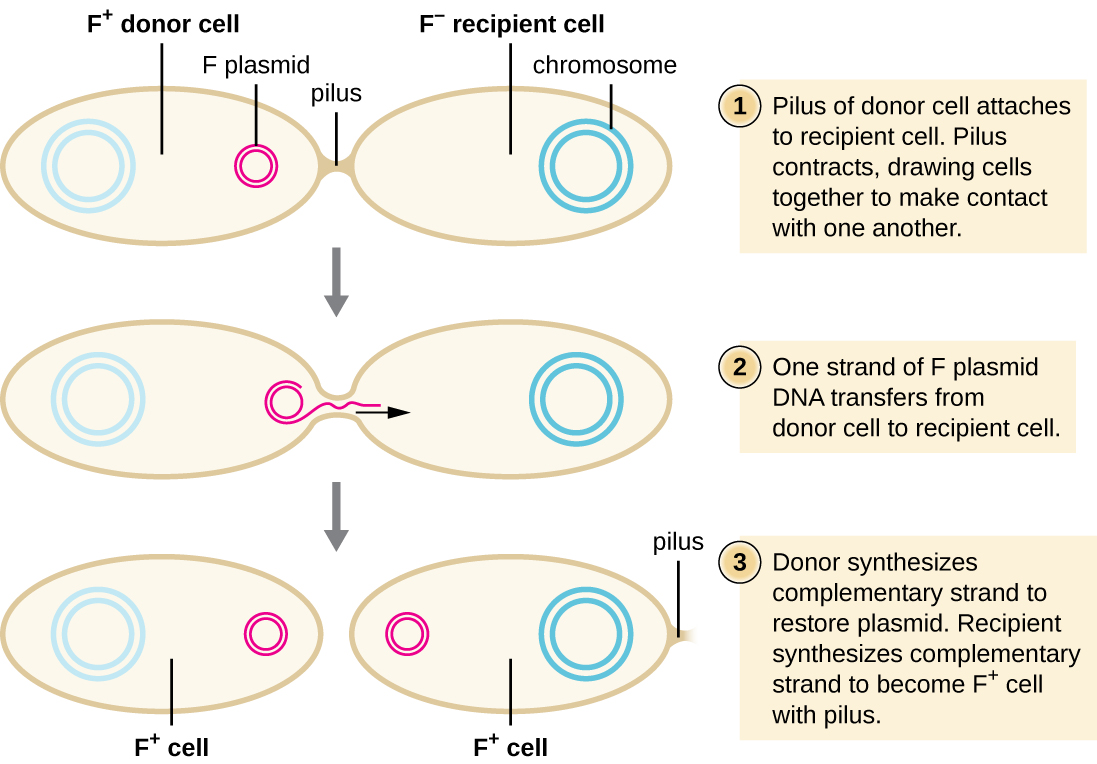
Lors de la conjugaison, l'ADN est directement transféré d'un procaryote à un autre au moyen d'un pilus de conjugaison, qui met les organismes en contact les uns avec les autres. Chez E. coli, les gènes codant pour la capacité de conjugaison sont situés sur un plasmide bactérien appelé plasmide F, également connu sous le nom de facteur de fertilité, et le pilus de conjugaison est appelé pilus F. Les gènes du plasmide F codent à la fois les protéines composant le pilus F et celles impliquées dans la réplication circulaire du plasmide. Les cellules contenant le plasmide F, capable de former un pilus F, sont appelées cellules F+ ou cellules donneuses, et celles dépourvues de plasmide F sont appelées cellules F − ou cellules réceptrices.
