
47.1 : La crise de la biodiversité
- Page ID
- 189713

Compétences à développer
- Définir la biodiversité
- Décrire la biodiversité comme l'équilibre des fluctuations naturelles des taux d'extinction et de spéciation
- Identifier les causes historiques des taux d'extinction élevés dans l'histoire de la Terre
Traditionnellement, les écologistes mesuraient la biodiversité, terme général désignant la variété présente dans la biosphère, en tenant compte à la fois du nombre d'espèces et de leur caractère commun. La biodiversité peut être estimée à différents niveaux d'organisation du vivant. Ces indices d'estimation, issus de la théorie de l'information, sont très utiles comme première étape pour quantifier la biodiversité entre et au sein des écosystèmes ; ils le sont moins lorsque la principale préoccupation des biologistes de la conservation est simplement la perte de biodiversité. Les biologistes reconnaissent toutefois que les mesures de la biodiversité, en termes de diversité des espèces, peuvent aider à concentrer les efforts visant à préserver les éléments importants de la biodiversité sur le plan biologique ou technologique.
Les cichlidés du lac Victoria sont un exemple qui nous permet de commencer à comprendre la biodiversité. Les biologistes qui ont étudié les cichlidés dans les années 1980 ont découvert des centaines d'espèces de cichlidés représentant diverses spécialisations liées à des types d'habitats particuliers et à des stratégies alimentaires spécifiques : manger du plancton flottant dans l'eau, gratter puis manger des algues sur les rochers, manger des larves d'insectes du fond, et manger les œufs d'autres espèces de cichlidés. Les cichlidés du lac Victoria sont le produit d'un rayonnement adaptatif. Un rayonnement adaptatif est un rayonnement rapide (moins de trois millions d'années dans le cas des cichlidés du lac Victoria) qui se ramifie par spéciation d'un arbre phylogénétique en de nombreuses espèces étroitement apparentées ; en général, les espèces « rayonnent » dans différents habitats et niches. Les pinsons des Galápagos sont un exemple de rayonnement adaptatif modeste avec 15 espèces. Les cichlidés du lac Victoria sont un exemple de rayonnement adaptatif spectaculaire qui inclut environ 500 espèces.
Au moment où les biologistes ont fait cette découverte, certaines espèces ont commencé à disparaître rapidement. L'une des causes de ce déclin est une espèce de gros poissons introduite dans le lac Victoria par la pêche pour nourrir les habitants du lac. La perche du Nil a été introduite en 1963, mais elle est restée faible jusqu'aux années 1980, date à laquelle ses populations ont commencé à augmenter. La population de perches du Nil a augmenté en consommant des cichlidés, entraînant une espèce après l'autre jusqu'à l'extinction (disparition d'une espèce). En fait, plusieurs facteurs ont joué un rôle dans l'extinction de quelque 200 espèces de cichlidés dans le lac Victoria : la perche du Nil, la baisse de la qualité de l'eau du lac due à l'agriculture et au défrichement des terres sur les rives du lac Victoria et l'augmentation de la pression de pêche. Les scientifiques n'avaient même pas catalogué toutes les espèces présentes ; tant d'entre elles ont disparu qu'elles n'ont jamais été nommées. La diversité est aujourd'hui l'ombre de ce qu'elle était autrefois.
Les cichlidés du lac Victoria sont un aperçu de la perte rapide d'espèces qui se produit aujourd'hui partout sur Terre et qui est causée par l'activité humaine. L'extinction est un processus naturel de macroévolution qui se produit au rythme de l'extinction d'environ une espèce sur un million chaque année. Les archives fossiles révèlent qu'il y a eu cinq périodes d'extinction massive dans l'histoire avec des taux de perte d'espèces beaucoup plus élevés, et le taux de perte d'espèces aujourd'hui est comparable à ces périodes d'extinction massive. Cependant, il existe une différence majeure entre les anciennes extinctions massives et l'extinction actuelle que nous connaissons : l'activité humaine. Plus précisément, trois activités humaines ont un impact majeur : la destruction de l'habitat, l'introduction d'espèces exotiques et la surexploitation. Les prévisions de perte d'espèces au cours du prochain siècle, soit un laps de temps minuscule sur des échelles de temps géologiques, vont de 10 % à 50 %. Des extinctions de cette ampleur ne se sont produites que cinq autres fois dans l'histoire de la planète, et elles ont été causées par des événements cataclysmiques qui ont changé le cours de l'histoire de la vie dans chaque cas. La Terre est aujourd'hui dans l'une de ces époques.
Types de biodiversité
Les scientifiques reconnaissent généralement que le terme biodiversité décrit le nombre et les types d'espèces présentes dans un lieu ou sur la planète. Les espèces peuvent être difficiles à définir, mais la plupart des biologistes se sentent toujours à l'aise avec le concept et sont capables d'identifier et de dénombrer les espèces eucaryotes dans la plupart des contextes. Les biologistes ont également identifié d'autres mesures de la biodiversité, dont certaines sont importantes pour planifier la manière de préserver la biodiversité.
