
3.3 : Cellules eucaryotes
- Page ID
- 186099

À ce stade, il doit être clair que les cellules eucaryotes ont une structure plus complexe que les cellules procaryotes. Les organites permettent à diverses fonctions de se produire simultanément dans la cellule. Avant d'aborder les fonctions des organites au sein d'une cellule eucaryote, examinons d'abord deux composants importants de la cellule : la membrane plasmique et le cytoplasme.
ART CONNECTION
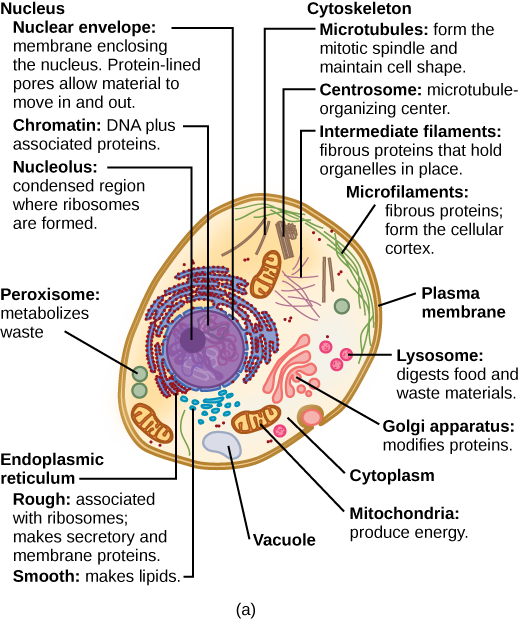
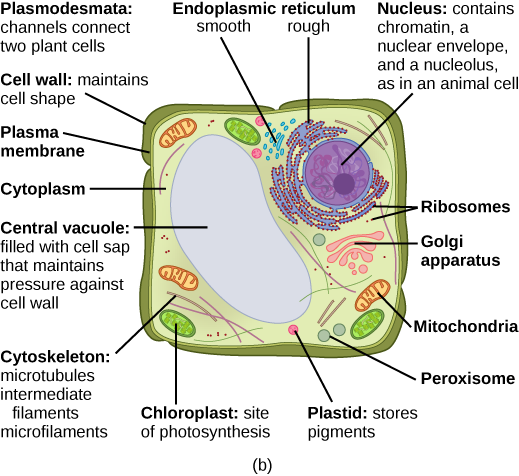
Quelles sont les structures d'une cellule végétale qu'une cellule animale n'a pas ? Quelles sont les structures d'une cellule animale qu'une cellule végétale n'a pas ?
La membrane plasmique
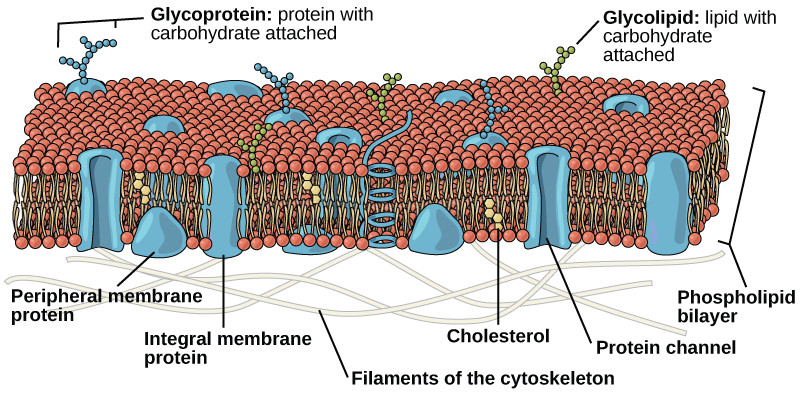
Comme les procaryotes, les cellules eucaryotes possèdent une membrane plasmique (Figure\(\PageIndex{2}\)) constituée d'une bicouche de phospholipides contenant des protéines qui sépare le contenu interne de la cellule de son environnement environnant. Un phospholipide est une molécule lipidique composée de deux chaînes d'acides gras, d'un squelette de glycérol et d'un groupe phosphate. La membrane plasmique régule le passage de certaines substances, telles que les molécules organiques, les ions et l'eau, empêchant ainsi le passage de certaines substances afin de maintenir les conditions internes, tout en introduisant ou en retirant activement d'autres. D'autres composés se déplacent passivement à travers la membrane.
Les membranes plasmiques des cellules spécialisées dans l'absorption sont repliées en projections en forme de doigts appelées microvillosités (singulier = microvillosités). Ce pliage augmente la surface de la membrane plasmique. Ces cellules recouvrent généralement l'intestin grêle, l'organe qui absorbe les nutriments contenus dans les aliments digérés. Il s'agit d'un excellent exemple de forme correspondant à la fonction d'une structure.
Les personnes atteintes de la maladie cœliaque ont une réponse immunitaire au gluten, une protéine présente dans le blé, l'orge et le seigle. La réponse immunitaire endommage les microvillosités et, par conséquent, les personnes atteintes ne peuvent pas absorber les nutriments. Cela entraîne de la malnutrition, des crampes et de la diarrhée. Les patients atteints de la maladie cœliaque doivent suivre un régime sans gluten.
Le cytoplasme
Le cytoplasme comprend le contenu d'une cellule située entre la membrane plasmique et l'enveloppe nucléaire (structure qui sera discutée prochainement). Il est composé d'organites en suspension dans le cytosol gélatineux, le cytosquelette et divers produits chimiques (Figure\(\PageIndex{1}\)). Même si le cytoplasme est composé de 70 à 80 pour cent d'eau, il a une consistance semi-solide, qui provient des protéines qu'il contient. Cependant, les protéines ne sont pas les seules molécules organiques présentes dans le cytoplasme. On y trouve également du glucose et d'autres sucres simples, des polysaccharides, des acides aminés, des acides nucléiques, des acides gras et des dérivés du glycérol. Des ions de sodium, de potassium, de calcium et de nombreux autres éléments sont également dissous dans le cytoplasme. De nombreuses réactions métaboliques, y compris la synthèse des protéines, se produisent dans le cytoplasme.
Le cytosquelette
Si vous deviez retirer tous les organites d'une cellule, la membrane plasmique et le cytoplasme seraient-ils les seuls composants restants ? Non Dans le cytoplasme, il y aurait encore des ions et des molécules organiques, ainsi qu'un réseau de fibres protéiques qui aide à maintenir la forme de la cellule, à fixer certains organites dans des positions spécifiques, à permettre au cytoplasme et aux vésicules de se déplacer au sein de la cellule et aux organismes unicellulaires de se déplacer indépendamment. Ensemble, ce réseau de fibres protéiques est connu sous le nom de cytosquelette. Il existe trois types de fibres dans le cytosquelette : les microfilaments, également appelés filaments d'actine, les filaments intermédiaires et les microtubules (Figure\(\PageIndex{3}\)).
