
46.1 : Écologie des écosystèmes
- Page ID
- 189712

Compétences à développer
- Décrire les principaux types d'écosystèmes de la Terre
- Expliquer les méthodes utilisées par les écologistes pour étudier la structure et la dynamique des écosystèmes
- Identifier les différentes méthodes de modélisation des écosystèmes
- Faites la différence entre les chaînes alimentaires et les réseaux alimentaires et reconnaissez l'importance de chacun
La vie dans un écosystème repose souvent sur la compétition pour des ressources limitées, une caractéristique de la théorie de la sélection naturelle. La compétition au sein des communautés (tous les êtres vivants dans des habitats spécifiques) est observée à la fois au sein des espèces et entre différentes espèces. Les ressources pour lesquelles les organismes se font concurrence comprennent les matières organiques provenant d'organismes vivants ou anciens, la lumière du soleil et les nutriments minéraux, qui fournissent l'énergie nécessaire aux processus vivants et la matière nécessaire à la constitution des structures physiques des organismes. Les autres facteurs critiques qui influencent la dynamique de la communauté sont les composantes de son environnement physique et géographique : latitude de l'habitat, quantité de précipitations, topographie (altitude) et espèces disponibles. Ce sont toutes des variables environnementales importantes qui déterminent quels organismes peuvent exister dans une zone donnée.
Un écosystème est une communauté d'organismes vivants et de leurs interactions avec leur environnement abiotique (non vivant). Les écosystèmes peuvent être petits, comme les bassins de marée situés près des rives rocheuses de nombreux océans, ou grands, comme la forêt amazonienne au Brésil (Figure\(\PageIndex{1}\)).

Il existe trois grandes catégories d'écosystèmes en fonction de leur environnement général : l'eau douce, l'eau de mer et les écosystèmes terrestres. Ces grandes catégories comprennent des types d'écosystèmes individuels basés sur les organismes présents et le type d'habitat environnemental.
Les écosystèmes océaniques sont les plus courants, représentant 75 % de la surface de la Terre et se répartissent en trois types de base : les océans peu profonds, les eaux profondes de l'océan et les surfaces océaniques profondes (les zones de faible profondeur des océans profonds). Les écosystèmes océaniques peu profonds comprennent des écosystèmes de récifs coralliens extrêmement riches en biodiversité, et la surface profonde de l'océan est connue pour la grande quantité de plancton et de krill (petits crustacés) qui la soutiennent. Ces deux environnements sont particulièrement importants pour les respirateurs aérobies du monde entier, car le phytoplancton est responsable de 40 % de toutes les photosynthèses sur Terre. Bien qu'ils ne soient pas aussi diversifiés que les deux autres, les écosystèmes océaniques profonds abritent une grande variété d'organismes marins. De tels écosystèmes existent même au fond de l'océan, où la lumière ne peut pas pénétrer dans l'eau.
Les écosystèmes d'eau douce sont les plus rares, ne couvrant que 1,8 % de la surface de la Terre. Les lacs, les rivières, les ruisseaux et les sources constituent ces systèmes ; ils sont très diversifiés et abritent une variété de poissons, d'amphibiens, de reptiles, d'insectes, de phytoplancton, de champignons et de bactéries.
Les écosystèmes terrestres, également connus pour leur diversité, sont regroupés en grandes catégories appelées biomes, telles que les forêts tropicales humides, les savanes, les déserts, les forêts de conifères, les forêts de feuillus et la toundra. Le fait de regrouper ces écosystèmes en quelques catégories de biomes masque la grande diversité des écosystèmes individuels qui les composent. Par exemple, la végétation du désert est très variée : les cactus saguaro et les autres plantes du désert de Sonora, aux États-Unis, sont relativement abondants par rapport au désert rocheux désolé de Boa Vista, une île au large des côtes de l'Afrique de l'Ouest (Figure\(\PageIndex{2}\)).

Les écosystèmes sont complexes et comportent de nombreuses parties interactives. Ils sont régulièrement exposés à diverses perturbations ou modifications de l'environnement qui influent sur leur composition : variations annuelles des précipitations et de la température et ralentissement du processus de croissance des plantes, qui peut prendre plusieurs années. Nombre de ces perturbations sont le résultat de processus naturels. Par exemple, lorsque la foudre provoque un incendie de forêt et détruit une partie d'un écosystème forestier, le sol est finalement peuplé de graminées, puis de buissons et d'arbustes, et plus tard d'arbres matures, redonnant à la forêt son état antérieur. L'impact des perturbations environnementales causées par les activités humaines est aussi important que les changements provoqués par les processus naturels. Les pratiques agricoles humaines, la pollution de l'air, les pluies acides, la déforestation mondiale, la surpêche, l'eutrophisation, les déversements de pétrole et les déversements illégaux sur terre et dans l'océan sont autant de sujets de préoccupation pour les défenseurs de l'environnement.