La diversité génétique est l'un de ces concepts alternatifs. La diversité ou la variation génétique est la matière première de l'adaptation d'une espèce. Le potentiel d'adaptation futur d'une espèce dépend de la diversité génétique contenue dans les génomes des individus des populations qui la composent. Il en va de même pour les catégories taxonomiques supérieures. Un genre composé de types d'espèces très différents aura une plus grande diversité génétique qu'un genre dont les espèces se ressemblent et ont une écologie similaire. S'il était possible de choisir entre l'un de ces genres d'espèces à préserver, celui qui présente le plus grand potentiel d'évolution ultérieure serait le plus diversifié sur le plan génétique. L'idéal serait de ne pas avoir à faire de tels choix, mais cela peut devenir de plus en plus la norme.
De nombreux gènes codent pour les protéines, qui à leur tour exécutent les processus métaboliques qui permettent aux organismes de vivre et de se reproduire. La diversité génétique peut être mesurée en tant que diversité chimique dans la mesure où différentes espèces produisent une variété de substances chimiques dans leurs cellules, à la fois les protéines et les produits et sous-produits du métabolisme. Cette diversité chimique présente des avantages potentiels pour les humains en tant que source de produits pharmaceutiques. Elle fournit donc un moyen de mesurer la diversité qui est importante pour la santé et le bien-être humains.
Les humains ont créé de la diversité chez les animaux domestiques, les plantes et les champignons. Cette diversité subit également des pertes en raison de la migration, des forces du marché et de la mondialisation croissante de l'agriculture, en particulier dans les régions densément peuplées telles que la Chine, l'Inde et le Japon. La population humaine dépend directement de cette diversité en tant que source de nourriture stable, et son déclin inquiète les biologistes et les agronomes.
Il est également utile de définir la diversité des écosystèmes, c'est-à-dire le nombre d'écosystèmes différents sur la planète ou dans une zone géographique donnée (Figure\(\PageIndex{1}\)). Des écosystèmes entiers peuvent disparaître même si certaines espèces peuvent survivre en s'adaptant à d'autres écosystèmes. La perte d'un écosystème signifie la perte des interactions entre les espèces, la perte des caractéristiques uniques de la coadaptation et la perte de productivité biologique qu'un écosystème est capable de créer. L'écosystème des prairies est un exemple d'écosystème largement éteint en Amérique du Nord. Les prairies s'étendaient autrefois sur le centre de l'Amérique du Nord, depuis la forêt boréale du nord du Canada jusqu'au Mexique. Elles ont pratiquement disparu, remplacées par des champs de culture, des pâturages et des banlieues. De nombreuses espèces survivent, mais l'écosystème extrêmement productif qui était responsable de la création des sols agricoles les plus productifs a maintenant disparu. En conséquence, les sols disparaissent ou doivent être entretenus à plus grands frais.

Diversité des espèces actuelle
Malgré des efforts considérables, les connaissances sur les espèces qui peuplent la planète sont limitées. Une estimation récente suggère que les espèces d'eucaryotes nommées par la science, soit environ 1,5 million d'espèces, représentent moins de 20 pour cent du nombre total d'espèces d'eucaryotes présentes sur la planète (8,7 millions d'espèces, selon une estimation). Les estimations du nombre d'espèces procaryotes ne sont que des suppositions, mais les biologistes s'accordent à dire que la science ne fait que commencer à cataloguer leur diversité. Même avec ce que l'on sait, il n'existe pas de répertoire central des noms ou des échantillons des espèces décrites ; il n'y a donc aucun moyen de s'assurer que les 1,5 million de descriptions sont un chiffre exact. Il s'agit d'une estimation basée sur les opinions d'experts de différents groupes taxonomiques. Étant donné que la Terre perd des espèces à un rythme accéléré, la science n'a rien à voir avec les cichlidés du lac Victoria : elle ne sait que peu de choses sur ce qui est perdu. Le tableau\(\PageIndex{1}\) présente les estimations récentes de la biodiversité dans différents groupes.
Tableau\(\PageIndex{1}\) : Estimations des espèces décrites et prévues par groupe taxonomique
| Mora et coll. 2011 1 | Chapman 2009 2 | Groombridge et Jenkins 2002 3 | ||||
|---|---|---|---|---|---|---|
| Décrit | Prédit | Décrit | Prédit | Décrit | Prédit | |
| Animalia | 1 124 516 | 9 920 000 | 1 424 153 | 6 836 330 | 1 225 500 | 10 820 000 |
| Chromista | 17 892 | 34 900 | 25 044 | 200 500 | — | — |
| Champignons | 44 368 | 616 320 | 98 998 | 1 500 000 | 72 000 | 1 500 000 |
| Plantae | 224 244 | 314 600 | 310 129 | 390 800 | 270 000 | 320 000 |
| Protozoaires | 16 236 | 72 800 | 28 871 | 1 000 000 | 80 000 | 600 000 |
| Procaryotes | — | — | 10 307 | 1 000 000 | 10 175 | — |
| Totale | 1 438 769 | 10 960 000 | 1 897 502 | 10 897 630 | 1 657 675 | 13 240 000 |
Il existe diverses initiatives visant à cataloguer les espèces décrites de manière accessible, et Internet facilite cet effort. Néanmoins, il a été souligné qu'au rythme actuel de description des espèces, qui, selon le rapport sur l'état des espèces observées, est de 17 000 à 20 000 nouvelles espèces par an, il faudra près de 500 ans pour terminer la description de la vie sur cette planète. 4 Au fil du temps, la tâche devient de plus en plus impossible et de plus en plus facile à mesure que l'extinction élimine des espèces de la planète.
