
30.6 : Systèmes sensoriels et réponses des plantes
- Page ID
- 189418

Compétences à développer
- Décrire comment la lumière rouge et bleue affecte la croissance des plantes et les activités métaboliques
- Discutez du gravitropisme
- Comprendre comment les hormones affectent la croissance et le développement des plantes
- Décrire le thigmotropisme, le thigmonastisme et la thigmogenèse
- Expliquer comment les plantes se défendent contre les prédateurs et réagissent aux blessures
Les animaux peuvent réagir aux facteurs environnementaux en s'installant dans un nouvel endroit. Les plantes sont toutefois enracinées sur place et doivent répondre aux facteurs environnementaux environnants. Les plantes disposent de systèmes sophistiqués pour détecter et réagir à la lumière, à la gravité, à la température et au toucher physique. Les récepteurs détectent les facteurs environnementaux et transmettent l'information aux systèmes effecteurs, souvent par le biais de messagers chimiques intermédiaires, afin de provoquer la réponse des plantes.
Réponses des plantes à la lumière
Les plantes ont un certain nombre d'utilisations sophistiquées de la lumière qui vont bien au-delà de leur capacité à photosynthétiser des sucres de faible poids moléculaire en utilisant uniquement du dioxyde de carbone, de la lumière et de l'eau. La photomorphogenèse est la croissance et le développement des plantes en réponse à la lumière. Il permet aux plantes d'optimiser leur utilisation de la lumière et de l'espace. La photopériodisme est la capacité d'utiliser la lumière pour suivre le temps. Les plantes peuvent connaître l'heure du jour et l'époque de l'année en détectant et en utilisant différentes longueurs d'onde de la lumière solaire. Le phototropisme est une réponse directionnelle qui permet aux plantes de pousser vers la lumière ou même à s'en éloigner.
La détection de la lumière dans l'environnement est importante pour les plantes ; elle peut être cruciale pour la compétition et la survie. La réponse des plantes à la lumière est médiée par différents photorécepteurs, qui sont composés d'une protéine liée de manière covalente à un pigment absorbant la lumière appelé chromophore. Ensemble, ces deux éléments sont appelés chromoprotéines.
Les régions rouge/rouge foncé et bleu-violet du spectre de la lumière visible déclenchent le développement structurel des plantes. Les photorécepteurs sensoriels absorbent la lumière dans ces régions particulières du spectre de la lumière visible en raison de la qualité de la lumière disponible dans le spectre de la lumière du jour. Dans les habitats terrestres, l'absorption de lumière par les chlorophylles atteint son maximum dans les régions bleues et rouges du spectre. À mesure que la lumière traverse la canopée et que les longueurs d'onde bleues et rouges sont absorbées, le spectre se déplace vers l'extrémité rouge extrême, déplaçant la communauté végétale vers les plantes les mieux adaptées pour répondre à la lumière rouge lointain. Les récepteurs de lumière bleue permettent aux plantes de mesurer la direction et l'abondance de la lumière solaire, qui est riche en émissions bleu-vert. L'eau absorbe la lumière rouge, ce qui rend la détection de la lumière bleue essentielle pour les algues et les plantes aquatiques.
Le système phytochrome et la réponse rouge/rouge profond
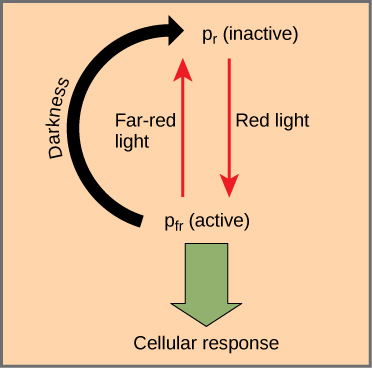
Les phytochromes sont une famille de chromoprotéines dotées d'un chromophore tétrapyrrole linéaire, similaire au groupe de tête photoabsorbeur de lumière du tétrapyrrole annelé de la chlorophylle. Les phytochromes ont deux formes photoconvertibles : Pr et Pfr. Le Pr absorbe la lumière rouge (~667 nm) et est immédiatement converti en Pfr. Le Pfr absorbe la lumière rouge lointaine (~730 nm) et est rapidement reconverti en Pr. L'absorption de lumière rouge ou rouge lointain entraîne une modification massive de la forme du chromophore, modifiant ainsi la conformation et l'activité de la protéine phytochrome à laquelle il est lié. Le Pfr est la forme physiologiquement active de la protéine ; par conséquent, l'exposition à la lumière rouge produit une activité physiologique. L'exposition à une lumière rouge lointaine inhibe l'activité du phytochrome. Ensemble, les deux formes représentent le système phytochrome (Figure\(\PageIndex{1}\)).