L'équilibre est l'état stable d'un écosystème où tous les organismes sont en équilibre avec leur environnement et entre eux. En écologie, deux paramètres sont utilisés pour mesurer les changements dans les écosystèmes : la résistance et la résilience. La capacité d'un écosystème à maintenir son équilibre malgré les perturbations est appelée résistance. La vitesse à laquelle un écosystème retrouve son équilibre après avoir été perturbé, appelée résilience. La résistance et la résilience des écosystèmes sont particulièrement importantes lorsque l'on considère l'impact humain. La nature d'un écosystème peut changer à tel point qu'il peut perdre totalement sa résilience. Ce processus peut mener à la destruction complète ou à la modification irréversible de l'écosystème.
Chaînes alimentaires et réseaux alimentaires
Le terme « chaîne alimentaire » est parfois utilisé métaphoriquement pour décrire des situations sociales humaines. En ce sens, les chaînes alimentaires sont considérées comme une compétition pour la survie, comme « qui mange qui ? » Quelqu'un mange et quelqu'un est mangé. Il n'est donc pas surprenant que, dans notre société compétitive où les chiens sont mangeurs de chiens, les personnes considérées comme prospères soient considérées comme se situant au sommet de la chaîne alimentaire, consommant toutes les autres pour leur bénéfice, tandis que les moins prospères sont considérées comme se situant au bas de la chaîne.
La compréhension scientifique d'une chaîne alimentaire est plus précise que celle de son utilisation quotidienne. En écologie, une chaîne alimentaire est une séquence linéaire d'organismes par laquelle passent les nutriments et l'énergie : les producteurs primaires, les consommateurs primaires et les consommateurs de niveau supérieur sont utilisés pour décrire la structure et la dynamique de l'écosystème. Il y a un seul chemin à travers la chaîne. Chaque organisme d'une chaîne alimentaire occupe ce que l'on appelle un niveau trophique. En fonction de leur rôle de producteurs ou de consommateurs, des espèces ou des groupes d'espèces peuvent être affectés à différents niveaux trophiques.
Dans de nombreux écosystèmes, le bas de la chaîne alimentaire est constitué d'organismes photosynthétiques (plantes et/ou phytoplancton), appelés producteurs primaires. Les organismes qui consomment les principaux producteurs sont des herbivores : les principaux consommateurs. Les consommateurs secondaires sont généralement des carnivores qui mangent les principaux consommateurs. Les consommateurs tertiaires sont des carnivores qui mangent d'autres carnivores. Les consommateurs de haut niveau se nourrissent des niveaux trophiques inférieurs suivants, et ainsi de suite, jusqu'aux organismes situés au sommet de la chaîne alimentaire : les consommateurs les plus élevés. Dans la chaîne alimentaire du lac Ontario illustrée à la figure\(\PageIndex{3}\), le saumon quinnat est le principal consommateur au sommet de cette chaîne alimentaire.

L'énergie est l'un des principaux facteurs qui limitent la longueur des chaînes alimentaires. L'énergie est perdue sous forme de chaleur entre chaque niveau trophique en raison de la deuxième loi de la thermodynamique. Ainsi, après un nombre limité de transferts d'énergie trophique, la quantité d'énergie restant dans la chaîne alimentaire peut ne pas être suffisante pour soutenir des populations viables à un niveau trophique encore plus élevé.
La perte d'énergie entre les niveaux trophiques est illustrée par les études pionnières de Howard T. Odum dans l'écosystème de Silver Springs, en Floride, dans les années 1940 (Figure\(\PageIndex{4}\)). Les producteurs primaires produisaient 20 819 kcal/m 2 /an (kilocalories par mètre carré et par an), les consommateurs primaires 3 368 kcal/m 2 /an, les consommateurs secondaires 383 kcal/m 2 /an et les consommateurs tertiaires seulement 21 kcal/m 2 /an. Il reste donc peu d'énergie pour un autre niveau de consommateurs dans cet écosystème.
