
3.4 : La membrane cellulaire
- Page ID
- 186117

La membrane plasmique d'une cellule définit les limites de la cellule et détermine la nature de son contact avec l'environnement. Les cellules excluent certaines substances, en absorbent d'autres et en excrètent d'autres encore, le tout en quantités contrôlées. Les membranes plasmiques entourent les frontières des cellules, mais au lieu d'être un sac statique, elles sont dynamiques et en constante évolution. La membrane plasmique doit être suffisamment souple pour permettre à certaines cellules, telles que les globules rouges et les globules blancs, de changer de forme lorsqu'elles traversent des capillaires étroits. Ce sont les fonctions les plus évidentes d'une membrane plasmique. De plus, la surface de la membrane plasmique contient des marqueurs qui permettent aux cellules de se reconnaître mutuellement, ce qui est vital lorsque les tissus et les organes se forment au début du développement, et qui joue ensuite un rôle dans la distinction entre « soi » et « non-soi » de la réponse immunitaire.
La membrane plasmique contient également des récepteurs, qui sont des sites de fixation pour des substances spécifiques qui interagissent avec la cellule. Chaque récepteur est structuré pour se lier à une substance spécifique. Par exemple, les récepteurs de surface de la membrane créent des modifications à l'intérieur, telles que des modifications des enzymes des voies métaboliques. Ces voies métaboliques peuvent être vitales pour fournir de l'énergie à la cellule, fabriquer des substances spécifiques pour la cellule ou décomposer les déchets cellulaires ou les toxines en vue de leur élimination. Les récepteurs situés à la surface extérieure de la membrane plasmique interagissent avec les hormones ou les neurotransmetteurs et permettent à leurs messages d'être transmis à la cellule. Certains sites de reconnaissance sont utilisés par les virus comme points d'attache. Bien qu'ils soient très spécifiques, des agents pathogènes tels que les virus peuvent évoluer pour exploiter les récepteurs afin d'entrer dans une cellule en imitant la substance spécifique que le récepteur est censé lier. Cette spécificité permet d'expliquer pourquoi le virus de l'immunodéficience humaine (VIH) ou l'un des cinq types de virus de l'hépatite n'envahissent que des cellules spécifiques.
Modèle Fluid Mosaic
En 1972, S. J. Singer et Garth L. Nicolson ont proposé un nouveau modèle de la membrane plasmique qui, par rapport aux connaissances antérieures, expliquait mieux à la fois les observations microscopiques et la fonction de la membrane plasmique. C'est ce qu'on a appelé le modèle de mosaïque fluide. Le modèle a quelque peu évolué au fil du temps, mais il rend encore mieux compte de la structure et des fonctions de la membrane plasmique telles que nous les comprenons aujourd'hui. Le modèle de mosaïque fluide décrit la structure de la membrane plasmique comme une mosaïque de composants, notamment des phospholipides, du cholestérol, des protéines et des glucides, dans laquelle les composants peuvent s'écouler et changer de position, tout en préservant l'intégrité fondamentale de la membrane. Les molécules de phospholipides et les protéines incorporées peuvent diffuser rapidement et latéralement dans la membrane. La fluidité de la membrane plasmique est nécessaire aux activités de certaines enzymes et molécules de transport au sein de la membrane. Les membranes plasmiques ont une épaisseur comprise entre 5 et 10 nm. À titre de comparaison, les globules rouges humains, visibles par microscopie optique, ont une épaisseur d'environ 8 µm, soit environ 1 000 fois plus épaisse qu'une membrane plasmique. (Figurine\(\PageIndex{1}\))
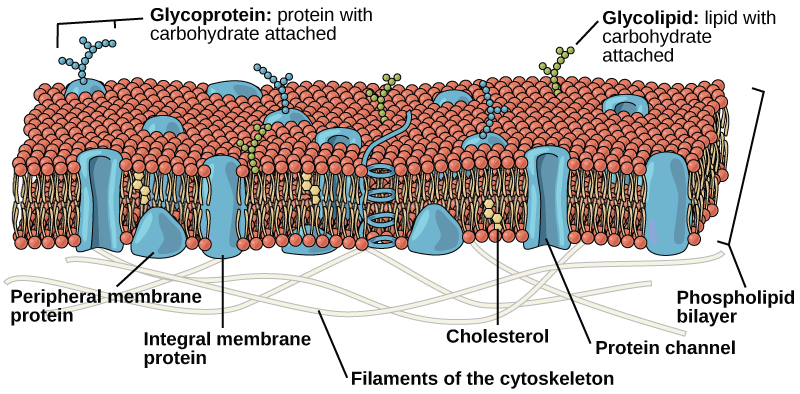
La membrane plasmique est principalement constituée d'une bicouche de phospholipides incorporant des protéines, des glucides, des glycolipides et des glycoprotéines et, dans les cellules animales, du cholestérol. La quantité de cholestérol dans les membranes plasmiques des animaux régule la fluidité de la membrane et change en fonction de la température de l'environnement cellulaire. En d'autres termes, le cholestérol agit comme antigel dans la membrane cellulaire et est plus abondant chez les animaux vivant dans des climats froids.
Le tissu principal de la membrane est composé de deux couches de molécules de phospholipides, et les extrémités polaires de ces molécules (qui ressemblent à un ensemble de boules dans une interprétation artistique du modèle) (Figure\(\PageIndex{1}\)) sont en contact avec un fluide aqueux à la fois à l'intérieur et à l'extérieur de la cellule. Ainsi, les deux surfaces de la membrane plasmique sont hydrophiles. En revanche, l'intérieur de la membrane, entre ses deux surfaces, est une région hydrophobe ou non polaire en raison des queues d'acides gras. Cette région n'est pas attirée par l'eau ou d'autres molécules polaires.