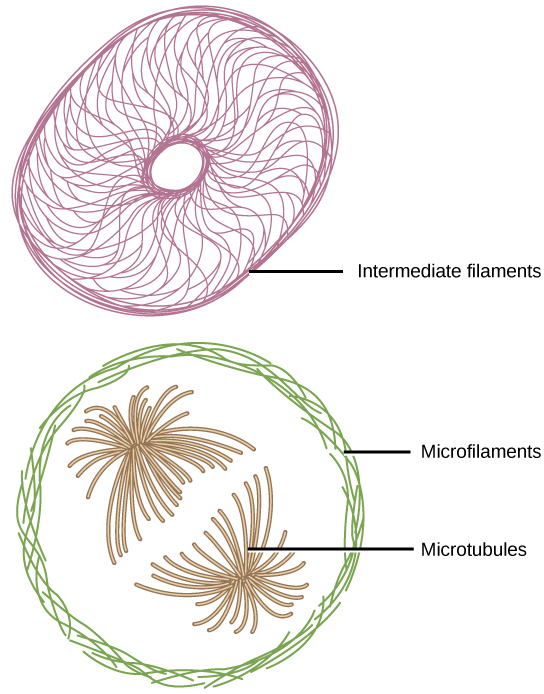
Les microfilaments sont les fibres cytosquelettiques les plus fines et servent à déplacer les composants cellulaires, par exemple lors de la division cellulaire. Ils préservent également la structure des microvillosités, c'est-à-dire le repliement important de la membrane plasmique présent dans les cellules dédiées à l'absorption. Ces composants sont également courants dans les cellules musculaires et sont responsables de la contraction des cellules musculaires. Les filaments intermédiaires ont un diamètre intermédiaire et ont des fonctions structurales, telles que le maintien de la forme de la cellule et l'ancrage des organites. La kératine, le composé qui renforce les cheveux et les ongles, forme un type de filament intermédiaire. Les microtubules sont les fibres cytosquelettiques les plus épaisses. Ce sont des tubes creux qui peuvent se dissoudre et se reformer rapidement. Les microtubules guident le mouvement des organites et sont les structures qui attirent les chromosomes vers leurs pôles lors de la division cellulaire. Ils sont également les composants structuraux des flagelles et des cils. Dans les cils et les flagelles, les microtubules sont organisés en cercle de neuf microtubules doubles à l'extérieur et de deux microtubules au centre.
Le centrosome est une région proche du noyau des cellules animales qui fonctionne comme un centre organisateur des microtubules. Il contient une paire de centrioles, deux structures perpendiculaires l'une à l'autre. Chaque centriole est un cylindre de neuf triplets de microtubules.
Le centrosome se réplique avant qu'une cellule ne se divise, et les centrioles jouent un rôle en attirant les chromosomes dupliqués vers les extrémités opposées de la cellule en division. Cependant, la fonction exacte des centrioles dans la division cellulaire n'est pas claire, car les cellules dont les centrioles ont été retirés peuvent toujours se diviser et les cellules végétales, qui n'ont pas de centrioles, sont capables de division cellulaire.
Flagella et Cilia
Les flagelles (singulier = flagelle) sont de longues structures ressemblant à des cheveux qui s'étendent à partir de la membrane plasmique et sont utilisées pour déplacer une cellule entière (par exemple, le sperme, l'Euglène). Lorsqu'elle est présente, la cellule ne possède qu'un seul flagelle ou quelques flagelles. Toutefois, lorsque des cils (singulier = cil) sont présents, ils sont nombreux et s'étendent sur toute la surface de la membrane plasmique. Il s'agit de structures courtes ressemblant à des cheveux qui sont utilisées pour déplacer des cellules entières (comme le paramécium) ou déplacer des substances le long de la surface externe de la cellule (par exemple, les cils des cellules qui tapissent les trompes de Fallope qui déplacent l'ovule vers l'utérus) ou les cils qui tapissent les cellules des voies respiratoires qui se déplacent matières particulaires vers la gorge que le mucus a piégées).
Le système endomembranaire
Le système endomembranaire (endo = within) est un groupe de membranes et d'organites (Figure\(\PageIndex{3}\)) de cellules eucaryotes qui agissent ensemble pour modifier, conditionner et transporter les lipides et les protéines. Il comprend l'enveloppe nucléaire, les lysosomes et les vésicules, le réticulum endoplasmique et l'appareil de Golgi, que nous aborderons sous peu. Bien qu'elle ne se trouve pas techniquement à l'intérieur de la cellule, la membrane plasmique est incluse dans le système endomembranaire car, comme vous le verrez, elle interagit avec les autres organites endomembraneux.
Le noyau
En général, le noyau est l'organite le plus important d'une cellule (Figure\(\PageIndex{1}\)). Le noyau (pluriel = noyaux) abrite l'ADN de la cellule sous forme de chromatine et dirige la synthèse des ribosomes et des protéines. Regardons-le plus en détail (Figure\(\PageIndex{4}\)).
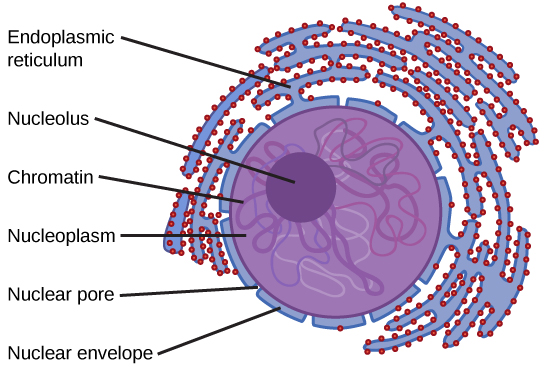
L'enveloppe nucléaire est une structure à double membrane qui constitue la partie la plus externe du noyau (Figure\(\PageIndex{4}\)). Les membranes interne et externe de l'enveloppe nucléaire sont toutes deux des bicouches de phospholipides.
L'enveloppe nucléaire est ponctuée de pores qui contrôlent le passage des ions, des molécules et de l'ARN entre le nucléoplasme et le cytoplasme.
