
8.2 : Catabolisme des glucides
- Page ID
- 187731

Objectifs d'apprentissage
- Décrire pourquoi la glycolyse ne dépend pas de l'oxygène
- Définir et décrire le rendement net des molécules à trois carbones, de l'ATP et du NADH résultant de la glycolyse
- Expliquez comment les molécules de pyruvate à trois carbones sont converties en groupes acétyle à deux carbones qui peuvent être canalisés dans le cycle de Krebs.
- Définir et décrire le rendement net en CO 2, GTP/ATP, FADH 2 et NADH à partir du cycle de Krebs
- Expliquer comment les molécules de carbone intermédiaires du cycle de Krebs peuvent être utilisées dans une cellule
De nombreuses voies enzymatiques existent pour décomposer les glucides afin de capter l'énergie dans les liaisons ATP. De plus, de nombreuses voies cataboliques produisent des molécules intermédiaires qui sont également utilisées comme éléments constitutifs de l'anabolisme. Il est important de comprendre ces processus pour plusieurs raisons. Tout d'abord, étant donné que les principaux processus métaboliques impliqués sont communs à un large éventail d'organismes chimiohétérotrophes, nous pouvons en apprendre beaucoup sur le métabolisme humain en étudiant le métabolisme de bactéries plus faciles à manipuler comme E. coli. Deuxièmement, étant donné que les agents pathogènes animaux et humains sont également des chimiohétérotrophes, il est utile de connaître les détails du métabolisme de ces bactéries, y compris les différences possibles entre les voies bactériennes et humaines, pour le diagnostic des agents pathogènes ainsi que pour la découverte de thérapies antimicrobiennes ciblant agents pathogènes spécifiques. Enfin, l'apprentissage spécifique des voies impliquées dans le métabolisme chimiohétérotrophe permet également de comparer d'autres stratégies métaboliques plus inhabituelles utilisées par les microbes. Bien que la source chimique des électrons initiant le transfert d'électrons soit différente entre les chimiohétérotrophes et les chimioautotrophes, de nombreux procédés similaires sont utilisés dans les deux types d'organismes.
L'exemple type utilisé pour présenter les concepts du métabolisme aux étudiants est le catabolisme des glucides. Pour les chimiohétérotrophes, nos exemples de métabolisme commencent par le catabolisme de polysaccharides tels que le glycogène, l'amidon ou la cellulose. Des enzymes telles que l'amylase, qui décompose le glycogène ou l'amidon, et les cellulases, qui dégradent la cellulose, peuvent provoquer l'hydrolyse des liaisons glycosidiques entre les monomères de glucose de ces polymères, libérant du glucose pour un catabolisme ultérieur.
Glycolyse
Pour les bactéries, les eucaryotes et la plupart des archées, la glycolyse est la voie la plus courante du catabolisme du glucose ; elle produit de l'énergie, réduit les porteurs d'électrons et les molécules précurseurs du métabolisme cellulaire. Chaque organisme vivant effectue une forme de glycolyse, ce qui suggère que ce mécanisme est un ancien processus métabolique universel. Le processus lui-même n'utilise pas d'oxygène ; toutefois, la glycolyse peut être associée à d'autres processus métaboliques aérobies ou anaérobies. La glycolyse a lieu dans le cytoplasme des cellules procaryotes et eucaryotes. Il commence par une seule molécule de glucose à six carbones et se termine par deux molécules d'un sucre à trois carbones appelé pyruvate. Le pyruvate peut être dégradé davantage après la glycolyse pour capter plus d'énergie par la respiration aérobie ou anaérobie, mais de nombreux organismes, y compris de nombreux microbes, peuvent être incapables de respirer ; pour ces organismes, la glycolyse peut être leur seule source de production d'ATP.