Conjugaison du plasmide F
Lors de la conjugaison typique chez E. coli, le pilus F d'un lymphocyte F+ entre en contact avec un lymphocyte F et se rétracte, mettant ainsi en contact les deux enveloppes cellulaires (Figure\(\PageIndex{3}\)). Ensuite, un pont cytoplasmique se forme entre les deux cellules au site du pilus de conjugaison. Lorsque la réplication circulaire du plasmide F se produit dans la cellule F+, une copie monocaténaire du plasmide F est transférée par le pont cytoplasmique à la cellule F −, qui synthétise ensuite le brin complémentaire, le rendant bicaténaire. La cellule F − devient maintenant une cellule F+ capable de fabriquer son propre pilus de conjugaison. Finalement, dans une population bactérienne mixte contenant à la fois des cellules F+ et des cellules F −, toutes les cellules deviendront des cellules F+. Les gènes du plasmide E. coli F codent également pour des protéines empêchant la conjugaison entre les cellules F+.
Conjugaison des cellules F' et Hfr
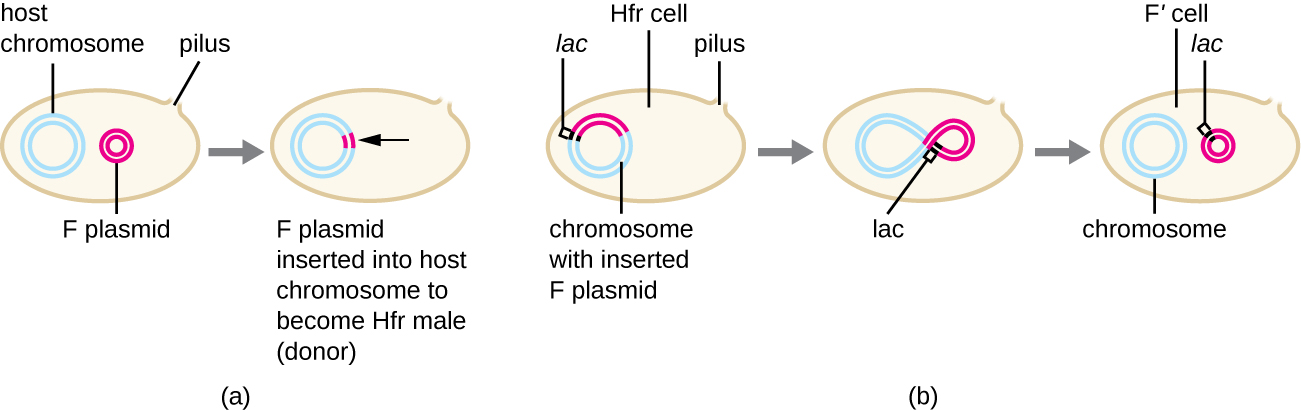
Bien que la conjugaison typique chez E. coli entraîne le transfert de l'ADN du plasmide F uniquement, la conjugaison peut également transférer l'ADN chromosomique. En effet, le plasmide F s'intègre occasionnellement dans le chromosome bactérien par recombinaison entre le plasmide et le chromosome, formant une cellule Hfr (Figure\(\PageIndex{4}\)). « Hfr » fait référence à la fréquence élevée de recombinaison observée lorsque les cellules F − réceptrices reçoivent des informations génétiques des cellules Hfr par conjugaison. À l'instar de l'excision imprécise d'un prophage lors d'une transduction spécialisée, le plasmide F intégré peut également être excisé de manière imprécise du chromosome, produisant un plasmide F' qui transporte avec lui de l'ADN chromosomique adjacent au site d'intégration. Lors de la conjugaison, cet ADN est introduit dans la cellule réceptrice et peut être soit conservé dans le plasmide F', soit recombiné dans le chromosome bactérien de la cellule réceptrice.
Les cellules Hfr peuvent également traiter le chromosome bactérien comme un énorme plasmide F et tenter d'en transférer une copie à une cellule F − réceptrice. En raison de la taille du chromosome bactérien, le transfert de l'ensemble du chromosome prend beaucoup de temps (Figure\(\PageIndex{5}\)). Cependant, le contact entre les cellules bactériennes pendant la conjugaison est transitoire, de sorte qu'il est rare que l'ensemble du chromosome soit transféré. L'ADN chromosomique de l'hôte situé à proximité du site d'intégration du plasmide F, déplacé par le processus unidirectionnel de réplication circulaire, est plus susceptible d'être transféré et recombiné dans le chromosome de la cellule réceptrice que les gènes hôtes plus éloignés. Ainsi, la localisation relative des gènes bactériens sur le génome de la cellule Hfr peut être cartographiée en fonction du moment où ils sont transférés par conjugaison. Par conséquent, avant l'ère du séquençage généralisé du génome bactérien, les distances sur les cartes du génome procaryote étaient souvent mesurées en quelques minutes.