Nommer et dénombrer les espèces peut sembler une activité sans importance compte tenu des autres besoins de l'humanité, mais il ne s'agit pas simplement d'une comptabilité. La description d'une espèce est un processus complexe qui permet aux biologistes de déterminer les caractéristiques uniques d'un organisme et de déterminer si cet organisme appartient ou non à une autre espèce décrite. Il permet aux biologistes de trouver et de reconnaître l'espèce après la découverte initiale, et leur permet de suivre les questions concernant sa biologie. De plus, les caractéristiques uniques de chaque espèce la rendent potentiellement précieuse pour les humains ou pour d'autres espèces dont dépendent les humains. Il est important de comprendre ces caractéristiques pour trouver et nommer des espèces.
Modèles de biodiversité
La biodiversité n'est pas uniformément répartie sur Terre. Le lac Victoria contenait à lui seul près de 500 espèces de cichlidés, sans tenir compte des autres familles de poissons présentes dans le lac. Toutes ces espèces n'étaient présentes que dans le lac Victoria ; par conséquent, les 500 espèces de cichlidés étaient endémiques. Les espèces endémiques ne se trouvent que dans un seul endroit. Les espèces endémiques dont l'aire de répartition est très limitée sont particulièrement vulnérables à l'extinction. Les niveaux taxonomiques supérieurs, tels que les genres et les familles, peuvent également être endémiques. Le lac Huron abrite environ 79 espèces de poissons, que l'on trouve toutes dans de nombreux autres lacs d'Amérique du Nord. Qu'est-ce qui explique la différence de diversité des poissons dans ces deux lacs ? Le lac Victoria est un lac tropical, tandis que le lac Huron est un lac tempéré. Le lac Huron dans sa forme actuelle n'a que 7 000 ans environ, tandis que le lac Victoria dans sa forme actuelle a environ 15 000 ans. Les biogéographes ont suggéré que ces deux facteurs, la latitude et l'âge, sont deux des nombreuses hypothèses permettant d'expliquer les modèles de biodiversité sur la planète.
Career Connection : biogéographe
La biogéographie est l'étude de la distribution des espèces du monde, à la fois dans le passé et dans le présent. Le travail des biogéographes est essentiel pour comprendre notre environnement physique, comment l'environnement affecte les espèces et comment les changements environnementaux influent sur la distribution d'une espèce ; il a également joué un rôle essentiel dans le développement de la théorie de l'évolution. Les biogéographes doivent comprendre à la fois la biologie et l'écologie. Ils doivent également bien connaître les études évolutionnistes, la science des sols et la climatologie.
La biogéographie comporte trois principaux domaines d'étude : la biogéographie écologique, la biogéographie historique (appelée paléobiogéographie) et la biogéographie de conservation. La biogéographie écologique étudie les facteurs actuels qui influent sur la distribution des plantes et des animaux. La biogéographie historique, comme son nom l'indique, étudie la distribution passée des espèces. La biogéographie de conservation, quant à elle, est axée sur la protection et la restauration des espèces sur la base d'informations écologiques historiques et actuelles connues. Chacun de ces domaines prend en compte à la fois la zoogéographie et la phytogéographie, c'est-à-dire la distribution passée et présente des animaux et des plantes.
L'une des plus anciennes tendances observées en écologie est que la biodiversité des espèces de presque tous les groupes taxonomiques augmente à mesure que la latitude diminue. En d'autres termes, la biodiversité augmente plus près de l'équateur (Figure\(\PageIndex{2}\)).
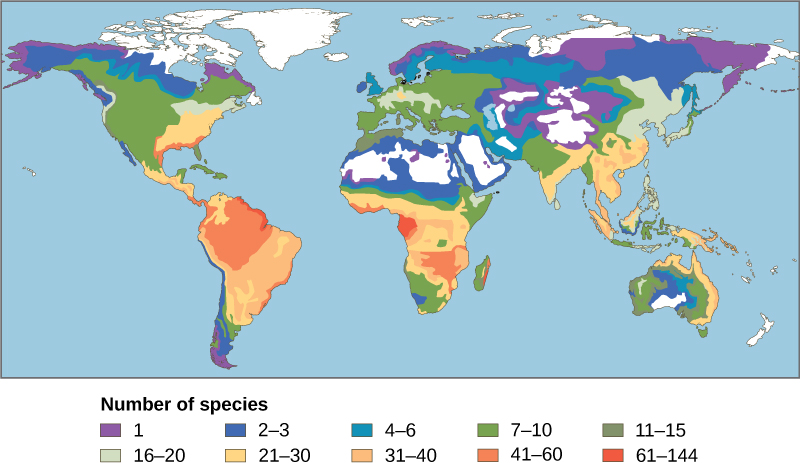
On ne sait pas encore pourquoi la biodiversité augmente à proximité de l'équateur, mais les hypothèses incluent un vieillissement plus avancé des écosystèmes des régions tropicales par rapport aux régions tempérées qui étaient largement dépourvues de vie ou considérablement appauvries lors de la dernière glaciation. L'idée est qu'un âge plus avancé donne plus de temps pour la spéciation. Une autre explication possible est l'augmentation de l'énergie que les tropiques reçoivent du soleil par rapport à la diminution de l'énergie que reçoivent les régions tempérées et polaires. On ne sait pas exactement comment un apport énergétique accru pourrait se traduire par un plus grand nombre d'espèces. La complexité des écosystèmes tropicaux peut favoriser la spéciation en augmentant l'hétérogénéité, ou le nombre de niches écologiques, dans les tropiques par rapport aux latitudes plus élevées. L'hétérogénéité accrue offre davantage de possibilités de coévolution, de spécialisation et peut-être de pressions de sélection plus importantes conduisant à la différenciation des populations. Cependant, cette hypothèse souffre d'une certaine circularité : les écosystèmes comptant un plus grand nombre d'espèces encouragent la spéciation, mais comment ont-ils réussi à obtenir plus d'espèces au départ ? Les régions tropicales ont été perçues comme étant plus stables que les régions tempérées, qui ont un climat et une saisonnalité de la journée prononcés. Les tropiques ont leurs propres formes de saisonnalité, telles que les précipitations, mais on suppose généralement qu'il s'agit d'environnements plus stables et cette stabilité pourrait favoriser la spéciation.