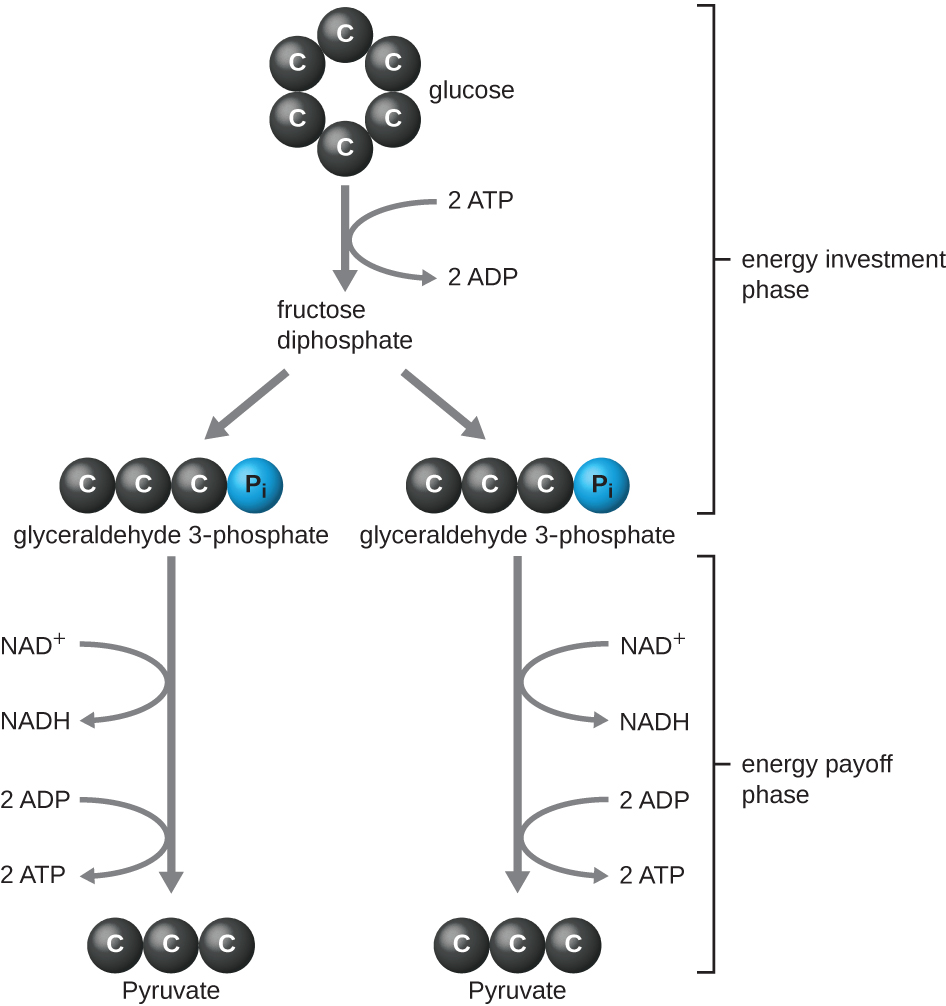
Le type de glycolyse observé chez les animaux et le plus fréquent chez les microbes est la voie Embden-Meyerhof-Parnas (EMP), nommée d'après Gustav Embden (1874-1933), Otto Meyerhof (1884-1951) et Jakub Parnas (1884-1949). La glycolyse utilisant la voie EMP comprend deux phases distinctes (Figure\(\PageIndex{1}\)). La première partie de la voie, appelée phase d'investissement énergétique, utilise l'énergie de deux molécules d'ATP pour modifier une molécule de glucose afin que la molécule de sucre à six carbones puisse être divisée uniformément en deux molécules à trois carbones phosphorylées appelées glycéraldéhyde 3-phosphate (G3P). La deuxième partie de la voie, appelée phase de récupération d'énergie, extrait de l'énergie en oxydant le G3P en pyruvate, en produisant quatre molécules d'ATP et en réduisant deux molécules de NAD+ en deux molécules de NADH, à l'aide d'électrons provenant du glucose. (Une discussion et une illustration de la voie EMP complète avec les structures chimiques et les noms des enzymes figurent à l'annexe C.)