Le système phytochrome agit comme un interrupteur biologique. Il surveille le niveau, l'intensité, la durée et la couleur de la lumière ambiante. L'effet de la lumière rouge est réversible en projetant immédiatement une lumière rouge lointaine sur l'échantillon, qui convertit la chromoprotéine en forme Pr inactive. De plus, le Pfr peut lentement revenir à Pr dans l'obscurité ou se décomposer avec le temps. Dans tous les cas, la réponse physiologique induite par la lumière rouge est inversée. La forme active du phytochrome (Pfr) peut activer directement d'autres molécules du cytoplasme, ou elle peut être acheminée vers le noyau, où elle active ou réprime directement l'expression de gènes spécifiques.
Une fois que le système phytochrome a évolué, les plantes l'ont adapté pour répondre à divers besoins. Non filtré, le plein soleil contient beaucoup plus de lumière rouge que de lumière rouge lointaine. Comme la chlorophylle est fortement absorbée dans la région rouge du spectre visible, mais pas dans la région rouge extrême, toute plante se trouvant à l'ombre d'une autre plante du sol forestier sera exposée à une lumière appauvrie en rouge et enrichie en rouge profond. La prépondérance de la lumière rouge foncé transforme le phytochrome présent dans les feuilles ombragées en forme Pr (inactive), ralentissant ainsi la croissance. Les zones non ombragées (ou même moins ombragées) les plus proches du sol forestier émettent plus de lumière rouge ; les feuilles exposées à ces zones détectent la lumière rouge, qui active la forme Pfr et induit la croissance. En bref, les pousses des plantes utilisent le système phytochrome pour pousser loin de l'ombre et se diriger vers la lumière. La concurrence pour la lumière étant si féroce dans une communauté végétale dense, les avantages évolutifs du système phytochrome sont évidents.
Dans les graines, le système phytochrome n'est pas utilisé pour déterminer la direction et la qualité de la lumière (ombragée ou non ombragée). Au lieu de cela, est-il simplement utilisé pour déterminer s'il y a de la lumière ? Ceci est particulièrement important pour les espèces à très petites graines, comme la laitue. En raison de leur taille, les graines de laitue ont peu de réserves alimentaires. Leurs plants ne peuvent pas pousser longtemps avant de manquer de combustible. S'ils germaient même à un centimètre sous la surface du sol, les plantules n'atteindraient jamais la lumière du soleil et mourraient. Dans l'obscurité, le phytochrome se trouve sous forme Pr (forme inactive) et la graine ne germe pas ; elle ne germe que si elle est exposée à la lumière à la surface du sol. Lors de l'exposition à la lumière, le Pr est converti en Pfr et la germination se poursuit.
Les plantes utilisent également le système phytochrome pour détecter le changement de saison. La photopériodie est une réponse biologique au moment et à la durée du jour et de la nuit. Il contrôle la floraison, la formation des bourgeons d'hiver et la croissance végétative. La détection des changements saisonniers est essentielle à la survie des plantes. Bien que la température et l'intensité lumineuse influencent la croissance des plantes, elles ne constituent pas des indicateurs fiables de la saison car elles peuvent varier d'une année à l'autre. La durée du jour est un meilleur indicateur de la période de l'année.