Quels que soient les mécanismes, il est certainement vrai que tous les niveaux de biodiversité sont les plus élevés sous les tropiques. De plus, le taux d'endémisme est le plus élevé et il existe de plus en plus de points chauds de biodiversité. Cependant, cette richesse de diversité signifie également que les connaissances sur les espèces sont les plus faibles et qu'il existe un fort potentiel de perte de biodiversité.
Conservation de la biodiversité
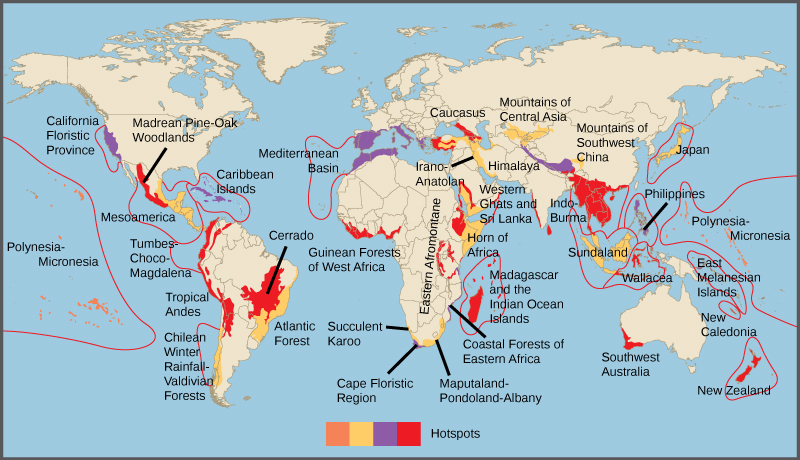
En 1988, l'écologiste britannique Norman Myers a développé un concept de conservation pour identifier les zones riches en espèces et présentant un risque important de disparition d'espèces : les points chauds de biodiversité. Les points chauds de biodiversité sont des zones géographiques qui abritent un grand nombre d'espèces endémiques. Le but du concept était d'identifier les sites importants de la planète pour les efforts de conservation, une sorte de triage de conservation. En protégeant les zones sensibles, les gouvernements sont en mesure de protéger un plus grand nombre d'espèces. Les critères initiaux pour désigner un point névralgique incluaient la présence de 1 500 espèces végétales endémiques ou plus et 70 pour cent de la zone perturbée par l'activité humaine. Il existe aujourd'hui 34 points chauds de biodiversité (Figure\(\PageIndex{3}\)) abritant un grand nombre d'espèces endémiques, dont la moitié des plantes endémiques de la Terre.
Modification de la biodiversité au fil du temps
Le nombre d'espèces présentes sur la planète, ou dans n'importe quelle zone géographique, est le résultat d'un équilibre entre deux processus évolutifs en cours : la spéciation et l'extinction. Les deux sont des processus naturels de « naissance » et de « mort » de la macroévolution. Lorsque les taux de spéciation commenceront à dépasser les taux d'extinction, le nombre d'espèces augmentera ; de même, le nombre d'espèces diminuera lorsque les taux d'extinction commenceront à dépasser les taux de spéciation. Tout au long de l'histoire de la Terre, ces deux processus ont fluctué, entraînant parfois des changements spectaculaires dans le nombre d'espèces présentes sur Terre, comme en témoignent les archives fossiles (Figure\(\PageIndex{4}\)).