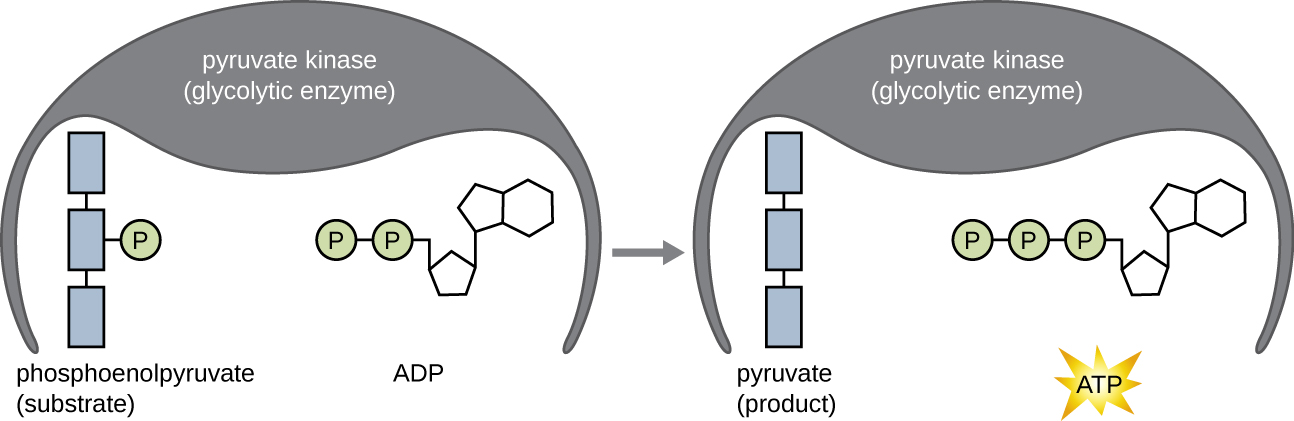
Les molécules d'ATP produites pendant la phase de récupération d'énergie de la glycolyse sont formées par phosphorylation au niveau du substrat (Figure\(\PageIndex{1}\)), l'un des deux mécanismes de production d'ATP. Lors de la phosphorylation au niveau du substrat, un groupe phosphate est retiré d'une molécule organique et est directement transféré vers une molécule d'ADP disponible, produisant de l'ATP. Au cours de la glycolyse, des groupes phosphates à haute énergie provenant des molécules intermédiaires sont ajoutés à l'ADP pour fabriquer de l'ATP.
Dans l'ensemble, dans ce processus de glycolyse, le gain net résultant de la dégradation d'une seule molécule de glucose est le suivant :
- deux molécules d'ATP
- deux molécules de NADH, et
- deux molécules de pyruvate.
Autres voies glycolytiques
Lorsque nous parlons de glycolyse, sauf indication contraire, nous faisons référence à la voie EMP utilisée par les animaux et de nombreuses bactéries. Cependant, certains procaryotes utilisent des voies glycolytiques alternatives. Une alternative importante est le sentier Entner-Doudoroff (ED), nommé d'après ses découvreurs Nathan Entner et Michael Doudoroff (1911-1975). Bien que certaines bactéries, y compris le pathogène opportuniste à Gram négatif Pseudomonas aeruginosa, ne contiennent que la voie de la dysfonction érectile pour la glycolyse, d'autres bactéries, comme E. coli, peuvent utiliser soit la voie ED soit la voie EMP.
Un troisième type de voie glycolytique qui se produit dans toutes les cellules, et qui est très différent des deux voies précédentes, est la voie du pentose phosphate (PPP), également appelée voie du phosphogluconate ou shunt de l'hexose monophosphate. Les preuves suggèrent que le PPP est peut-être la plus ancienne voie glycolytique universelle. Les intermédiaires du PPP sont utilisés pour la biosynthèse des nucléotides et des acides aminés. Par conséquent, cette voie glycolytique peut être favorisée lorsque la cellule a besoin d'une synthèse d'acides nucléiques et/ou de protéines, respectivement. Une discussion et une illustration de l'ensemble de la voie de la dysfonction érectile et du PPP avec les structures chimiques et les noms des enzymes figurent à
Exercice\(\PageIndex{1}\)
Quand un organisme peut-il utiliser la voie ED ou le PPP pour la glycolyse ?