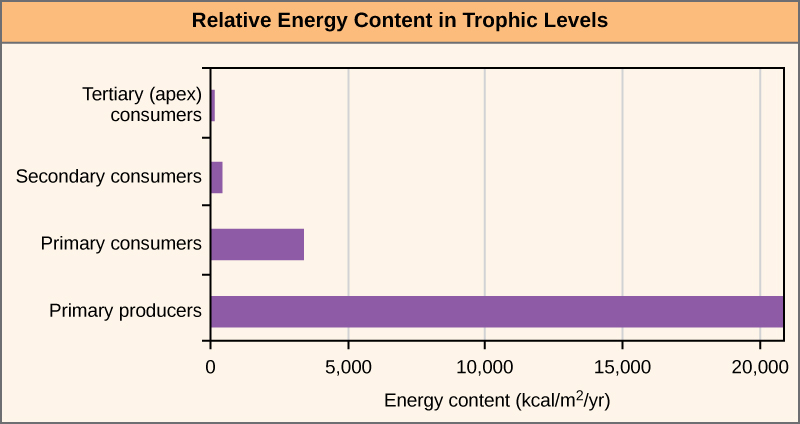
L'utilisation des chaînes alimentaires pour décrire avec précision la plupart des écosystèmes pose un problème. Même lorsque tous les organismes sont regroupés dans les niveaux trophiques appropriés, certains d'entre eux peuvent se nourrir d'espèces de plus d'un niveau trophique ; de même, certains de ces organismes peuvent être consommés par des espèces de plusieurs niveaux trophiques. En d'autres termes, le modèle linéaire des écosystèmes, la chaîne alimentaire, ne décrit pas complètement la structure de l'écosystème. Un modèle holistique, qui tient compte de toutes les interactions entre les différentes espèces et de leurs relations interconnectées complexes entre elles et avec l'environnement, est un modèle plus précis et plus descriptif pour les écosystèmes. Un réseau alimentaire est une représentation graphique d'un réseau holistique et non linéaire de producteurs primaires, de consommateurs primaires et de consommateurs de haut niveau utilisé pour décrire la structure et la dynamique de l'écosystème (Figure\(\PageIndex{5}\)).

La comparaison des deux types de modèles d'écosystèmes structuraux montre la force des deux. Les chaînes alimentaires sont plus souples pour la modélisation analytique, sont plus faciles à suivre et à expérimenter, tandis que les modèles de réseaux alimentaires représentent plus précisément la structure et la dynamique des écosystèmes, et les données peuvent être directement utilisées comme entrées pour la modélisation de simulation.
Lien vers l'apprentissage

Dirigez-vous vers ce simulateur interactif en ligne pour étudier le fonctionnement du réseau alimentaire. Dans la zone Interactive Labs, sous Food Web, cliquez sur Étape 1. Lisez d'abord les instructions, puis cliquez sur Étape 2 pour obtenir des instructions supplémentaires. Lorsque vous êtes prêt à créer une simulation, dans l'angle supérieur droit de la zone Interactive Labs, cliquez sur OUVRIR LE SIMULATEUR.
Deux types généraux de réseaux alimentaires interagissent souvent au sein d'un même écosystème. Un réseau trophique de pâturage (tel que le réseau trophique du lac Ontario sur la figure\(\PageIndex{5}\)) has plants or other photosynthetic organisms at its base, followed by herbivores and various carnivores. A detrital food web consists of a base of organisms that feed on decaying organic matter (dead organisms), called decomposers or detritivores. These organisms are usually bacteria or fungi that recycle organic material back into the biotic part of the ecosystem as they themselves are consumed by other organisms. As all ecosystems require a method to recycle material from dead organisms, most grazing food webs have an associated detrital food web. For example, in a meadow ecosystem, plants may support a grazing food web of different organisms, primary and other levels of consumers, while at the same time supporting a detrital food web of bacteria, fungi, and detrivorous invertebrates feeding off dead plants and animals.