Pour comprendre la chromatine, il est utile de considérer d'abord les chromosomes. Les chromosomes sont des structures du noyau composées d'ADN, de matériel héréditaire et de protéines. Cette combinaison d'ADN et de protéines est appelée chromatine. Chez les eucaryotes, les chromosomes sont des structures linéaires. Chaque espèce possède un nombre spécifique de chromosomes dans le noyau des cellules de son corps. Par exemple, chez l'homme, le nombre de chromosomes est de 46, alors que chez les mouches des fruits, il est de huit.
Les chromosomes ne sont visibles et peuvent être distingués les uns des autres que lorsque la cellule s'apprête à se diviser. Lorsque la cellule est dans les phases de croissance et de maintien de son cycle de vie, les chromosomes ressemblent à un ensemble de fils déroulés et mélangés.
Nous savons déjà que le noyau dirige la synthèse des ribosomes, mais comment s'y prend-il ? Certains chromosomes possèdent des sections d'ADN qui codent pour l'ARN ribosomal. Une zone sombre du noyau, appelée nucléole (pluriel = nucléoles), agrège l'ARN ribosomal avec les protéines associées pour assembler les sous-unités ribosomales qui sont ensuite transportées par les pores nucléaires jusqu'au cytoplasme.
Le réticulum endoplasmique
Le réticulum endoplasmique (RE) (Figure\(\PageIndex{7}\)) est une série de tubules membraneux interconnectés qui modifient collectivement les protéines et synthétisent les lipides. Cependant, ces deux fonctions sont exécutées dans des zones distinctes du réticulum endoplasmique : le réticulum endoplasmique rugueux et le réticulum endoplasmique lisse, respectivement.
La partie creuse des tubules ER est appelée lumen ou espace cisternal. La membrane du RE, qui est une bicouche de phospholipides incorporée à des protéines, est continue avec l'enveloppe nucléaire.
Le réticulum endoplasmique rugueux (RER) est ainsi nommé parce que les ribosomes attachés à sa surface cytoplasmique lui confèrent une apparence cloutée lorsqu'on l'observe au microscope électronique.
Les ribosomes synthétisent des protéines lorsqu'ils sont attachés au RE, ce qui entraîne le transfert de leurs protéines nouvellement synthétisées dans la lumière du RER où ils subissent des modifications telles que le repliement ou l'ajout de sucres. Le RER fabrique également des phospholipides pour les membranes cellulaires.
Si les phospholipides ou les protéines modifiées ne sont pas destinés à rester dans le RER, ils seront emballés dans des vésicules et transportés depuis le RER par bourgeonnement depuis la membrane (Figure\(\PageIndex{7}\)). Comme le RER est engagé dans la modification des protéines qui seront sécrétées par la cellule, il est abondant dans les cellules qui sécrètent des protéines, comme le foie.
Le réticulum endoplasmique lisse (SER) est continu avec le RER mais ne possède que peu ou pas de ribosomes sur sa surface cytoplasmique (voir Figure\(\PageIndex{1}\)). Les fonctions du SER incluent la synthèse des glucides, des lipides (y compris les phospholipides) et des hormones stéroïdiennes, la désintoxication des médicaments et des poisons, le métabolisme de l'alcool et le stockage des ions calcium.
L'appareil de Golgi
Nous avons déjà mentionné que les vésicules peuvent bourgeonner à partir des urgences, mais où vont les vésicules ? Avant d'atteindre leur destination finale, les lipides ou les protéines contenus dans les vésicules de transport doivent être triés, emballés et étiquetés afin qu'ils se retrouvent au bon endroit. Le tri, le marquage, le conditionnement et la distribution des lipides et des protéines ont lieu dans l'appareil de Golgi (également appelé corps de Golgi), une série de sacs membraneux aplatis (Figure\(\PageIndex{5}\)).
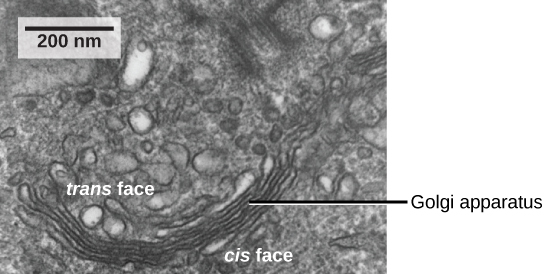
L'appareil de Golgi possède une face de réception située à proximité du réticulum endoplasmique et une face de libération située du côté opposé au RE, en direction de la membrane cellulaire. Les vésicules de transport qui se forment à partir des urgences se déplacent vers la face réceptrice, fusionnent avec celle-ci et déversent leur contenu dans la lumière de l'appareil de Golgi. Au fur et à mesure que les protéines et les lipides traversent le Golgi, ils subissent d'autres modifications. La modification la plus fréquente est l'ajout de courtes chaînes de molécules de sucre. Les protéines et les lipides récemment modifiés sont ensuite marqués par de petits groupes moléculaires afin de leur permettre d'être acheminés vers leurs destinations appropriées.
Enfin, les protéines modifiées et marquées sont emballées dans des vésicules qui bourgeonnent à partir de la face opposée du Golgi. Alors que certaines de ces vésicules transportent des vésicules et déposent leur contenu dans d'autres parties de la cellule où elles seront utilisées, d'autres, des vésicules sécrétoires, fusionnent avec la membrane plasmique et libèrent leur contenu à l'extérieur de la cellule.
La quantité de Golgi dans les différents types de cellules montre à nouveau que la forme suit la fonction au sein des cellules. Les cellules qui exercent une activité sécrétoire importante (telles que les cellules des glandes salivaires qui sécrètent des enzymes digestives ou les cellules du système immunitaire qui sécrètent des anticorps) contiennent un nombre abondant de Golgi.
Dans les cellules végétales, le Golgi joue un rôle supplémentaire dans la synthèse de polysaccharides, dont certains sont incorporés dans la paroi cellulaire et d'autres sont utilisés dans d'autres parties de la cellule.
Lysosomes
Dans les cellules animales, les lysosomes sont les « déchets » de la cellule. Les enzymes digestives présentes dans les lysosomes contribuent à la dégradation des protéines, des polysaccharides, des lipides, des acides nucléiques et même des organites usés. Chez les eucaryotes unicellulaires, les lysosomes sont importants pour la digestion des aliments qu'ils ingèrent et pour le recyclage des organites. Ces enzymes sont actives à un pH beaucoup plus bas (plus acide) que ceux situés dans le cytoplasme. De nombreuses réactions qui se produisent dans le cytoplasme ne peuvent pas se produire à un faible pH, de sorte que l'avantage de compartimenter la cellule eucaryote en organites est évident.