Réaction de transition, coenzyme A et cycle de Krebs
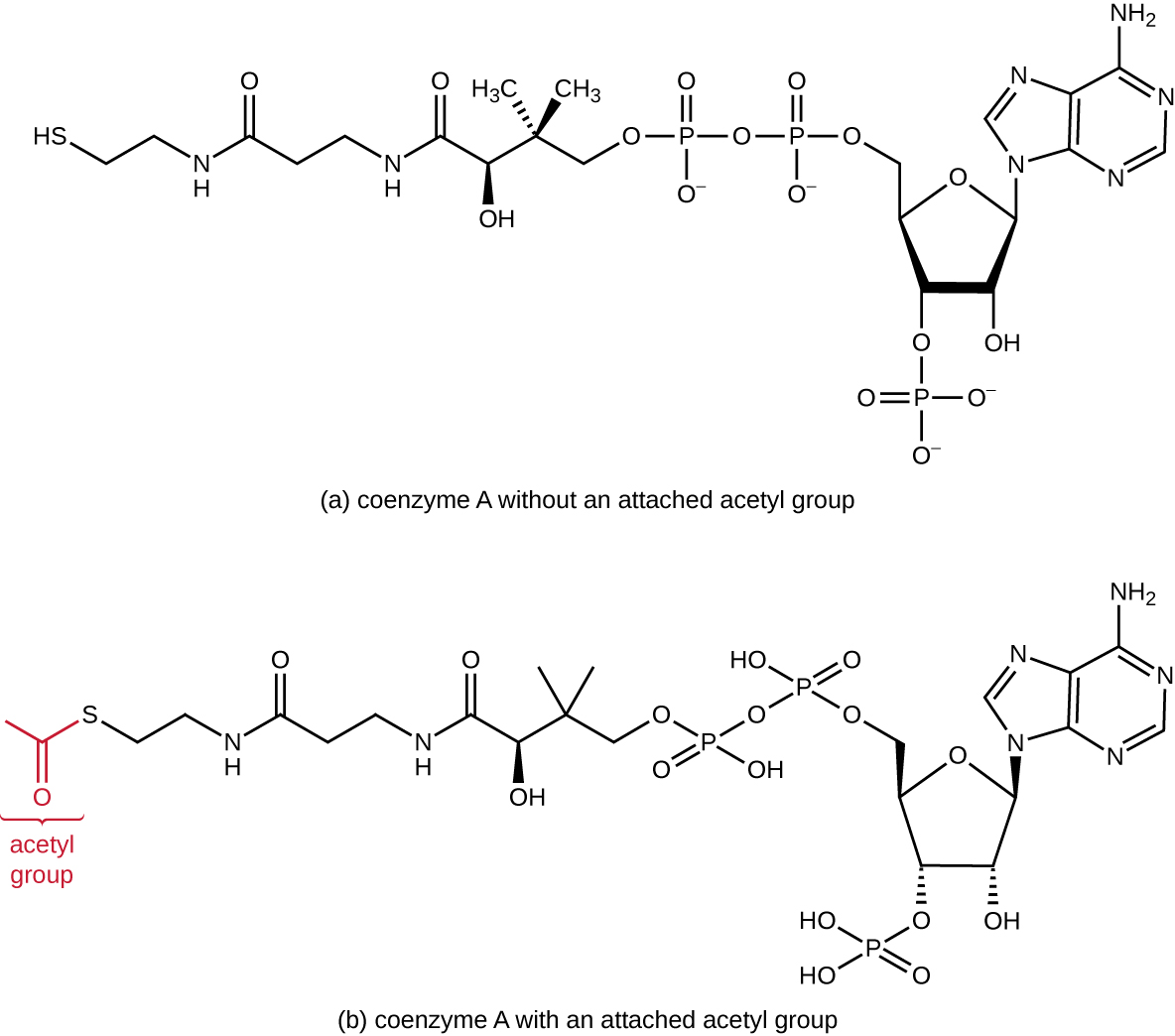
La glycolyse produit du pyruvate, qui peut être oxydé davantage pour capter plus d'énergie. Pour que le pyruvate passe par la voie oxydative suivante, il doit d'abord être décarboxylé par le complexe enzymatique pyruvate déshydrogénase en un groupe acétyle à deux carbones lors de la réaction de transition, également appelée réaction de pont (voir l'annexe C et la figure\(\PageIndex{3}\)). Lors de la réaction de transition, les électrons sont également transférés au NAD+ pour former du NADH. Pour passer à la phase suivante de ce processus métabolique, l'acétyle à deux carbones, relativement petit, doit être attaché à un très gros composé porteur appelé coenzyme A (CoA). La réaction de transition se produit dans la matrice mitochondriale des eucaryotes ; chez les procaryotes, elle se produit dans le cytoplasme parce que les procaryotes sont dépourvus d'organites enfermés dans la membrane.