Comme indiqué ci-dessus, la lumière solaire non filtrée est riche en lumière rouge mais déficiente en lumière rouge lointaine. Ainsi, à l'aube, toutes les molécules de phytochrome présentes dans une feuille se transforment rapidement en la forme Pfr active et restent sous cette forme jusqu'au coucher du soleil. Dans l'obscurité, la forme Pfr met des heures à revenir lentement à la forme Pr. Si la nuit est longue (comme en hiver), toute la forme Pfr revient. Si la nuit est courte (comme en été), une quantité considérable de Pfr peut subsister au lever du soleil. En détectant le rapport Pr/Pfr à l'aube, une plante peut déterminer la durée du cycle jour/nuit. De plus, les feuilles conservent ces informations pendant plusieurs jours, ce qui permet de comparer la durée de la nuit précédente à celle des nuits précédentes. Les nuits plus courtes indiquent le printemps pour la plante ; lorsque les nuits s'allongent, l'automne approche. Ces informations, associées à la détection de la température et de la disponibilité de l'eau, permettent aux plantes de déterminer la période de l'année et d'ajuster leur physiologie en conséquence. Les plantes à jours courts (longues nuits) utilisent ces informations pour fleurir à la fin de l'été et au début de l'automne, lorsque les nuits dépassent une durée critique (souvent huit heures ou moins). Les plantes qui durent de longues journées (courtes nuits) fleurissent au printemps, lorsque l'obscurité est inférieure à une durée critique (souvent de 8 à 15 heures). Toutes les plantes n'utilisent pas le système phytochrome de cette manière. La floraison des plantes neutres en jours n'est pas régulée par la durée du jour.
Lien de carrière : horticulteur
Le mot « horticulteur » vient des mots latins désignant jardin (hortus) et culture (cultura). Cette carrière a été révolutionnée par les progrès réalisés dans la compréhension des réponses des plantes aux stimuli environnementaux. Les producteurs de cultures, de fruits, de légumes et de fleurs étaient auparavant limités par le fait qu'ils devaient semer et récolter en fonction de la saison. Les horticulteurs peuvent désormais manipuler les plantes pour augmenter la production de feuilles, de fleurs ou de fruits en comprenant comment les facteurs environnementaux influent sur la croissance et le développement des plantes.
La gestion des serres est un élément essentiel de la formation des horticulteurs. Pour allonger la nuit, les plantes sont recouvertes d'un tissu occultant. Les plantes à jours longs sont irradiées à la lumière rouge en hiver pour favoriser une floraison précoce. Par exemple, la lumière fluorescente (blanc froid) à haute longueur d'onde bleue favorise la croissance des feuilles et est excellente pour démarrer les semis. Les lampes à incandescence (ampoules standard) sont riches en lumière rouge et favorisent la floraison de certaines plantes. Le moment de la maturation des fruits peut être augmenté ou retardé en appliquant des hormones végétales. Récemment, des progrès considérables ont été réalisés dans le développement de variétés de plantes adaptées à différents climats et résistantes aux ravageurs et aux dommages dus au transport. Le rendement et la qualité des cultures se sont améliorés grâce aux applications pratiques des connaissances sur les réponses des plantes aux stimuli et aux hormones externes.
Les horticulteurs trouvent un emploi dans des laboratoires privés et gouvernementaux, des serres, des jardins botaniques et dans les domaines de la production ou de la recherche. Ils améliorent les cultures en appliquant leurs connaissances en génétique et en physiologie végétale. Pour se préparer à une carrière en horticulture, les étudiants suivent des cours de botanique, de physiologie végétale, de pathologie végétale, d'aménagement paysager et de sélection végétale. Pour compléter ces cours traditionnels, les majors en horticulture ajoutent des études en économie, en commerce, en informatique et en communication.
Les réponses de la lumière bleue
Le phototropisme, c'est-à-dire la flexion directionnelle d'une plante vers ou loin d'une source de lumière, est une réponse aux longueurs d'onde bleues de la lumière. Le phototropisme positif est une croissance vers une source de lumière (Figure\(\PageIndex{2}\)), tandis que le phototropisme négatif (également appelé skototropisme) est une croissance loin de la lumière.
Les phototropines, bien nommées, sont des récepteurs à base de protéines responsables de la médiation de la réponse phototropique. Comme tous les photorécepteurs végétaux, les phototropines se composent d'une partie protéique et d'une partie absorbant la lumière, appelée chromophore. Dans les phototropines, le chromophore est une molécule de flavine liée par covalence ; les phototropines appartiennent donc à une classe de protéines appelées flavoprotéines.
Les autres réactions contrôlées par les phototropines sont l'ouverture et la fermeture des feuilles, le mouvement des chloroplastes et l'ouverture des stomates. Cependant, de toutes les réponses contrôlées par les phototropines, c'est le phototropisme qui a été étudié le plus longtemps et qui est le mieux compris.