Evolution Connection: Three-spined Stickleback
It is well established by the theory of natural selection that changes in the environment play a major role in the evolution of species within an ecosystem. However, little is known about how the evolution of species within an ecosystem can alter the ecosystem environment. In 2009, Dr. Luke Harmon, from the University of Idaho in Moscow, published a paper that for the first time showed that the evolution of organisms into subspecies can have direct effects on their ecosystem environment.1
The three-spines stickleback (Gasterosteus aculeatus) is a freshwater fish that evolved from a saltwater fish to live in freshwater lakes about 10,000 years ago, which is considered a recent development in evolutionary time (Figure \(\PageIndex{6}\)). Over the last 10,000 years, these freshwater fish then became isolated from each other in different lakes. Depending on which lake population was studied, findings showed that these sticklebacks then either remained as one species or evolved into two species. The divergence of species was made possible by their use of different areas of the pond for feeding called micro niches.
Dr. Harmon and his team created artificial pond microcosms in 250-gallon tanks and added muck from freshwater ponds as a source of zooplankton and other invertebrates to sustain the fish. In different experimental tanks they introduced one species of stickleback from either a single-species or double-species lake.
Over time, the team observed that some of the tanks bloomed with algae while others did not. This puzzled the scientists, and they decided to measure the water's dissolved organic carbon (DOC), which consists of mostly large molecules of decaying organic matter that give pond-water its slightly brownish color. It turned out that the water from the tanks with two-species fish contained larger particles of DOC (and hence darker water) than water with single-species fish. This increase in DOC blocked the sunlight and prevented algal blooming. Conversely, the water from the single-species tank contained smaller DOC particles, allowing more sunlight penetration to fuel the algal blooms.
This change in the environment, which is due to the different feeding habits of the stickleback species in each lake type, probably has a great impact on the survival of other species in these ecosystems, especially other photosynthetic organisms. Thus, the study shows that, at least in these ecosystems, the environment and the evolution of populations have reciprocal effects that may now be factored into simulation models.

Research into Ecosystem Dynamics: Ecosystem Experimentation and Modeling
The study of the changes in ecosystem structure caused by changes in the environment (disturbances) or by internal forces is called ecosystem dynamics. Ecosystems are characterized using a variety of research methodologies. Some ecologists study ecosystems using controlled experimental systems, while some study entire ecosystems in their natural state, and others use both approaches.
A holistic ecosystem model attempts to quantify the composition, interaction, and dynamics of entire ecosystems; it is the most representative of the ecosystem in its natural state. A food web is an example of a holistic ecosystem model. However, this type of study is limited by time and expense, as well as the fact that it is neither feasible nor ethical to do experiments on large natural ecosystems. To quantify all different species in an ecosystem and the dynamics in their habitat is difficult, especially when studying large habitats such as the Amazon Rainforest, which covers 1.4 billion acres (5.5 million km2) of the Earth’s surface.
For these reasons, scientists study ecosystems under more controlled conditions. Experimental systems usually involve either partitioning a part of a natural ecosystem that can be used for experiments, termed a mesocosm, or by re-creating an ecosystem entirely in an indoor or outdoor laboratory environment, which is referred to as a microcosm. A major limitation to these approaches is that removing individual organisms from their natural ecosystem or altering a natural ecosystem through partitioning may change the dynamics of the ecosystem. These changes are often due to differences in species numbers and diversity and also to environment alterations caused by partitioning (mesocosm) or re-creating (microcosm) the natural habitat. Thus, these types of experiments are not totally predictive of changes that would occur in the ecosystem from which they were gathered.
As both of these approaches have their limitations, some ecologists suggest that results from these experimental systems should be used only in conjunction with holistic ecosystem studies to obtain the most representative data about ecosystem structure, function, and dynamics.
Scientists use the data generated by these experimental studies to develop ecosystem models that demonstrate the structure and dynamics of ecosystems. Three basic types of ecosystem modeling are routinely used in research and ecosystem management: a conceptual model, an analytical model, and a simulation model. A conceptual model is an ecosystem model that consists of flow charts to show interactions of different compartments of the living and nonliving components of the ecosystem. A conceptual model describes ecosystem structure and dynamics and shows how environmental disturbances affect the ecosystem; however, its ability to predict the effects of these disturbances is limited. Analytical and simulation models, in contrast, are mathematical methods of describing ecosystems that are indeed capable of predicting the effects of potential environmental changes without direct experimentation, although with some limitations as to accuracy. An analytical model is an ecosystem model that is created using simple mathematical formulas to predict the effects of environmental disturbances on ecosystem structure and dynamics. A simulation model is an ecosystem model that is created using complex computer algorithms to holistically model ecosystems and to predict the effects of environmental disturbances on ecosystem structure and dynamics. Ideally, these models are accurate enough to determine which components of the ecosystem are particularly sensitive to disturbances, and they can serve as a guide to ecosystem managers (such as conservation ecologists or fisheries biologists) in the practical maintenance of ecosystem health.