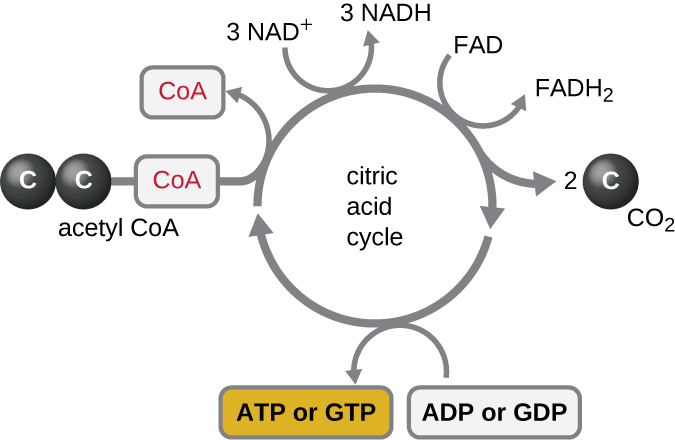
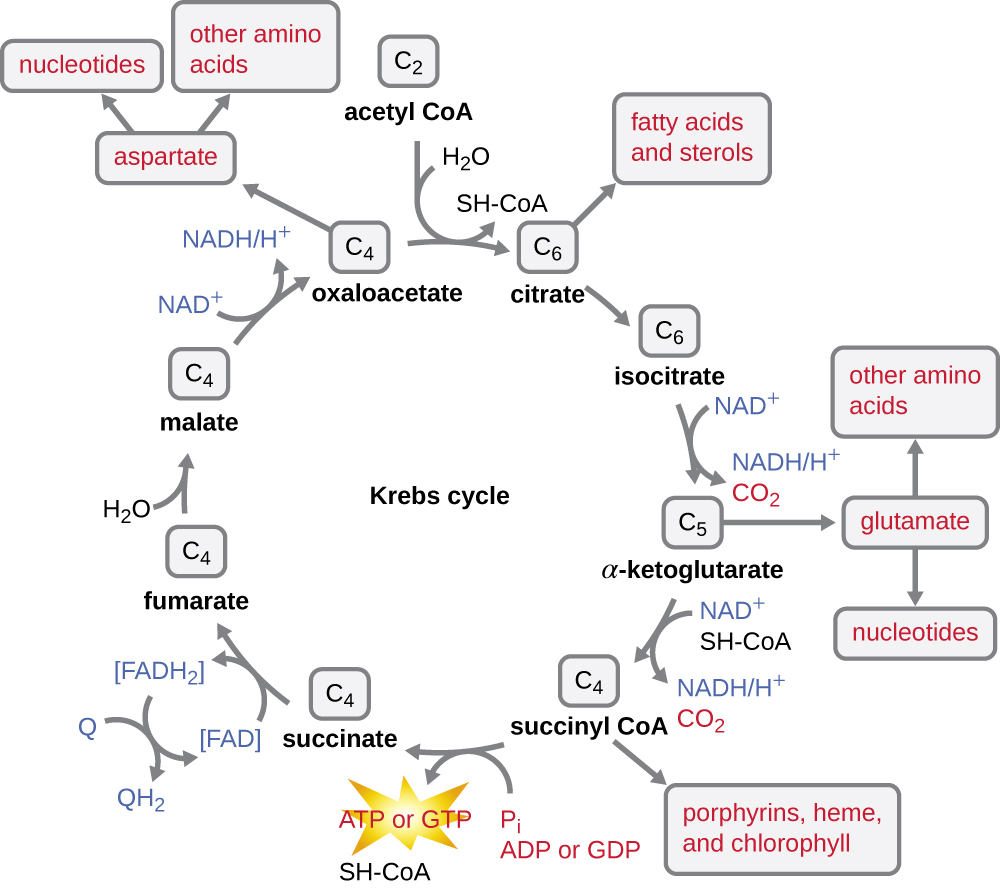
Le cycle de Krebs transfère les électrons restants du groupe acétyle produit lors de la réaction de transition vers des molécules porteuses d'électrons, les réduisant ainsi. Le cycle de Krebs se produit également dans le cytoplasme des procaryotes en même temps que la glycolyse et la réaction de transition, mais il a lieu dans la matrice mitochondriale des cellules eucaryotes où se produit également la réaction de transition. Le cycle de Krebs doit son nom à son découvreur, le scientifique britannique Hans Adolf Krebs (1900—1981) et est également appelé cycle de l'acide citrique, ou cycle de l'acide tricarboxylique (TCA) parce que l'acide citrique possède trois groupes carboxyle dans sa structure. Contrairement à la glycolyse, le cycle de Krebs est une boucle fermée : la dernière partie du processus régénère le composé utilisé dans la première étape (Figure\(\PageIndex{4}\)). Les huit étapes du cycle sont une série de réactions chimiques qui capturent le groupe acétyle à deux carbones (le support de CoA n'entre pas dans le cycle de Krebs) issu de la réaction de transition, qui est ajouté à un intermédiaire à quatre carbones dans le cycle de Krebs, produisant l'acide citrique intermédiaire à six carbones (donnant autre nom pour ce cycle). Lorsqu'un tour du cycle revient au point de départ de l'intermédiaire à quatre carbones, le cycle produit deux molécules de CO 2, une molécule d'ATP (ou une molécule équivalente, telle que le guanosine triphosphate [GTP]) produite par phosphorylation au niveau du substrat, et trois molécules de NADH et une de FADH 2 . (Une discussion et une illustration détaillée du cycle complet de Krebs figurent à l'annexe C.)