Dans leur traité de 1880 The Power of Movements in Plants, Charles Darwin et son fils Francis ont d'abord décrit le phototropisme comme la flexion des plantules vers la lumière. Darwin a observé que la lumière était perçue par l'extrémité de la plante (le méristème apical), mais que la réponse (flexion) se produisait dans une autre partie de la plante. Ils ont conclu que le signal devait se déplacer du méristème apical jusqu'à la base de la plante.

En 1913, Peter Boysen-Jensen a démontré qu'un signal chimique produit à l'extrémité de la plante était responsable de la flexion de la base. Il a coupé l'extrémité d'un plant, recouvert la section coupée d'une couche de gélatine, puis a remplacé la pointe. La plantule se penchait vers la lumière lorsqu'elle était illuminée. Cependant, lorsque des flocons de mica imperméables ont été insérés entre la pointe et la base coupée, le plant ne s'est pas plié. Un raffinement de l'expérience a montré que le signal se déplaçait sur la face ombragée de la plantule. Lorsque la plaque de mica a été insérée du côté éclairé, la plante s'est pliée vers la lumière. Le signal chimique stimulait donc la croissance, car la réponse phototropique impliquait une élongation cellulaire plus rapide sur la face ombrée que sur la face éclairée. Nous savons maintenant que lorsque la lumière traverse la tige d'une plante, elle est diffractée et génère une activation de la phototropine à travers la tige. La majeure partie de l'activation se produit du côté éclairé, provoquant l'accumulation de l'hormone végétale acide indole acétique (IAA) sur le côté ombragé. Les cellules souches s'allongent sous l'influence de l'IAA.
Les cryptochromes sont une autre classe de photorécepteurs absorbant la lumière bleue qui contiennent également un chromophore à base de flavine. Les cryptochromes définissent le cycle d'activité de 24 heures de la plante, également connu sous le nom de rythme circadien, à l'aide de signaux de lumière bleue. Certaines preuves indiquent que les cryptochromes agissent de concert avec les phototropines pour médier la réponse phototropique.
Lien vers l'apprentissage
Utilisez le menu de navigation situé dans le panneau de gauche de ce site Web pour voir des images de plantes en mouvement.
Réponses des plantes à la gravité
Qu'elles germent ou non à la lumière ou dans l'obscurité totale, les pousses poussent généralement du sol et les racines poussent vers le bas dans le sol. Une plante posée sur le côté dans l'obscurité enverra les pousses vers le haut lorsqu'on lui en donnera suffisamment de temps. Le gravitropisme garantit que les racines poussent dans le sol et que les pousses poussent vers la lumière du soleil. La croissance de l'extrémité apicale des pousses vers le haut est appelée gravitropisme négatif, tandis que la croissance des racines vers le bas est appelée gravitropisme positif.
Les amyloplastes (également appelés statolithes) sont des plastes spécialisés qui contiennent des granules d'amidon et se déposent vers le bas sous l'effet de la gravité. Les amyloplastes se trouvent dans les pousses et dans les cellules spécialisées de la calotte racinaire. Lorsqu'une plante est inclinée, les statolithes tombent sur la nouvelle paroi cellulaire inférieure. Quelques heures plus tard, la pousse ou la racine poussera dans la nouvelle direction verticale.
Le mécanisme qui intervient dans le gravitropisme est relativement bien compris. Lorsque les amyloplastes se déposent au bas des cellules sensibles à la gravité de la racine ou de la pousse, ils entrent en contact physique avec le réticulum endoplasmique (RE), provoquant la libération d'ions calcium depuis l'intérieur du RE. Cette signalisation calcique dans les cellules provoque le transport polaire de l'hormone végétale IAA vers le bas de la cellule. Dans les racines, une forte concentration d'IAA inhibe l'élongation cellulaire. L'effet ralentit la croissance sur la face inférieure de la racine, tandis que les cellules se développent normalement sur la face supérieure. L'IAA a l'effet inverse sur les pousses, où une concentration plus élevée sur la face inférieure de la pousse stimule l'expansion cellulaire, provoquant ainsi la croissance de la pousse. Une fois que la pousse ou la racine commence à se développer verticalement, les amyloplastes reprennent leur position normale. D'autres hypothèses, impliquant la cellule entière dans l'effet gravitropisme, ont été proposées pour expliquer pourquoi certains mutants dépourvus d'amyloplastes peuvent encore présenter une faible réponse gravitropique.