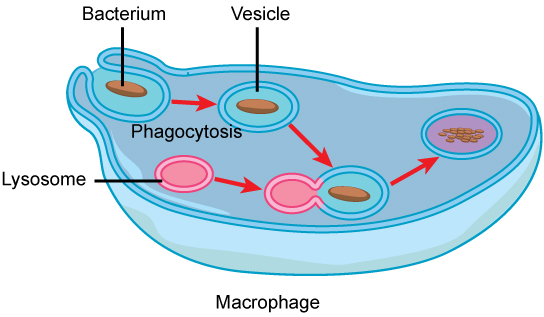
Les lysosomes utilisent également leurs enzymes hydrolytiques pour détruire les organismes pathogènes susceptibles de pénétrer dans la cellule. Un bon exemple de cela se produit dans un groupe de globules blancs appelés macrophages, qui font partie du système immunitaire de votre corps. Lors d'un processus connu sous le nom de phagocytose, une section de la membrane plasmique du macrophage invagine (se replie) et engloutit un pathogène. La section invaginée, avec le pathogène à l'intérieur, se pince ensuite de la membrane plasmique et devient une vésicule. La vésicule fusionne avec un lysosome. Les enzymes hydrolytiques du lysosome détruisent ensuite le pathogène (Figure\(\PageIndex{6}\)).
Vésicules et vacuoles
Les vésicules et les vacuoles sont des sacs liés à la membrane qui servent au stockage et au transport. Les vacuoles sont un peu plus grosses que les vésicules et la membrane d'une vacuole ne fusionne pas avec les membranes des autres composants cellulaires. Les vésicules peuvent fusionner avec d'autres membranes du système cellulaire. De plus, les enzymes présentes dans les vacuoles végétales peuvent décomposer les macromolécules.
ART CONNECTION
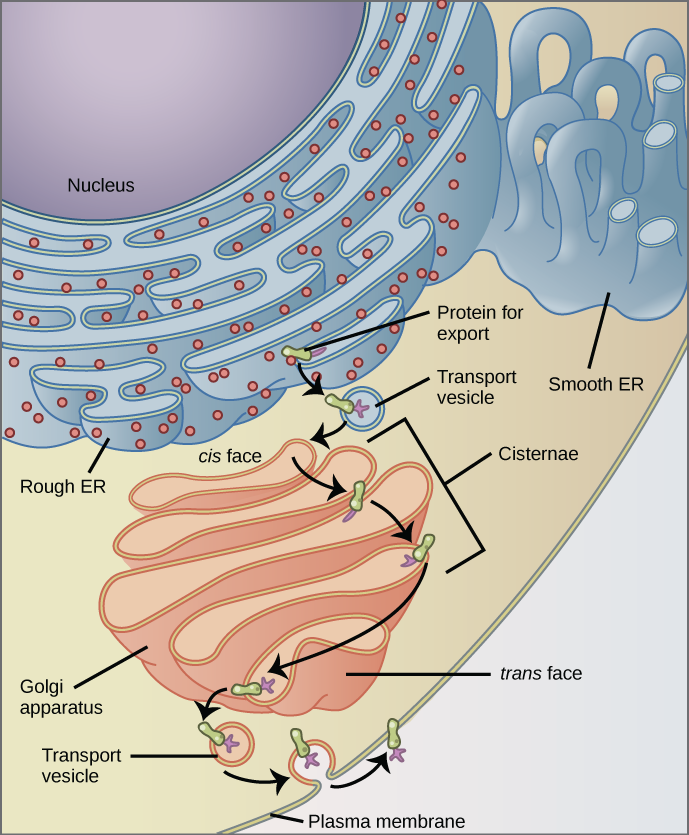
Pourquoi la face cis du Golgi ne fait-elle pas face à la membrane plasmique ?
Ribosomes
Les ribosomes sont les structures cellulaires responsables de la synthèse des protéines. Lorsqu'ils sont observés au microscope électronique, les ribosomes libres apparaissent sous forme d'amas ou de petits points individuels flottant librement dans le cytoplasme. Les ribosomes peuvent être fixés soit à la face cytoplasmique de la membrane plasmique, soit à la face cytoplasmique du réticulum endoplasmique (Figure\(\PageIndex{7}\)). La microscopie électronique a montré que les ribosomes sont constitués de grandes et de petites sous-unités. Les ribosomes sont des complexes enzymatiques responsables de la synthèse des protéines.
La synthèse des protéines étant essentielle pour toutes les cellules, les ribosomes sont présents dans pratiquement toutes les cellules, bien qu'ils soient plus petits dans les cellules procaryotes. Ils sont particulièrement abondants dans les globules rouges immatures pour la synthèse de l'hémoglobine, qui joue un rôle dans le transport de l'oxygène dans tout le corps.
Mitochondries
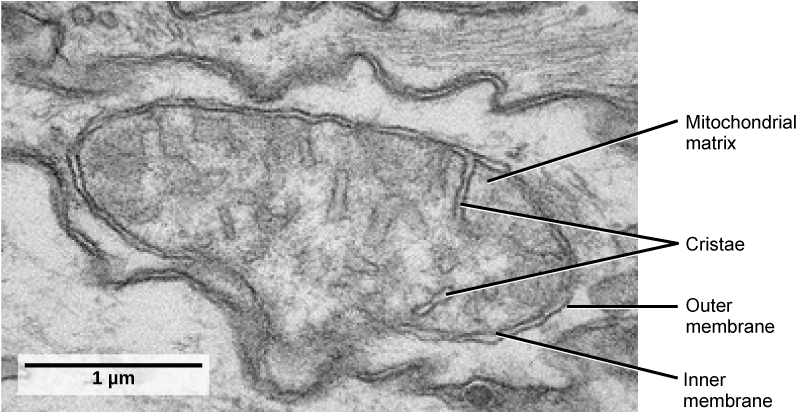
Les mitochondries (singulier = mitochondrie) sont souvent appelées les « centrales » ou les « usines énergétiques » d'une cellule parce qu'elles sont responsables de la fabrication de l'adénosine triphosphate (ATP), la principale molécule porteuse d'énergie de la cellule. La formation d'ATP à partir de la dégradation du glucose est connue sous le nom de respiration cellulaire. Les mitochondries sont des organites à double membrane de forme ovale (Figure\(\PageIndex{8}\)) qui possèdent leurs propres ribosomes et ADN. Chaque membrane est une bicouche de phospholipides incorporée à des protéines. La couche interne comporte des plis appelés crêtes, qui augmentent la surface de la membrane interne. La zone entourée par les plis est appelée matrice mitochondriale. Les crêtes et la matrice jouent des rôles différents dans la respiration cellulaire.