Bien que de nombreux organismes utilisent le cycle de Krebs tel que décrit dans le cadre du métabolisme du glucose, plusieurs des composés intermédiaires du cycle de Krebs peuvent être utilisés pour synthétiser une grande variété de molécules cellulaires importantes, notamment des acides aminés, des chlorophylles, des acides gras et des nucléotides ; par conséquent, le cycle est à la fois anabolique et catabolique (Figure\(\PageIndex{5}\)).
Concepts clés et résumé
- La glycolyse est la première étape de la dégradation du glucose, entraînant la formation d'ATP, produit par phosphorylation au niveau du substrat, du NADH et de deux molécules de pyruvate. La glycolyse n'utilise pas d'oxygène et ne dépend pas de l'oxygène.
- Après glycolyse, un pyruvate à trois carbones est décarboxylé pour former un groupe acétyle à deux carbones, couplé à la formation de NADH. Le groupe acétyle est lié à un gros composé porteur appelé coenzyme A.
- Après l'étape de transition, la coenzyme A transporte l'acétyle à deux carbones vers le cycle de Krebs, où les deux carbones entrent dans le cycle. À chaque tour du cycle, un groupe acétyle dérivé de la glycolyse est oxydé davantage, produisant trois molécules de NADH, une FADH 2 et un ATP par phosphorylation au niveau du substrat, et libérant deux molécules de CO 2.
- Le cycle de Krebs peut être utilisé à d'autres fins. De nombreux intermédiaires sont utilisés pour synthétiser des molécules cellulaires importantes, notamment des acides aminés, des chlorophylles, des acides gras et des nucléotides.