Conceptual Models
Conceptual models are useful for describing ecosystem structure and dynamics and for demonstrating the relationships between different organisms in a community and their environment. Conceptual models are usually depicted graphically as flow charts. The organisms and their resources are grouped into specific compartments with arrows showing the relationship and transfer of energy or nutrients between them. Thus, these diagrams are sometimes called compartment models.
To model the cycling of mineral nutrients, organic and inorganic nutrients are subdivided into those that are bioavailable (ready to be incorporated into biological macromolecules) and those that are not. For example, in a terrestrial ecosystem near a deposit of coal, carbon will be available to the plants of this ecosystem as carbon dioxide gas in a short-term period, not from the carbon-rich coal itself. However, over a longer period, microorganisms capable of digesting coal will incorporate its carbon or release it as natural gas (methane, CH4), changing this unavailable organic source into an available one. This conversion is greatly accelerated by the combustion of fossil fuels by humans, which releases large amounts of carbon dioxide into the atmosphere. This is thought to be a major factor in the rise of the atmospheric carbon dioxide levels in the industrial age. The carbon dioxide released from burning fossil fuels is produced faster than photosynthetic organisms can use it. This process is intensified by the reduction of photosynthetic trees because of worldwide deforestation. Most scientists agree that high atmospheric carbon dioxide is a major cause of global climate change.
Conceptual models are also used to show the flow of energy through particular ecosystems. Figure \(\PageIndex{7}\) is based on Howard T. Odum’s classical study of the Silver Springs, Florida, holistic ecosystem in the mid-twentieth century.2 This study shows the energy content and transfer between various ecosystem compartments.
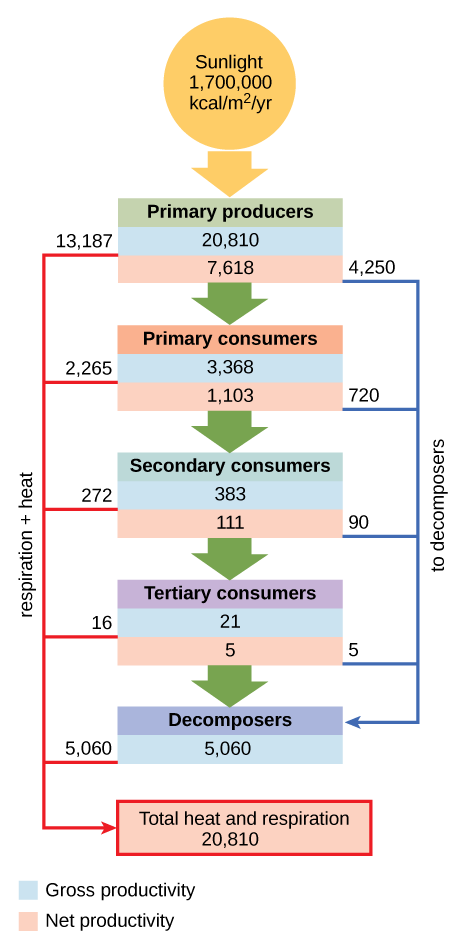
Exercise
Why do you think the value for gross productivity of the primary producers is the same as the value for total heat and respiration (20,810 kcal/m2/yr)?
- Answer
-
According to the first law of thermodynamics, energy can neither be created nor destroyed. Eventually, all energy consumed by living systems is lost as heat or used for respiration, and the total energy output of the system must equal the energy that went into it.
Analytical and Simulation Models
The major limitation of conceptual models is their inability to predict the consequences of changes in ecosystem species and/or environment. Ecosystems are dynamic entities and subject to a variety of abiotic and biotic disturbances caused by natural forces and/or human activity. Ecosystems altered from their initial equilibrium state can often recover from such disturbances and return to a state of equilibrium. As most ecosystems are subject to periodic disturbances and are often in a state of change, they are usually either moving toward or away from their equilibrium state. There are many of these equilibrium states among the various components of an ecosystem, which affects the ecosystem overall. Furthermore, as humans have the ability to greatly and rapidly alter the species content and habitat of an ecosystem, the need for predictive models that enable understanding of how ecosystems respond to these changes becomes more crucial.