Conformément à notre thème selon lequel la forme suit la fonction, il est important de souligner que les cellules musculaires ont une très forte concentration de mitochondries, car les cellules musculaires ont besoin de beaucoup d'énergie pour se contracter.
Peroxysomes
Les peroxysomes sont de petits organites ronds entourés de membranes uniques. Ils effectuent des réactions d'oxydation qui décomposent les acides gras et les acides aminés. Ils détoxifient également de nombreux poisons qui peuvent pénétrer dans l'organisme. L'alcool est détoxifié par les peroxysomes dans les cellules hépatiques. Un sous-produit de ces réactions d'oxydation est le peroxyde d'hydrogène, H 2 O 2, qui est contenu dans les peroxysomes pour empêcher le produit chimique d'endommager les composants cellulaires situés à l'extérieur de l'organite. Le peroxyde d'hydrogène est décomposé en toute sécurité par les enzymes peroxysomales en eau et en oxygène.
Cellules animales contre cellules végétales
Malgré leurs similitudes fondamentales, il existe des différences frappantes entre les cellules animales et végétales (voir tableau\(\PageIndex{1}\)). Les cellules animales possèdent des centrioles, des centrosomes (dont il est question dans le cytosquelette) et des lysosomes, alors que les cellules végétales n'en possèdent pas. Les cellules végétales possèdent une paroi cellulaire, des chloroplastes, des plasmodesmes et des plastides utilisés pour le stockage, ainsi qu'une grande vacuole centrale, contrairement aux cellules animales.
La paroi cellulaire
Dans la Figure\(\PageIndex{1}\) b, le schéma d'une cellule végétale, vous pouvez voir une structure externe à la membrane plasmique appelée paroi cellulaire. La paroi cellulaire est un revêtement rigide qui protège la cellule, fournit un support structurel et donne forme à la cellule. Les cellules fongiques et protistes possèdent également des parois cellulaires.
Alors que le principal composant des parois des cellules procaryotes est le peptidoglycane, la principale molécule organique de la paroi cellulaire végétale est la cellulose, un polysaccharide composé de longues chaînes droites d'unités de glucose. Lorsque l'information nutritionnelle fait référence aux fibres alimentaires, elle fait référence à la teneur en cellulose des aliments.
Chloroplastes
Comme les mitochondries, les chloroplastes possèdent également leur propre ADN et leurs propres ribosomes. Les chloroplastes agissent dans la photosynthèse et peuvent être trouvés dans les cellules eucaryotes telles que les plantes et les algues. Lors de la photosynthèse, le dioxyde de carbone, l'eau et l'énergie lumineuse sont utilisés pour produire du glucose et de l'oxygène. C'est la principale différence entre les plantes et les animaux : les plantes (autotrophes) peuvent produire leur propre nourriture, comme le glucose, tandis que les animaux (hétérotrophes) doivent compter sur d'autres organismes pour leurs composés organiques ou leur source de nourriture.
Comme les mitochondries, les chloroplastes possèdent des membranes externe et interne, mais dans l'espace délimité par la membrane interne d'un chloroplaste se trouvent un ensemble de sacs membranaires interconnectés et empilés remplis de liquide appelés thylakoïdes (Figure\(\PageIndex{9}\)). Chaque pile de thylakoïdes est appelée granum (pluriel = grana). Le liquide enfermé par la membrane interne et entourant le grana est appelé stroma.
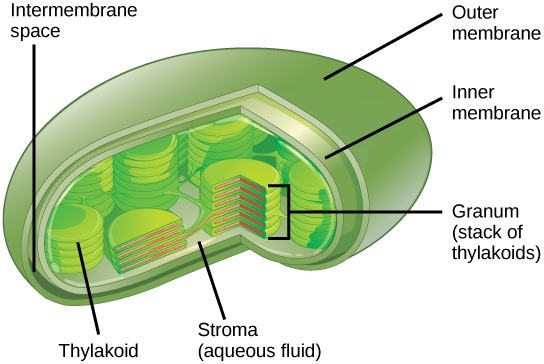
Les chloroplastes contiennent un pigment vert appelé chlorophylle, qui capte l'énergie du soleil pour la photosynthèse. Comme les cellules végétales, les protistes photosynthétiques possèdent également des chloroplastes. Certaines bactéries effectuent également la photosynthèse, mais elles ne possèdent pas de chloroplastes. Leurs pigments photosynthétiques sont situés dans la membrane thylakoïde de la cellule elle-même.
L'ÉVOLUTION EN ACTION : L'endosymbiose
Nous avons mentionné que les mitochondries et les chloroplastes contiennent de l'ADN et des ribosomes. Vous êtes-vous demandé pourquoi ? Des preuves solides indiquent que l'endosymbiose en est l'explication.
La symbiose est une relation dans laquelle des organismes de deux espèces distinctes vivent en étroite association et présentent généralement des adaptations spécifiques les uns aux autres. L'endosymbiose (endo- = interne) est une relation dans laquelle un organisme vit à l'intérieur de l'autre. Les relations endosymbiotiques abondent dans la nature. Les microbes qui produisent de la vitamine K vivent dans l'intestin humain. Cette relation est bénéfique pour nous car nous ne sommes pas en mesure de synthétiser la vitamine K. Elle est également bénéfique pour les microbes, car ils sont protégés des autres organismes et bénéficient d'un habitat stable et d'une nourriture abondante en vivant dans le gros intestin.
Les scientifiques ont remarqué depuis longtemps que les bactéries, les mitochondries et les chloroplastes sont de taille similaire. Nous savons également que les mitochondries et les chloroplastes possèdent de l'ADN et des ribosomes, tout comme les bactéries. Les scientifiques pensent que les cellules hôtes et les bactéries ont formé une relation endosymbiotique mutuellement bénéfique lorsque les cellules hôtes ont ingéré des bactéries aérobies et des cyanobactéries sans les détruire. Au fil de l'évolution, ces bactéries ingérées se sont spécialisées dans leurs fonctions, les bactéries aérobies devenant des mitochondries et les bactéries photosynthétiques des chloroplastes.