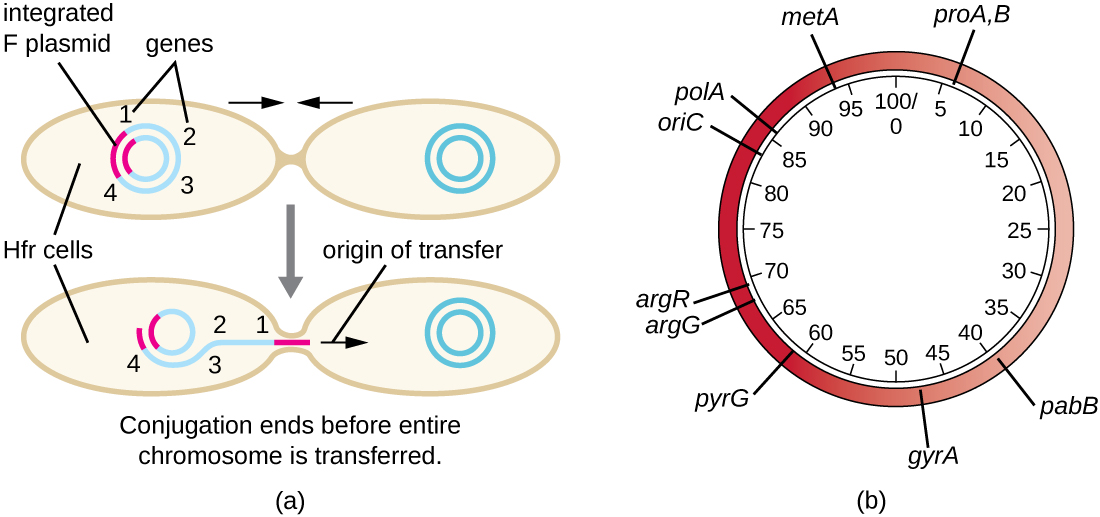
Conséquences et applications de la conjugaison
Les plasmides sont un type important d'élément extrachromosomique de l'ADN chez les bactéries et, dans les cellules qui les hébergent, ils sont considérés comme faisant partie du génome bactérien. D'un point de vue clinique, les plasmides codent souvent pour des gènes impliqués dans la virulence. Par exemple, les gènes codant pour des protéines qui rendent une cellule bactérienne résistante à un antibiotique particulier sont codés sur des plasmides R. Les plasmides R, en plus de leurs gènes de résistance aux antimicrobiens, contiennent des gènes qui contrôlent la conjugaison et le transfert du plasmide. Les plasmides R sont capables de se transférer entre des cellules d'une même espèce et entre des cellules d'espèces différentes. Les plasmides R simples contiennent généralement plusieurs gènes conférant une résistance à de multiples antibiotiques.
Les gènes nécessaires à la production de diverses toxines et molécules importantes pour la colonisation au cours de l'infection peuvent également être trouvés codés sur des plasmides. Par exemple, les souches d'E. coli productrices de vérotoxine (VTEC) semblent avoir acquis les gènes codant pour la toxine Shiga à partir de son parent Gram négatif Shigella dysenteriae grâce à l'acquisition d'un grand plasmide codant pour cette toxine. Le VTEC provoque une maladie diarrhéique sévère qui peut entraîner un syndrome hémolytique et urémique (SHU), qui peut entraîner une insuffisance rénale et la mort.