Analytical models often use simple, linear components of ecosystems, such as food chains, and are known to be complex mathematically; therefore, they require a significant amount of mathematical knowledge and expertise. Although analytical models have great potential, their simplification of complex ecosystems is thought to limit their accuracy. Simulation models that use computer programs are better able to deal with the complexities of ecosystem structure.
A recent development in simulation modeling uses supercomputers to create and run individual-based simulations, which accounts for the behavior of individual organisms and their effects on the ecosystem as a whole. These simulations are considered to be the most accurate and predictive of the complex responses of ecosystems to disturbances.

Résumé
Les écosystèmes existent sur terre, en mer, dans l'air et sous terre. Différentes méthodes de modélisation des écosystèmes sont nécessaires pour comprendre comment les perturbations environnementales affecteront la structure et la dynamique des écosystèmes. Les modèles conceptuels sont utiles pour montrer les relations générales entre les organismes et le flux de matières ou d'énergie entre eux. Des modèles analytiques sont utilisés pour décrire les chaînes alimentaires linéaires, et les modèles de simulation fonctionnent mieux avec des réseaux alimentaires holistiques.
Notes
- 1 Nature (vol. 458, 1er avril 2009)
- 2 Howard T. Odum, « Structure trophique et productivité de Silver Springs, Floride », Monographies écologiques 27, n° 1 (1957) : 47—112.
Lexique
- modèle analytique
- modèle d'écosystème créé à l'aide de formules mathématiques pour prédire les effets des perturbations environnementales sur la structure et la dynamique des écosystèmes
- consommateur principal
- organisme au sommet de la chaîne alimentaire
- modèle conceptuel
- (également, modèles de compartiments) modèle d'écosystème qui consiste en des organigrammes qui montrent les interactions entre les différents compartiments des composantes vivantes et non vivantes de l'écosystème
- réseau alimentaire détritique
- type de réseau alimentaire dans lequel les principaux consommateurs sont des décomposeurs ; ceux-ci sont souvent associés à des réseaux alimentaires de pâturage au sein d'un même écosystème
- écosystème
- communauté d'organismes vivants et leurs interactions avec leur environnement abiotique
- dynamique des écosystèmes
- étude des modifications de la structure de l'écosystème causées par des modifications de l'environnement ou des forces internes
- équilibre
- état stable d'un écosystème où tous les organismes sont en équilibre avec leur environnement et les uns avec les autres
- chaîne alimentaire
- représentation linéaire d'une chaîne de producteurs primaires, de consommateurs primaires et de consommateurs de haut niveau utilisée pour décrire la structure et la dynamique de l'écosystème
- réseau alimentaire
- représentation graphique d'un réseau holistique et non linéaire de producteurs primaires, de consommateurs primaires et de consommateurs de haut niveau utilisée pour décrire la structure et la dynamique de l'écosystème
- réseau alimentaire de pâturage
- type de réseau trophique dans lequel les principaux producteurs sont soit des plantes terrestres, soit du phytoplancton dans l'eau ; souvent associé à un réseau trophique détritique au sein du même écosystème
- modèle d'écosystème holistique
- étude qui tente de quantifier la composition, les interactions et la dynamique d'écosystèmes entiers ; souvent limitée par des difficultés économiques et logistiques, selon l'écosystème
- mésocosme
- portion d'un écosystème naturel à utiliser pour des expériences
- microcosme
- recréation d'écosystèmes naturels entièrement dans un environnement de laboratoire à des fins expérimentales
- consommateur principal
- niveau trophique qui tire son énergie des principaux producteurs d'un écosystème
- producteur principal
- niveau trophique qui tire son énergie de la lumière solaire, de produits chimiques inorganiques ou de matières organiques mortes et/ou en décomposition
- résilience (écologique)
- vitesse à laquelle un écosystème retrouve son équilibre après avoir été perturbé
- résistance (écologique)
- capacité d'un écosystème à maintenir son équilibre malgré les perturbations
- consommateur secondaire
- généralement un carnivore qui mange des consommateurs primaires
- modèle de simulation
- modèle d'écosystème créé à l'aide de programmes informatiques pour modéliser les écosystèmes de manière holistique et pour prédire les effets des perturbations environnementales sur la structure et la dynamique des écosystèmes
- consommateur tertiaire
- carnivore qui mange d'autres carnivores
- niveau trophique
- position d'une espèce ou d'un groupe d'espèces dans une chaîne alimentaire ou un réseau alimentaire