La vacuole centrale
Nous avons déjà mentionné les vacuoles en tant que composants essentiels des cellules végétales. Si vous regardez la Figure\(\PageIndex{1}\), vous verrez que les cellules végétales possèdent chacune une grande vacuole centrale qui occupe la majeure partie de la cellule. La vacuole centrale joue un rôle clé dans la régulation de la concentration d'eau de la cellule dans des conditions environnementales changeantes. Dans les cellules végétales, le liquide à l'intérieur de la vacuole centrale fournit une pression de turgescence, qui est la pression extérieure provoquée par le fluide à l'intérieur de la cellule. Avez-vous déjà remarqué que si vous oubliez d'arroser une plante pendant quelques jours, elle flétrit ? En effet, lorsque la concentration d'eau dans le sol devient inférieure à celle de la plante, l'eau quitte les vacuoles centrales et le cytoplasme pour se retrouver dans le sol. Lorsque la vacuole centrale se rétrécit, elle laisse la paroi cellulaire sans support. Cette perte de soutien des parois cellulaires d'une plante se traduit par un aspect flétri. De plus, ce liquide a un goût très amer, ce qui décourage la consommation par les insectes et les animaux. La vacuole centrale sert également à emmagasiner les protéines dans les cellules de la graine en développement.
Matrice extracellulaire de cellules animales
La plupart des cellules animales libèrent des matières dans l'espace extracellulaire. Les principaux composants de ces matériaux sont les glycoprotéines et la protéine collagène. Ensemble, ces matériaux sont appelés matrice extracellulaire (Figure\(\PageIndex{10}\)). Non seulement la matrice extracellulaire maintient les cellules ensemble pour former un tissu, mais elle permet également aux cellules du tissu de communiquer entre elles.
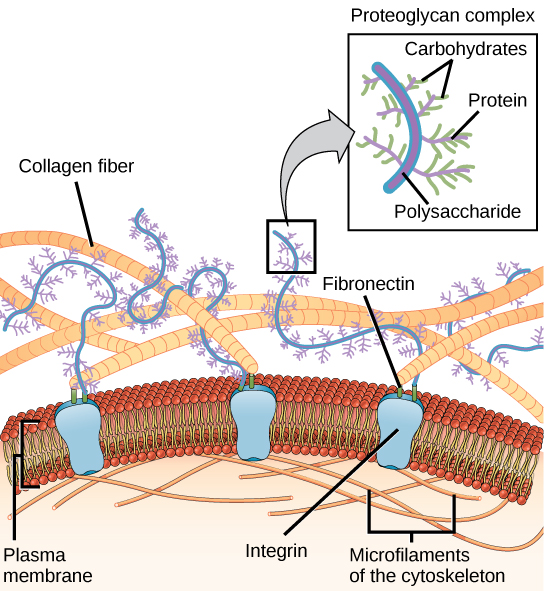
La coagulation du sang fournit un exemple du rôle de la matrice extracellulaire dans la communication cellulaire. Lorsque les cellules qui tapissent un vaisseau sanguin sont endommagées, elles présentent un récepteur protéique appelé facteur tissulaire. Lorsque le facteur tissulaire se lie à un autre facteur de la matrice extracellulaire, il provoque l'adhésion des plaquettes à la paroi du vaisseau sanguin endommagé, stimule la contraction des cellules musculaires lisses adjacentes du vaisseau sanguin (resserrant ainsi le vaisseau sanguin) et initie une série d'étapes qui stimulent des plaquettes pour produire des facteurs de coagulation.
Jonctions intercellulaires
Les cellules peuvent également communiquer entre elles par contact direct, ce que l'on appelle des jonctions intercellulaires. Il existe certaines différences dans la manière dont les cellules végétales et animales s'y prennent. Les plasmodesmes (singulier = plasmodesma) sont des jonctions entre les cellules végétales, tandis que les contacts entre cellules animales comprennent des jonctions serrées et lacunaires et des desmosomes.
En général, de longues parties des membranes plasmiques des cellules végétales voisines ne peuvent pas se toucher car elles sont séparées par les parois cellulaires qui entourent chaque cellule. Les plasmodesmes sont de nombreux canaux qui passent entre les parois cellulaires des cellules végétales adjacentes, reliant leur cytoplasme et permettant aux molécules signaux et aux nutriments d'être transportés d'\(\PageIndex{11}\)une cellule à l'autre (Figure a).
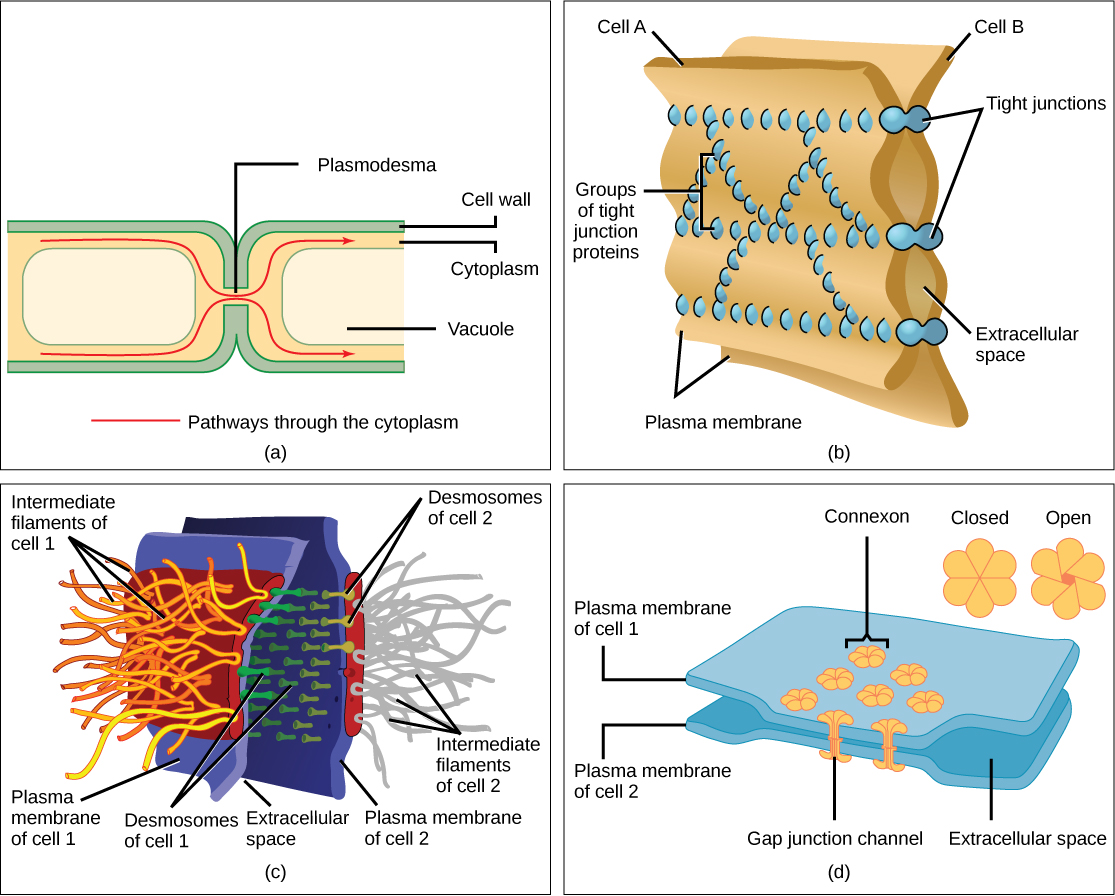
Une jonction étanche est un joint étanche entre deux cellules animales adjacentes (Figure\(\PageIndex{11}\) b). Les protéines maintiennent fermement les cellules les unes contre les autres. Cette adhérence étanche empêche les fuites de matériaux entre les cellules. Les jonctions serrées se trouvent généralement dans le tissu épithélial qui tapisse les organes internes et les cavités et qui compose la majeure partie de la peau. Par exemple, les jonctions étroites des cellules épithéliales qui tapissent la vessie empêchent l'urine de s'écouler dans l'espace extracellulaire.