Réponses à la croissance
La réponse sensorielle d'une plante à des stimuli externes repose sur des messagers chimiques (hormones). Les hormones végétales affectent tous les aspects de la vie végétale, de la floraison à la fructification et à la maturation, et du phototropisme à la chute des feuilles. Chaque cellule d'une plante peut potentiellement produire des hormones végétales. Elles peuvent agir dans leur cellule d'origine ou être transportées vers d'autres parties du corps végétal, avec de nombreuses réponses végétales impliquant l'interaction synergique ou antagoniste de deux hormones ou plus. En revanche, les hormones animales sont produites dans des glandes spécifiques et transportées vers un site éloigné pour agir, et elles agissent seules.
Les hormones végétales sont un groupe de substances chimiques non apparentées qui influent sur la morphogenèse des plantes. Cinq principales hormones végétales sont traditionnellement décrites : les auxines (en particulier l'IAA), les cytokinines, les gibbérellines, l'éthylène et l'acide abscissique. De plus, d'autres nutriments et conditions environnementales peuvent être considérés comme des facteurs de croissance.
Auxines
Le terme auxine est dérivé du mot grec auxéine, qui signifie « grandir ». Les auxines sont les principales hormones responsables de l'élongation cellulaire lors du phototropisme et du gravitropisme. Ils contrôlent également la différenciation du méristème en tissu vasculaire et favorisent le développement et la disposition des feuilles. Alors que de nombreuses auxines synthétiques sont utilisées comme herbicides, l'IAA est la seule auxine naturelle qui présente une activité physiologique. La dominance apicale, c'est-à-dire l'inhibition de la formation des bourgeons latéraux, est déclenchée par les auxines produites dans le méristème apical. La floraison, la nouaison et la maturation des fruits ainsi que l'inhibition de l'abscission (chute des feuilles) sont d'autres réactions des plantes soumises au contrôle direct ou indirect des auxines. Les auxines agissent également comme relais des effets de la lumière bleue et des réponses rouge/rouge lointain.
L'utilisation commerciale des auxines est très répandue dans les pépinières et pour la production végétale. L'IAA est utilisé comme hormone d'enracinement pour favoriser la croissance des racines adventives sur les boutures et les feuilles détachées. L'application d'auxines synthétiques sur les plants de tomates en serre favorise le développement normal des fruits. L'application extérieure d'auxine favorise la synchronisation de la nouaison et de la chute des fruits afin de coordonner la saison de récolte. Les fruits tels que les concombres sans pépins peuvent être incités à produire des fruits en traitant les fleurs des plantes non fertilisées avec des auxines.
Cytokinines
L'effet des cytokinines a été signalé pour la première fois lorsqu'il a été découvert que l'ajout de l'endosperme liquide des noix de coco à des embryons de plantes en développement en culture stimulait leur croissance. Le facteur de croissance stimulant s'est avéré être la cytokinine, une hormone qui favorise la cytokinèse (division cellulaire). Près de 200 cytokinines naturelles ou synthétiques sont connues à ce jour. Les cytokinines sont plus abondantes dans les tissus en croissance, tels que les racines, les embryons et les fruits, où se produit la division cellulaire. Les cytokinines sont connues pour retarder la sénescence dans les tissus foliaires, favoriser la mitose et stimuler la différenciation du méristème dans les pousses et les racines. De nombreux effets sur le développement des plantes sont dus à l'influence des cytokinines, soit en association avec l'auxine, soit avec une autre hormone. Par exemple, la dominance apicale semble résulter d'un équilibre entre les auxines qui inhibent les bourgeons latéraux et les cytokinines qui favorisent une croissance plus touffue.
Gibbérellins
Les gibbérellines (GA) sont un groupe d'environ 125 hormones végétales étroitement apparentées qui stimulent l'élongation des pousses, la germination des graines et la maturation des fruits et des fleurs. Les AG sont synthétisés dans les méristèmes apicaux des racines et des tiges, les jeunes feuilles et les embryons de graines. Dans les zones urbaines, des antagonistes de l'AG sont parfois appliqués aux arbres situés sous des lignes à haute tension afin de contrôler la croissance et de réduire la fréquence de l'élagage.