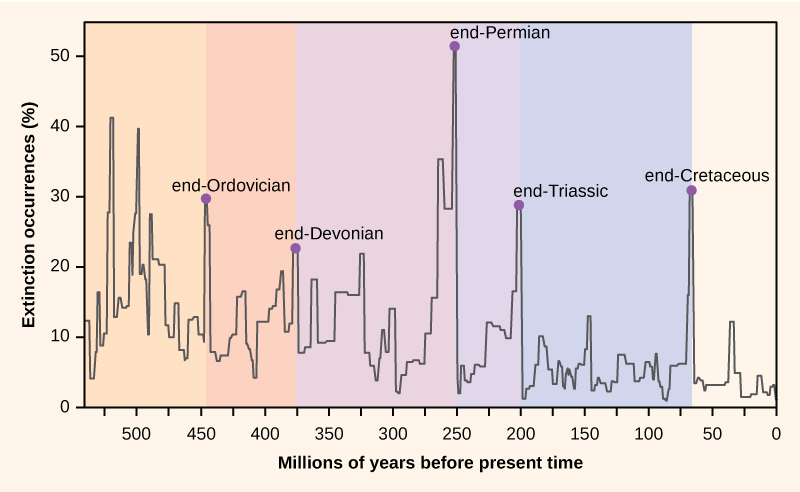
Les paléontologues ont identifié cinq strates dans les archives fossiles qui semblent présenter des pertes de biodiversité soudaines et spectaculaires (plus de la moitié de toutes les espèces existantes disparaissent des archives fossiles). C'est ce qu'on appelle des extinctions massives. Il existe de nombreux événements d'extinction de moindre importance, mais tout de même dramatiques, mais ce sont les cinq extinctions massives qui ont suscité le plus de recherches. On peut faire valoir que les cinq extinctions massives ne sont que les cinq événements les plus extrêmes d'une série continue de grandes extinctions à travers le Phanérozoïque (depuis 542 millions d'années). Dans la plupart des cas, les causes hypothétiques restent controversées ; toutefois, l'événement le plus récent semble clair.
Les cinq extinctions de masse
Les archives fossiles des extinctions massives ont servi de base à la définition des périodes de l'histoire géologique, de sorte qu'elles se produisent généralement au point de transition entre les périodes géologiques. La transition des fossiles d'une période à l'autre reflète la perte dramatique d'espèces et l'apparition progressive de nouvelles espèces. Ces transitions sont visibles dans les couches rocheuses. Le tableau\(\PageIndex{2}\) fournit des données sur les cinq extinctions massives.
| Période géologique | Nom de l'extinction massive | Temps (il y a des millions d'années) |
|---|---|---|
| Ordovicien-Silurien | Ordovicien final O-S | 450—440 |
| Dévonien tardif | Fin dévonien | 375-360 |
| Permien-Trias | Fin du Permien | 251 |
| Triasque-Jurassique | Fin du Trias | 205 |
| Crétacéo-Paléogène | Crétacé final K—Pg (K—T) | 65,5 |
L'extinction ordovicienne-silurienne est la première extinction de masse enregistrée et la deuxième plus importante. Au cours de cette période, environ 85 pour cent des espèces marines (peu d'espèces vivaient en dehors des océans) ont disparu. L'hypothèse principale de sa cause est une période de glaciation puis de réchauffement. L'événement d'extinction consiste en fait en deux événements d'extinction séparés par environ 1 million d'années. Le premier événement a été causé par le refroidissement et le second par le réchauffement qui a suivi. Les changements climatiques ont affecté les températures et le niveau de la mer. Certains chercheurs ont suggéré qu'un sursaut gamma, provoqué par une supernova voisine, pourrait être à l'origine de l'extinction de l'Ordovicien-Silurien. L'explosion de rayons gamma aurait détruit la couche d'ozone de la Terre, provoquant un intense rayonnement ultraviolet provenant du soleil et pourrait expliquer les changements climatiques observés à l'époque. L'hypothèse est spéculative, mais les influences extraterrestres sur l'histoire de la Terre constituent un axe de recherche actif. Le rétablissement de la biodiversité après l'extinction massive a pris de 5 à 20 millions d'années, selon le lieu.
L'extinction du Dévonien tardif s'est peut-être produite sur une période relativement longue. Il semble avoir affecté les espèces marines et non les plantes ou les animaux vivant dans les habitats terrestres. Les causes de cette extinction sont mal connues.
L'extinction de la fin du Permien a été la plus importante de l'histoire de la vie. En effet, on pourrait faire valoir que la Terre est presque devenue vide de vie lors de cet événement d'extinction. La planète était très différente avant et après cet événement. On estime que 96 pour cent de toutes les espèces marines et 70 pour cent de toutes les espèces terrestres ont disparu. C'est à cette époque, par exemple, que les trilobites, un groupe qui a survécu à l'extinction de l'Ordovicien-Silurien, ont disparu. Les causes de cette extinction massive ne sont pas claires, mais le principal suspect est l'activité volcanique étendue et généralisée qui a entraîné un réchauffement planétaire incontrôlable. Les océans sont devenus largement anoxiques, étouffant la vie marine. La diversité des tétrapodes terrestres a mis 30 millions d'années à se rétablir après l'extinction de la fin du Permien. L'extinction du Permien a radicalement modifié la composition de la biodiversité de la Terre et le cours de l'évolution.
Les causes de l'extinction du Triasque-Jurassique ne sont pas claires et des hypothèses de changement climatique, d'impact d'astéroïdes et d'éruptions volcaniques ont été avancées. L'extinction s'est produite juste avant l'éclatement du supercontinent Pangée, bien que des études récentes suggèrent que les extinctions se sont peut-être produites plus progressivement tout au long du Trias.