Les desmosomes, qui agissent comme des points de soudure entre les cellules épithéliales adjacentes, ne se trouvent également que dans les cellules animales (Figure\(\PageIndex{11}\) c). Ils maintiennent les cellules ensemble dans une formation semblable à une feuille dans les organes et les tissus qui s'étirent, comme la peau, le cœur et les muscles.
Les jonctions lacunaires des cellules animales ressemblent à des plasmodesmes dans les cellules végétales en ce sens qu'elles constituent des canaux entre des cellules adjacentes qui permettent le transport d'ions, de nutriments et d'autres substances qui permettent aux cellules de communiquer (Figure\(\PageIndex{11}\) d). Sur le plan structurel, cependant, les jonctions lacunaires et les plasmodesmes diffèrent.
| Composant cellulaire | Fonction | Présent chez les procaryotes ? | Présent dans les cellules animales ? | Présent dans les cellules végétales ? |
|---|---|---|---|---|
| Membrane plasmique | Sépare la cellule de l'environnement externe ; contrôle le passage des molécules organiques, des ions, de l'eau, de l'oxygène et des déchets à l'intérieur et à l'extérieur de la cellule | Oui | Oui | Oui |
| Cytoplasme | Fournit de la structure à la cellule ; siège de nombreuses réactions métaboliques ; milieu dans lequel se trouvent les organites | Oui | Oui | Oui |
| Nucléoïde | Localisation de l'ADN | Oui | Non | Non |
| Noyau | Organite cellulaire qui abrite l'ADN et dirige la synthèse des ribosomes et des protéines | Non | Oui | Oui |
| Ribosomes | Synthèse de protéines | Oui | Oui | Oui |
| Mitochondries | Production d'ATP/respiration cellulaire | Non | Oui | Oui |
| Peroxysomes | Oxyde et décompose les acides gras et les acides aminés, et détoxifie les poisons | Non | Oui | Oui |
| Vésicules et vacuoles | Stockage et transport ; fonction digestive dans les cellules végétales | Non | Oui | Oui |
| Centrosome | Rôle non précisé dans la division cellulaire des cellules animales ; centre organisateur des microtubules dans les cellules animales | Non | Oui | Non |
| Lysosomes | Digestion des macromolécules ; recyclage des organites usés | Non | Oui | Non |
| Paroi cellulaire | Protection, soutien structurel et maintien de la forme cellulaire | Oui, principalement du peptidoglycane dans les bactéries, mais pas chez les Archaea | Non | Oui, principalement de la cellulose |
| Chloroplastes | Photosynthèse | Non | Non | Oui |
| Réticulum endoplasmique | Modifie les protéines et synthétise les lipides | Non | Oui | Oui |
| Appareil Golgi | Modifie, trie, étiquette, conditionne et distribue les lipides et les protéines | Non | Oui | Oui |
| Cytosquelette | Maintient la forme de la cellule, maintient les organites dans des positions spécifiques, permet au cytoplasme et aux vésicules de se déplacer à l'intérieur de la cellule et permet aux organismes unicellulaires de se déplacer indépendamment | Oui | Oui | Oui |
| Flagelles | Locomotion cellulaire | Certains | Certains | Non, sauf pour certains spermatozoïdes végétaux |
| Cilia | Locomotion cellulaire, mouvement des particules le long de la surface extracellulaire de la membrane plasmique et filtration | Non | Certains | Non |
Résumé
Comme une cellule procaryote, une cellule eucaryote possède une membrane plasmique, un cytoplasme et des ribosomes, mais une cellule eucaryote est généralement plus grande qu'une cellule procaryote, possède un véritable noyau (c'est-à-dire que son ADN est entouré d'une membrane) et possède d'autres organites liés à la membrane qui permettent de compartimenter les fonctions. La membrane plasmique est une bicouche de phospholipides incorporée à des protéines. Le nucléole situé à l'intérieur du noyau est le site d'assemblage des ribosomes. Les ribosomes se trouvent dans le cytoplasme ou sont attachés au côté cytoplasmique de la membrane plasmique ou du réticulum endoplasmique. Ils effectuent la synthèse des protéines. Les mitochondries assurent la respiration cellulaire et produisent de l'ATP. Les peroxysomes décomposent les acides gras, les acides aminés et certaines toxines. Les vésicules et les vacuoles sont des compartiments de stockage et de transport. Dans les cellules végétales, les vacuoles aident également à décomposer les macromolécules.
Les cellules animales possèdent également un centrosome et des lysosomes. Le centrosome possède deux corps, les centrioles, dont le rôle dans la division cellulaire est inconnu. Les lysosomes sont les organites digestifs des cellules animales.
Les cellules végétales possèdent une paroi cellulaire, des chloroplastes et une vacuole centrale. La paroi cellulaire de la plante, dont le composant principal est la cellulose, protège la cellule, fournit un soutien structurel et donne forme à la cellule. La photosynthèse a lieu dans les chloroplastes. La vacuole centrale se dilate, agrandissant la cellule sans qu'il soit nécessaire de produire davantage de cytoplasme.
Le système endomembranaire comprend l'enveloppe nucléaire, le réticulum endoplasmique, l'appareil de Golgi, les lysosomes, les vésicules ainsi que la membrane plasmique. Ces composants cellulaires agissent ensemble pour modifier, conditionner, étiqueter et transporter les lipides et les protéines membranaires.