Les AG mettent fin à la dormance (état d'inhibition de la croissance et du développement) des graines des plantes qui ont besoin d'une exposition au froid ou à la lumière pour germer. L'acide abscissique est un puissant antagoniste de l'action de l'AG. Les autres effets des AG incluent l'expression sexuelle, le développement des fruits sans pépins et le retard de la sénescence des feuilles et des fruits. Les raisins sans pépins sont obtenus selon des méthodes de sélection standard et contiennent des graines discrètes qui ne se développent pas. Comme les AG sont produits par les graines et que le développement des fruits et l'allongement de la tige sont contrôlés par l'AG, ces variétés de raisins produisent normalement de petits fruits en grappes compactes. Les raisins en cours de maturation sont régulièrement traités à l'AG pour favoriser des fruits de plus grande taille, ainsi que des grappes plus lâches (tiges plus longues), ce qui réduit les cas d'infection par le mildiou (Figure\(\PageIndex{3}\)).

Acide abscissique
L'acide abscissique (ABA), une hormone végétale, a été découvert pour la première fois en tant qu'agent responsable de l'abscission ou de la chute des capsules de coton. Cependant, des études plus récentes indiquent que l'ABA ne joue qu'un rôle mineur dans le processus d'abscission. L'ABA s'accumule en réponse à des conditions environnementales stressantes, telles que la déshydratation, les températures froides ou la réduction de la durée de la journée. Son activité neutralise de nombreux effets de croissance des AG et des auxines. L'ABA inhibe l'allongement de la tige et induit la dormance des bourgeons latéraux.
L'ABA induit la dormance des graines en bloquant la germination et en favorisant la synthèse des protéines de réserve. Les plantes adaptées aux climats tempérés ont besoin d'une longue période de froid avant que les graines ne germent. Ce mécanisme empêche les jeunes plants de germer trop tôt par temps exceptionnellement chaud en hiver. Au fur et à mesure que l'hormone se décompose au cours de l'hiver, les graines sortent de la dormance et germent lorsque les conditions sont favorables au printemps. Un autre effet de l'ABA est de favoriser le développement des bourgeons d'hiver ; il intervient dans la conversion du méristème apical en bourgeon dormant. La faible humidité du sol entraîne une augmentation de l'ABA, ce qui provoque la fermeture des stomates, réduisant ainsi la perte d'eau dans les bourgeons d'hiver.
éthylène
L'éthylène est associé à la maturation des fruits, au flétrissement des fleurs et à la chute des feuilles. L'éthylène est inhabituel car il s'agit d'un gaz volatil (C 2 H 4). Il y a des centaines d'années, lorsque des lampadaires à gaz ont été installés dans les rues des villes, les arbres qui poussaient près des lampadaires ont développé des troncs tordus et épaissis et ont perdu leurs feuilles plus tôt que prévu. Ces effets ont été causés par la volatilisation de l'éthylène par les lampes.
Les tissus vieillissants (en particulier les feuilles sénescentes) et les nœuds des tiges produisent de l'éthylène. L'effet le plus connu de l'hormone est toutefois de favoriser la maturation des fruits. L'éthylène stimule la conversion de l'amidon et des acides en sucres. Certaines personnes stockent les fruits immatures, tels que les avocats, dans un sac en papier scellé pour accélérer la maturation ; le gaz libéré par le premier fruit à mûrir accélérera la maturation du fruit restant. L'éthylène déclenche également l'abscission des feuilles et des fruits, la décoloration et la chute des fleurs, et favorise la germination de certaines céréales et la germination des bulbes et des pommes de terre.
L'éthylène est largement utilisé en agriculture. Les producteurs de fruits commerciaux contrôlent le moment de la maturation des fruits grâce à l'application du gaz. Les horticulteurs empêchent la chute des feuilles chez les plantes ornementales en retirant l'éthylène des serres à l'aide de ventilateurs et d'une ventilation.
Hormones non traditionnelles
Des recherches récentes ont permis de découvrir un certain nombre de composés qui influencent également le développement des plantes. Leurs rôles sont moins connus que les effets des principales hormones décrites jusqu'à présent.
Les jasmonates jouent un rôle majeur dans les réponses défensives à l'herbivorie. Leurs niveaux augmentent lorsqu'une plante est blessée par un prédateur, ce qui entraîne une augmentation des métabolites secondaires toxiques. Ils contribuent à la production de composés volatils qui attirent les ennemis naturels des prédateurs. Par exemple, la mastication des plants de tomates par les chenilles entraîne une augmentation des niveaux d'acide jasmonique, ce qui déclenche la libération de composés volatils qui attirent les prédateurs du ravageur.