Les causes de l'extinction de la fin du Crétacé sont celles qui sont les mieux comprises. C'est lors de cette extinction, il y a environ 65 millions d'années, que les dinosaures, le groupe de vertébrés dominant depuis des millions d'années, ont disparu de la planète (à l'exception d'un clade de théropodes qui a donné naissance à des oiseaux). En effet, chaque animal terrestre pesant plus de 25 kg a disparu. La cause de cette extinction est désormais considérée comme le résultat de l'impact cataclysmique d'une grosse météorite, ou astéroïde, au large de ce qui est aujourd'hui la péninsule du Yucatán. Cette hypothèse, proposée pour la première fois en 1980, était une explication radicale basée sur une forte augmentation des niveaux d'iridium (qui pleut de l'espace sous forme de météores à un rythme assez constant mais qui est par ailleurs absent de la surface de la Terre) dans la couche rocheuse qui marque la limite entre le Crétacé et le Paléogène (Figurine\(\PageIndex{5}\)). Cette limite a marqué la disparition des dinosaures dans les fossiles ainsi que dans de nombreux autres taxons. Les chercheurs qui ont découvert le pic d'iridium l'ont interprété comme un afflux rapide d'iridium de l'espace vers l'atmosphère (sous la forme d'un gros astéroïde) plutôt que comme un ralentissement du dépôt de sédiments pendant cette période. C'était une explication radicale, mais le rapport faisant état d'un cratère d'impact vieilli et dimensionné de manière appropriée en 1991 a rendu l'hypothèse plus crédible. Aujourd'hui, de nombreuses preuves géologiques soutiennent cette théorie. Les temps de rétablissement de la biodiversité après l'extinction de la fin du Crétacé sont plus courts, en termes géologiques, que ceux de la fin du Permien, de l'ordre de 10 millions d'années.
Art Connection

Les scientifiques ont mesuré l'abondance relative des spores de fougères au-dessus et au-dessous de la limite K—Pg dans cet échantillon de roche. Parmi les affirmations suivantes, laquelle correspond le plus probablement à leurs conclusions ?
- Une abondance de spores de fougères provenant de plusieurs espèces a été trouvée en dessous de la limite K—Pg, mais aucune n'a été trouvée au-dessus.
- Une abondance de spores de fougères provenant de plusieurs espèces a été observée au-dessus de la limite K—Pg, mais aucune n'a été trouvée en dessous.
- Une abondance de spores de fougères a été observée à la fois au-dessus et en dessous de la limite K—Pg, mais une seule espèce a été trouvée en dessous de la limite et de nombreuses espèces ont été trouvées au-dessus de la limite.
- De nombreuses espèces de spores de fougères ont été trouvées à la fois au-dessus et en dessous de la limite, mais le nombre total de spores était plus élevé en dessous de la limite.

L'extinction du Pléistocène
L'extinction du Pléistocène est l'une des plus petites extinctions, et elle est récente. Il est bien connu que l'Amérique du Nord et, dans une certaine mesure, les animaux eurasiens, la mégafaune ou les grands animaux ont disparu vers la fin de la dernière période glaciaire. L'extinction semble s'être produite au cours d'une période relativement limitée, il y a 10 000 à 12 000 ans. En Amérique du Nord, les pertes ont été considérables, notamment les mammouths laineux (dont la dernière datation remonte à environ 4 000 ans dans une population isolée), le mastodonte, les castors géants, les paresseux géants, les chats à dents de sabre et le chameau d'Amérique du Nord, pour n'en nommer que quelques-uns. La possibilité que l'extinction rapide de ces grands animaux soit due à une chasse excessive a été suggérée pour la première fois dans les années 1900. Les recherches sur cette hypothèse se poursuivent aujourd'hui. Il semble probable que la chasse excessive ait causé de nombreuses extinctions historiques préécrites dans de nombreuses régions du monde.
En général, le moment des extinctions du Pléistocène était en corrélation avec l'arrivée des humains et non avec les événements liés au changement climatique, qui constitue la principale hypothèse concurrente à l'origine de ces extinctions. Les extinctions ont commencé en Australie il y a environ 40 000 à 50 000 ans, juste après l'arrivée des humains dans la région : un lion marsupial, un wombat géant d'une tonne et plusieurs espèces de kangourous géants ont disparu. En Amérique du Nord, l'extinction de presque tous les grands mammifères s'est produite il y a 10 000 à 12 000 ans. Il ne reste que les petits mammifères tels que les ours, les wapitis, les orignaux et les couguars. Enfin, sur de nombreuses îles océaniques éloignées, l'extinction de nombreuses espèces s'est produite en même temps que l'arrivée des humains. Toutes les îles n'avaient pas de gros animaux, mais lorsqu'il y avait de gros animaux, ils étaient perdus. Madagascar a été colonisée il y a environ 2 000 ans et les grands mammifères qui y vivaient ont disparu. L'Eurasie et l'Afrique ne présentent pas ce schéma, mais elles n'ont pas non plus connu d'arrivée récente d'humains. Les humains sont arrivés en Eurasie il y a des centaines de milliers d'années après l'origine de l'espèce en Afrique. Ce sujet continue de faire l'objet de recherches et d'hypothèses actives. Il semble clair que même si le climat a joué un rôle, dans la plupart des cas, la chasse humaine a précipité les extinctions.
Extinctions actuelles
La sixième extinction massive, ou Holocène, semble avoir commencé plus tôt qu'on ne le pensait auparavant et est principalement liée aux activités de l'Homo sapiens. Depuis le début de l'Holocène, de nombreuses extinctions récentes d'espèces individuelles sont enregistrées dans des écrits humains. La plupart d'entre elles coïncident avec l'expansion des colonies européennes depuis les années 1500.