Les protéines constituent le deuxième composant chimique majeur des membranes plasmiques. Les protéines intégrales sont intégrées dans la membrane plasmique et peuvent recouvrir la totalité ou une partie de la membrane. Les protéines intégrales peuvent servir de canaux ou de pompes pour déplacer des matériaux à l'intérieur ou à l'extérieur de la cellule. Les protéines périphériques se trouvent sur les surfaces extérieures ou intérieures des membranes, fixées soit à des protéines intégrales, soit à des molécules de phospholipides. Les protéines intégrales et périphériques peuvent servir d'enzymes, de fixations structurales pour les fibres du cytosquelette ou de sites de reconnaissance de la cellule.
Les glucides sont le troisième composant majeur des membranes plasmiques. Ils se trouvent toujours sur la surface extérieure des cellules et sont liés soit à des protéines (formant des glycoprotéines) soit à des lipides (formant des glycolipides). Ces chaînes glucidiques peuvent être constituées de 2 à 60 unités monosaccharidiques et peuvent être droites ou ramifiées. Avec les protéines périphériques, les glucides forment des sites spécialisés à la surface des cellules qui permettent aux cellules de se reconnaître mutuellement.
L'ÉVOLUTION EN ACTION : Comment les virus infectent des organes spécifiques
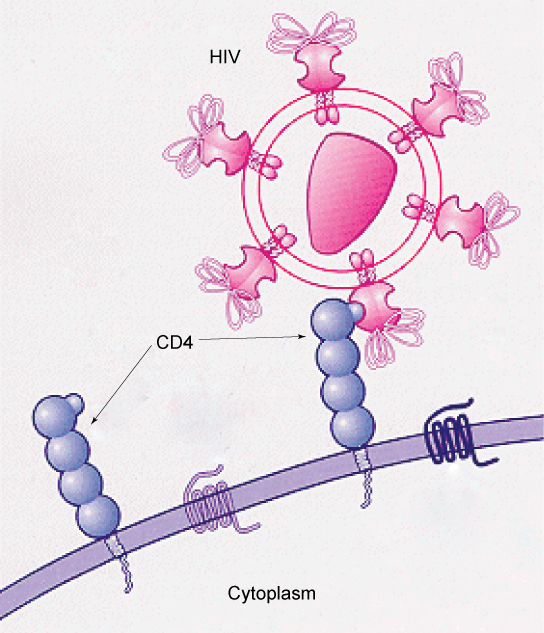
De nombreux virus exploitent des molécules de glycoprotéines spécifiques exposées à la surface des membranes cellulaires des cellules hôtes pour infecter des organes spécifiques. Par exemple, le VIH est capable de pénétrer dans les membranes plasmiques de types spécifiques de globules blancs appelés lymphocytes T auxiliaires et monocytes, ainsi que dans certaines cellules du système nerveux central. Le virus de l'hépatite n'attaque que les cellules du foie.
Ces virus sont capables d'envahir ces cellules, car les cellules possèdent des sites de liaison sur leur surface que les virus ont exploités avec des glycoprotéines tout aussi spécifiques dans leur enveloppe (Figure\(\PageIndex{2}\)). La cellule est piégée par le mimétisme des molécules de l'enveloppe du virus, et le virus peut pénétrer dans la cellule. D'autres sites de reconnaissance situés à la surface du virus interagissent avec le système immunitaire humain, incitant l'organisme à produire des anticorps. Les anticorps sont fabriqués en réponse aux antigènes (ou aux protéines associées à des agents pathogènes invasifs). Ces mêmes sites servent de lieux de fixation des anticorps et détruisent ou inhibent l'activité du virus. Malheureusement, ces sites du VIH sont codés par des gènes qui changent rapidement, ce qui rend très difficile la production d'un vaccin efficace contre le virus. La population virale d'un individu infecté évolue rapidement par mutation en différentes populations, ou variants, qui se distinguent par des différences dans ces sites de reconnaissance. Ce changement rapide des marqueurs de surface viraux diminue l'efficacité du système immunitaire de la personne à attaquer le virus, car les anticorps ne reconnaîtront pas les nouvelles variations des motifs de surface.
Résumé
La compréhension moderne de la membrane plasmique est appelée modèle de mosaïque fluide. La membrane plasmique est composée d'une bicouche de phospholipides, dont les queues d'acides gras hydrophobes sont en contact les unes avec les autres. Le paysage de la membrane est parsemé de protéines, dont certaines recouvrent la membrane. Certaines de ces protéines servent à transporter des matériaux à l'intérieur ou à l'extérieur de la cellule. Les glucides sont attachés à certaines protéines et lipides de la surface de la membrane orientée vers l'extérieur. Ils forment des complexes qui permettent d'identifier la cellule à d'autres cellules. La nature fluide de la membrane est due à la configuration des queues d'acides gras, à la présence de cholestérol incrusté dans la membrane (dans les cellules animales) et à la nature mosaïque des protéines et des complexes protéines-glucides, qui ne sont pas solidement fixés en place. Les membranes plasmiques entourent les frontières des cellules, mais au lieu d'être un sac statique, elles sont dynamiques et en constante évolution.
Lexique
- modèle de mosaïque fluide
- un modèle de la structure de la membrane plasmique sous la forme d'une mosaïque de composants, y compris les phospholipides, le cholestérol, les protéines et les glycolipides, résultant en un caractère fluide plutôt que statique