En milieu non clinique, les gènes bactériens codant pour les enzymes métaboliques nécessaires à la dégradation de composés atypiques spécialisés tels que les hydrocarbures aromatiques polycycliques (HAP) sont également fréquemment codés sur des plasmides. De plus, certains plasmides ont la capacité de passer des cellules bactériennes à d'autres types de cellules, comme celles des plantes et des animaux, par des mécanismes distincts de la conjugaison. Ces mécanismes et leur utilisation en génie génétique sont abordés dans la section Applications modernes de la génétique microbienne.
Exercice\(\PageIndex{5}\)
- Quel type de réplication se produit lors de la conjugaison ?
- Que se passe-t-il pour produire une cellule Hfr E. coli ?
- Quels types de caractères sont codés sur les plasmides ?
Transposition
Les éléments génétiques appelés transposons (éléments transposables), ou « gènes sauteurs », sont des molécules d'ADN qui comportent des séquences répétées inversées spéciales à leurs extrémités et un gène codant pour l'enzyme transposase (Figure\(\PageIndex{6}\)). Les transposons permettent à l'ensemble de la séquence d'être extraite indépendamment d'un emplacement dans une molécule d'ADN et de s'intégrer à l'ADN ailleurs grâce à un processus appelé transposition. Les transposons ont été découverts à l'origine dans le maïs par la généticienne américaine Barbara McClintock (1902-1992) dans les années 1940. Des transposons ont depuis été découverts dans tous les types d'organismes, à la fois des procaryotes et des eucaryotes. Ainsi, contrairement aux trois mécanismes précédemment discutés, la transposition n'est pas spécifique aux procaryotes. La plupart des transposons ne sont pas réplicatifs, c'est-à-dire qu'ils se déplacent de manière « copier-coller ». Certains peuvent toutefois être réplicatifs et conserver leur emplacement dans l'ADN tout en faisant une copie à insérer ailleurs (« copier-coller »). Comme les transposons peuvent se déplacer au sein d'une molécule d'ADN, d'une molécule d'ADN à une autre, voire d'une cellule à l'autre, ils ont la capacité d'introduire de la diversité génétique. Le mouvement au sein d'une même molécule d'ADN peut modifier le phénotype en inactivant ou en activant un gène.
Les transposons peuvent être porteurs de gènes supplémentaires, ce qui permet de déplacer ces gènes d'un endroit à un autre avec eux. Par exemple, les transposons bactériens peuvent déplacer les gènes de résistance aux antibiotiques, les déplaçant des chromosomes vers les plasmides. Il a été démontré que ce mécanisme est responsable de la colocalisation de plusieurs gènes de résistance aux antibiotiques sur un seul plasmide R chez des souches de Shigella provoquant une dysenterie bactérienne. Un tel plasmide R peut ensuite être facilement transféré au sein d'une population bactérienne par le biais du processus de conjugaison.
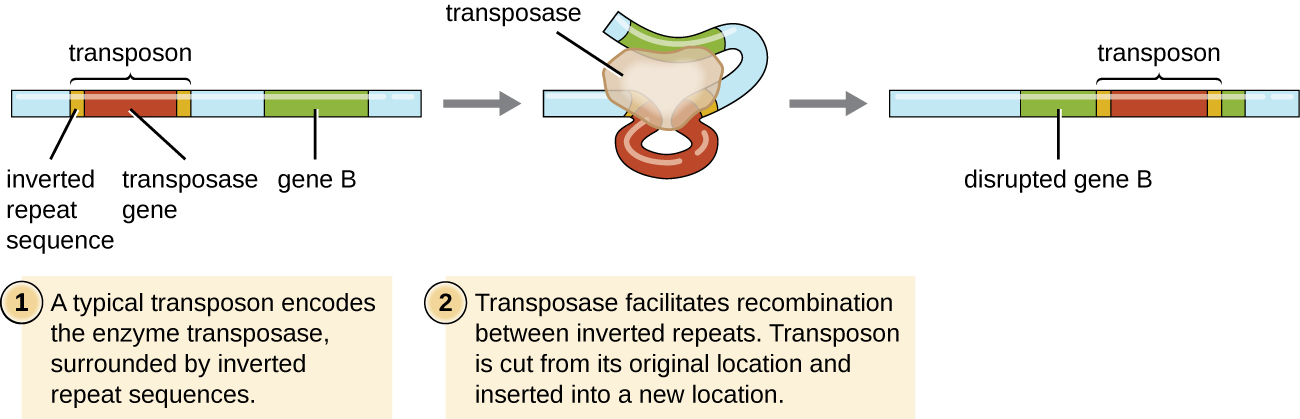
Exercice\(\PageIndex{6}\)
De quelles manières un transposon peut-il affecter le phénotype d'une cellule vers laquelle il se déplace ?