L'un des exemples les plus anciens et les plus connus est l'oiseau dodo. Le dodo vivait dans les forêts de Maurice, une île de l'océan Indien. L'oiseau dodo s'est éteint vers 1662. Il était chassé pour sa viande par les marins et était une proie facile car le dodo, qui n'évoluait pas avec les humains, s'approchait des gens sans crainte. Des porcs, des rats et des chiens introduits sur l'île par des navires européens ont également tué des jeunes dodo et des œufs.
La vache de mer de Steller a disparu en 1768 ; elle était apparentée au lamantin et vivait probablement autrefois le long de la côte nord-ouest de l'Amérique du Nord. La vache de mer de Steller a été découverte pour la première fois par les Européens en 1741 et était chassée pour sa viande et son huile. La dernière vache de mer a été tuée en 1768. Cela représente 27 ans entre le premier contact de la vache de mer avec les Européens et l'extinction de l'espèce.
En 1914, le dernier pigeon passager vivant est mort dans un zoo de Cincinnati, dans l'Ohio. Cette espèce avait autrefois assombri le ciel de l'Amérique du Nord lors de ses migrations, mais elle a été chassée et a souffert de la perte d'habitat due au défrichement des forêts pour les terres agricoles. En 1918, la dernière perruche de Caroline vivante est morte en captivité. Cette espèce était autrefois commune dans l'est des États-Unis, mais elle a souffert de la perte d'habitat. L'espèce a également été chassée parce qu'elle mangeait des fruits du verger lorsque ses aliments indigènes ont été détruits pour faire place à des terres agricoles. L'otarie japonaise, qui habitait une vaste zone autour du Japon et de la côte de la Corée, a disparu dans les années 1950 grâce aux pêcheurs. Le phoque moine des Caraïbes était répandu dans toute la mer des Caraïbes mais a été détruit par la chasse en 1952.
Ce ne sont là que quelques-unes des extinctions enregistrées au cours des 500 dernières années. L'Union internationale pour la conservation de la nature (UICN) tient une liste d'espèces éteintes et menacées appelée Liste rouge. La liste n'est pas complète, mais elle décrit 380 espèces de vertébrés disparues après 1500 après Jésus-Christ, dont 86 ont été chassées par la surchasse ou la surpêche.
Estimations des taux d'extinction actuels
Les estimations des taux d'extinction sont entravées par le fait que la plupart des extinctions se produisent probablement sans observation. L'extinction d'un oiseau ou d'un mammifère est susceptible d'être remarquée par les humains, en particulier s'il a été chassé ou utilisé d'une autre manière. Mais il existe de nombreux organismes qui présentent moins d'intérêt pour les humains (pas nécessairement de moindre valeur) et beaucoup ne sont pas décrits.
Le taux d'extinction de fond est estimé à environ une par million d'espèces par an (E/MSY). Par exemple, en supposant qu'il existe environ dix millions d'espèces, on s'attend à ce que dix espèces disparaissent chaque année (chaque année représente dix millions d'espèces par an).
Une estimation du taux d'extinction contemporain utilise les extinctions enregistrées depuis 1500. Pour les oiseaux uniquement, cette méthode donne une estimation de 26 E/MSY. Toutefois, cette valeur peut être sous-estimée pour trois raisons. Tout d'abord, de nombreuses espèces n'auraient été décrites que bien plus tard dans la période, de sorte que leur disparition serait passée inaperçue. Deuxièmement, le nombre d'espèces récemment éteintes augmente parce que des espèces éteintes sont maintenant décrites à partir de restes squelettiques. Enfin, certaines espèces sont probablement déjà éteintes, même si les défenseurs de l'environnement hésitent à les nommer ainsi. La prise en compte de ces facteurs augmente le taux d'extinction estimé de près de 100 E/MSY. Le taux prévu d'ici la fin du siècle est de 1 500 E/MSY.
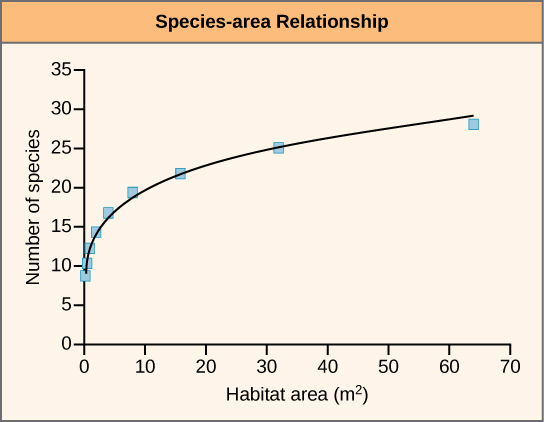
Une deuxième approche pour estimer les taux d'extinction actuels consiste à corréler la perte d'espèces à la perte d'habitat en mesurant la perte de superficie forestière et en comprenant les relations entre les espèces et la superficie. La relation entre l'espèce et la zone est la vitesse à laquelle de nouvelles espèces sont observées lorsque la superficie étudiée est augmentée. Des études ont montré que le nombre d'espèces présentes augmente à mesure que la taille de l'île augmente. Il a également été démontré que ce phénomène est également vrai dans d'autres habitats. En inversant cette relation, si la superficie de l'habitat est réduite, le nombre d'espèces qui y vivent diminuera également. Les estimations des taux d'extinction basées sur la perte d'habitat et les relations entre les espèces et les zones suggèrent qu'avec une perte d'habitat d'environ 90 pour cent, on s'attend à ce que 50 pour cent des espèces disparaissent. Les estimations de la superficie des espèces ont permis de calculer le taux d'extinction des espèces d'environ 1 000 E/MSY et plus. En général, les observations réelles ne montrent pas une telle ampleur de pertes et des suggestions ont été avancées selon lesquelles il y aurait un retard dans l'extinction. Des travaux récents ont également remis en question l'applicabilité de la relation entre l'espèce et la zone lors de l'estimation de la perte d'espèces. Ce travail soutient que la relation entre l'espèce et la zone conduit à une surestimation des taux d'extinction. Une meilleure relation d'utilisation peut être la relation entre les zones endémiques. L'utilisation de cette méthode ramènerait les estimations à environ 500 E/MSY au cours du siècle à venir. Notez que cette valeur est toujours 500 fois la fréquence de fond.