Le cytosquelette contient trois types différents d'éléments protéiques. Les microfilaments confèrent de la rigidité et de la forme à la cellule et facilitent les mouvements cellulaires. Les filaments intermédiaires supportent la tension et ancrent le noyau et les autres organites en place. Les microtubules aident la cellule à résister à la compression, servent de pistes aux protéines motrices qui déplacent les vésicules dans la cellule et attirent les chromosomes répliqués vers les extrémités opposées d'une cellule en division. Ils sont également les éléments structuraux des centrioles, des flagelles et des cils.
Les cellules animales communiquent par l'intermédiaire de leurs matrices extracellulaires et sont reliées entre elles par des jonctions serrées, des desmosomes et des jonctions lacunaires. Les cellules végétales sont connectées et communiquent entre elles par des plasmodesmes.
Connexions artistiques
Figure\(\PageIndex{1}\) : Quelles sont les structures d'une cellule végétale qu'une cellule animale n'a pas ? Quelles sont les structures d'une cellule animale qu'une cellule végétale n'a pas ?
- Réponse
-
Les cellules végétales possèdent des plasmodesmes, une paroi cellulaire, une grande vacuole centrale, des chloroplastes et des plastes. Les cellules animales possèdent des lysosomes et des centrosomes.
Figure\(\PageIndex{7}\) : Pourquoi la face cis du Golgi ne fait-elle pas face à la membrane plasmique ?
- Réponse
-
Parce que cette face reçoit des produits chimiques des urgences, qui se trouvent au centre de la cellule.
Lexique
- paroi cellulaire
- un revêtement cellulaire rigide composé de cellulose dans les plantes, de peptidoglycane dans les bactéries, de composés non peptidoglycanes dans les archées et de chitine dans les champignons qui protège la cellule, fournit un soutien structurel et donne forme à la cellule
- vacuole centrale
- un gros organite cellulaire végétal qui sert de compartiment de stockage, de réservoir d'eau et de site de dégradation des macromolécules
- chloroplaste
- un organite de cellules végétales qui effectue la photosynthèse
- cil
- (pluriel : cils) structure courte ressemblant à des cheveux qui s'étend en grand nombre à partir de la membrane plasmique et qui est utilisée pour déplacer une cellule entière ou déplacer des substances le long de la surface externe de la cellule
- cytoplasme
- toute la région située entre la membrane plasmique et l'enveloppe nucléaire, constituée d'organites en suspension dans le cytosol semblable à un gel, du cytosquelette et de divers produits chimiques
- cytosquelette
- le réseau de fibres protéiques qui, collectivement, maintient la forme de la cellule, maintient certains organites dans des positions spécifiques, permet au cytoplasme et aux vésicules de se déplacer au sein de la cellule et permet aux organismes unicellulaires de se déplacer
- cytosol
- la matière gélatineuse du cytoplasme dans laquelle les structures cellulaires sont suspendues
- desmosome
- une liaison entre les cellules épithéliales adjacentes qui se forme lorsque les cadhérines de la membrane plasmique se fixent à des filaments intermédiaires
- système endomembranaire
- groupe d'organites et de membranes des cellules eucaryotes qui agissent ensemble pour modifier, conditionner et transporter les lipides et les protéines
- réticulum endoplasmique (RE)
- une série de structures membraneuses interconnectées au sein de cellules eucaryotes qui modifient collectivement les protéines et synthétisent les lipides
- matrice extracellulaire
- le matériel, principalement du collagène, des glycoprotéines et des protéoglycanes, sécrété par des cellules animales qui maintient les cellules ensemble en tant que tissu, permet aux cellules de communiquer entre elles et fournit une protection mécanique et un ancrage aux cellules du tissu
- flagelle
- (Pluriel : flagelle) longue structure semblable à un poil qui part de la membrane plasmique et sert à déplacer la cellule
- jonction lacunaire
- un canal entre deux cellules animales adjacentes qui permet aux ions, aux nutriments et à d'autres substances de faible poids moléculaire de passer entre les cellules, permettant ainsi aux cellules de communiquer
- Appareil Golgi
- un organite eucaryote composé d'une série de membranes empilées qui trie, étiquette et conditionne les lipides et les protéines en vue de leur distribution
- lysosome
- un organite d'une cellule animale qui fait office de composant digestif de la cellule ; il décompose les protéines, les polysaccharides, les lipides, les acides nucléiques et même les organites usés
- mitochondries
- (singulier : mitochondrie) les organites cellulaires responsables de la respiration cellulaire, entraînant la production d'ATP, la principale molécule porteuse d'énergie de la cellule
- enveloppe nucléaire
- la structure à double membrane qui constitue la partie la plus externe du noyau
- nucléole
- le corps foncé du noyau qui est responsable de l'assemblage des sous-unités ribosomales
- noyau
- l'organite cellulaire qui abrite l'ADN de la cellule et dirige la synthèse des ribosomes et des protéines
- peroxysome
- un petit organite rond qui contient du peroxyde d'hydrogène, oxyde les acides gras et les acides aminés et détoxifie de nombreux poisons
- membrane plasmique
- une bicouche de phospholipides contenant des protéines intégrées (intégrales) ou attachées (périphériques) qui sépare le contenu interne de la cellule de son environnement environnant
- plasmodesme
- (pluriel : plasmodesmes) canal qui passe entre les parois cellulaires des cellules végétales adjacentes, relie leur cytoplasme et permet le transport de matériaux d'une cellule à l'autre
- ribosome
- une structure cellulaire qui réalise la synthèse des protéines
- réticulum endoplasmique rugueux (RER)
- région du réticulum endoplasmique parsemée de ribosomes et participant à la modification des protéines
- réticulum endoplasmique lisse (SER)
- région du réticulum endoplasmique qui possède peu ou pas de ribosomes sur sa surface cytoplasmique et synthétise les glucides, les lipides et les hormones stéroïdiennes ; détoxifie les produits chimiques tels que les pesticides, les conservateurs, les médicaments et les polluants environnementaux, et stocke les ions calcium
- jonction étanche
- un joint ferme entre deux cellules animales adjacentes créé par l'adhérence aux protéines
- vacuole
- un sac membranaire, légèrement plus grand qu'une vésicule, qui sert au stockage et au transport des cellules
- vésicule
- un petit sac lié à une membrane qui sert au stockage et au transport cellulaires ; sa membrane est capable de fusionner avec la membrane plasmique et les membranes du réticulum endoplasmique et de l'appareil de Golgi