| Terme | Définition |
|---|---|
| Conjugaison | Transfert d'ADN par contact direct à l'aide d'un pilus de conjugaison |
| Transduction | Mécanisme de transfert horizontal de gènes chez les bactéries dans lequel les gènes sont transférés par le biais d'une infection virale |
| Transformation | Mécanisme de transfert horizontal de gènes dans lequel l'ADN environnemental nu est absorbé par une cellule bactérienne |
| Transposition | Processus par lequel l'ADN est extrait indépendamment d'un emplacement dans une molécule d'ADN et s'intègre ailleurs |
Orientation clinique : 3e partie
Malgré la poursuite du traitement antibiotique, l'infection de Mark a continué de progresser rapidement. La région infectée a continué de s'étendre et il a dû être placé sous respirateur pour l'aider à respirer. Le médecin de Mark a ordonné l'ablation chirurgicale du tissu infecté. Après une première intervention chirurgicale, la blessure de Mark a été surveillée quotidiennement pour s'assurer que l'infection ne réapparaissait pas, mais qu'elle continuait de se propager.
Après deux autres cycles de chirurgie, l'infection a finalement semblé être maîtrisée. Quelques jours plus tard, Mark a été retiré du ventilateur et a pu respirer seul. Cependant, il avait perdu beaucoup de peau et de tissus mous sur la partie inférieure de sa jambe.
Exercice\(\PageIndex{7}\)
- Pourquoi l'ablation des tissus infectés permet-elle d'enrayer l'infection ?
- Quelles sont les complications probables de cette méthode de traitement ?
Concepts clés et résumé
- Le transfert horizontal de gènes est un moyen important pour les organismes se reproduisant de manière asexuée, tels que les procaryotes, d'acquérir de nouvelles caractéristiques.
- Il existe trois mécanismes de transfert horizontal de gènes généralement utilisés par les bactéries : la transformation, la transduction et la conjugaison.
- La transformation permet aux cellules compétentes d'absorber l'ADN nu, libéré par d'autres cellules à leur mort, dans leur cytoplasme, où il peut se recombiner avec le génome de l'hôte.
- Lors de la transduction généralisée, n'importe quel fragment d'ADN chromosomique peut être transféré par empaquetage accidentel du chromosome hôte dégradé dans une tête de phage. Lors d'une transduction spécialisée, seul l'ADN chromosomique adjacent au site d'intégration d'un phage lysogène peut être transféré à la suite d'une excision imprécise du prophage.
- La conjugaison est médiée par le plasmide F, qui code un pilus de conjugaison qui met en contact une cellule F+ contenant un plasmide F avec une cellule F -.
- L'intégration rare du plasmide F dans le chromosome bactérien, générant une cellule Hfr, permet le transfert de l'ADN chromosomique du donneur au receveur. De plus, l'excision imprécise du plasmide F du chromosome peut générer un plasmide F' qui peut être transféré à un receveur par conjugaison.
- Le transfert par conjugaison des plasmides R est un mécanisme important pour la propagation de la résistance aux antibiotiques dans les communautés bactériennes.
- Les transposons sont des molécules d'ADN comportant des répétitions inversées à leurs extrémités qui codent également pour l'enzyme transposase, ce qui permet leur déplacement d'un endroit de l'ADN à un autre. Bien qu'ils soient présents à la fois chez les procaryotes et les eucaryotes, les transposons sont cliniquement pertinents chez les bactéries pathogènes pour le mouvement des facteurs de virulence, y compris les gènes de résistance aux antibiotiques.