Lien vers l'apprentissage

Découvrez cette exploration interactive des espèces menacées et éteintes, de leurs écosystèmes et des causes de leur mise en danger ou de leur extinction.
Résumé
La biodiversité existe à de multiples niveaux de l'organisation et est mesurée de différentes manières en fonction des objectifs des personnes qui prennent les mesures. Ces mesures incluent le nombre d'espèces, la diversité génétique, la diversité chimique et la diversité des écosystèmes. Le nombre d'espèces décrites est estimé à 1,5 million et environ 17 000 nouvelles espèces sont décrites chaque année. Les estimations du nombre total d'espèces sur Terre varient mais sont de l'ordre de 10 millions. La biodiversité est négativement corrélée à la latitude pour la plupart des taxons, ce qui signifie que la biodiversité est plus élevée sous les tropiques. Le mécanisme de cette tendance n'est pas connu avec certitude, mais plusieurs hypothèses plausibles ont été avancées.
Cinq extinctions massives entraînant la perte de plus de 50 pour cent des espèces existantes sont observables dans les archives fossiles. Les temps de rétablissement de la biodiversité après des extinctions massives varient, mais peuvent atteindre 30 millions d'années. Les extinctions récentes sont enregistrées dans l'histoire écrite et constituent la base d'une méthode d'estimation des taux d'extinction contemporains. L'autre méthode utilise des mesures de la perte d'habitat et des relations entre espèces et zones. Les estimations des taux d'extinction contemporains varient, mais certains taux sont 500 fois supérieurs au taux de fond, tel que déterminé à partir des archives fossiles, et devraient augmenter.
Connexions artistiques
Figurine\(\PageIndex{5}\): Scientists measured the relative abundance of fern spores above and below the K-Pg boundary in this rock sample. Which of the following statements most likely represents their findings?
- An abundance of fern spores from several species was found below the K-Pg boundary, but none was found above.
- An abundance of fern spores from several species was found above the K-Pg boundary, but none was found below.
- An abundance of fern spores was found both above and below the K-Pg boundary, but only one species was found below the boundary , and many species were found above the boundary.
- Many species of fern spores were found both above and below the boundary, but the total number of spores was greater below the boundary.
- Answer
-
A. An abundance of fern spores from several species was found below the K-Pg boundary, but none was found above.
Footnotes
- 1 Mora Camilo et al., “How Many Species Are There on Earth and in the Ocean?” PLoS Biology (2011), doi:10.1371/journal.pbio.1001127.
- 2 Arthur D. Chapman, Numbers of Living Species in Australia and the World, 2nd ed. (Canberra, AU: Australian Biological Resources Study, 2009). www.environment.gov.au/biodiv...d-complete.pdf.
- 3 Brian Groombridge and Martin D. Jenkins. World Atlas of Biodiversity: Earth’s Living Resources in the 21st Century. Berkeley: University of California Press, 2002.
- 4 International Institute for Species Exploration (IISE), 2011 State of Observed Species (SOS). Tempe, AZ: IISE, 2011. Accessed May, 20, 2012. species.asu.edu/SOS.
Glossary
- adaptive radiation
- rapid branching through speciation of a phylogenetic tree into many closely related species
- biodiversity
- variety of a biological system, typically conceived as the number of species, but also applying to genes, biochemistry, and ecosystems
- biodiversity hotspot
- concept originated by Norman Myers to describe a geographical region with a large number of endemic species and a large percentage of degraded habitat
- chemical diversity
- variety of metabolic compounds in an ecosystem
- ecosystem diversity
- variety of ecosystems
- endemic species
- species native to one place
- extinction
- disappearance of a species from Earth; local extinction is the disappearance of a species from a region
- extinction rate
- number of species becoming extinct over time, sometimes defined as extinctions per million species–years to make numbers manageable (E/MSY)
- genetic diversity
- variety of genes in a species or other taxonomic group or ecosystem, the term can refer to allelic diversity or genome-wide diversity
- heterogeneity
- number of ecological niches
- megafauna
- large animals
- species-area relationship
- relationship between area surveyed and number of species encountered; typically measured by incrementally increasing the area of a survey and determining the cumulative numbers of species