Les oligosaccharines jouent également un rôle dans la défense des plantes contre les infections bactériennes et fongiques. Ils agissent localement sur le site de la lésion et peuvent également être transportés vers d'autres tissus. Les strigolactones favorisent la germination des graines chez certaines espèces et inhibent le développement apical latéral en l'absence d'auxines. Les strigolactones jouent également un rôle dans l'établissement des mycorhizes, une association mutualiste de racines végétales et de champignons. Les brassinostéroïdes jouent un rôle important dans de nombreux processus développementaux et physiologiques. Les signaux entre ces composés et d'autres hormones, notamment l'auxine et les GA, amplifient leur effet physiologique. La dominance apicale, la germination des graines, le gravitropisme et la résistance au gel sont tous influencés positivement par les hormones. La croissance des racines et la chute des fruits sont inhibées par les stéroïdes.
Réactions des plantes au vent et au toucher
La pousse d'un plant de pois s'enroule autour d'un treillis, tandis qu'un arbre pousse en biais en réponse à de forts vents dominants. Ce sont des exemples de la façon dont les plantes réagissent au toucher ou au vent.
Le mouvement d'une plante soumise à une pression directionnelle constante est appelé thigmotropisme, du grec thigma qui signifie « toucher » et du tropisme qui signifie « direction ». Les vrilles en sont un exemple. La région méristématique des vrilles est très sensible au toucher ; un toucher léger provoquera une réaction d'enroulement rapide. Les cellules en contact avec une surface de support se contractent, tandis que les cellules du côté opposé du support se dilatent. L'application d'acide jasmonique est suffisante pour provoquer l'enroulement des vrilles sans stimulus mécanique.
Une réponse thigmonastique est une réponse tactile indépendante de la direction du stimulus. Dans le piège à mouches Venus, deux feuilles modifiées sont jointes par une charnière et bordées de fines dents en forme de fourche le long des bords extérieurs. De minuscules poils se trouvent à l'intérieur du piège. Lorsqu'un insecte se brosse contre ces poils déclencheurs, en touchant deux ou plusieurs d'entre eux à la suite, les feuilles se referment rapidement, piégeant ainsi la proie. Les glandes situées à la surface des feuilles sécrètent des enzymes qui digèrent lentement l'insecte. Les nutriments libérés sont absorbés par les feuilles, qui rouvrent pour le repas suivant.
La thigmomorphogenèse est un lent changement évolutif de la forme d'une plante soumise à des contraintes mécaniques continues. Lorsque les arbres se plient sous l'effet du vent, par exemple, la croissance est généralement ralentie et le tronc s'épaissit. Les tissus de renforcement, en particulier le xylème, sont produits pour ajouter de la rigidité afin de résister à la force du vent. Les chercheurs émettent l'hypothèse que les contraintes mécaniques induisent la croissance et la différenciation afin de renforcer les tissus. L'éthylène et le jasmonate sont probablement impliqués dans la thigmomorphogenèse.
Lien vers l'apprentissage

Utilisez le menu de gauche pour accéder à trois courtes vidéos : un piège à mouches Venus capturant des proies, la fermeture progressive de folioles sensibles et l'entrelacement de vrilles.
Réponses de défense contre les herbivores et les agents pathogènes
Les plantes font face à deux types d'ennemis : les herbivores et les agents pathogènes. Les herbivores, grands et petits, utilisent les plantes comme nourriture et les mâchent activement. Les agents pathogènes sont des agents pathogènes. Ces microorganismes infectieux, tels que les champignons, les bactéries et les nématodes, vivent de la plante et endommagent ses tissus. Les plantes ont développé diverses stratégies pour décourager ou tuer les attaquants.
La première ligne de défense des plantes est une barrière intacte et impénétrable. L'écorce et la cuticule cireuse peuvent protéger contre les prédateurs. Les autres adaptations contre l'herbivorie incluent les épines, qui sont des branches modifiées, et les épines, qui sont des feuilles modifiées. Ils découragent les animaux en leur causant des dommages physiques et en provoquant des éruptions cutanées et des réactions allergiques. La protection extérieure d'une plante peut être compromise par des dommages mécaniques, qui peuvent constituer un point d'entrée pour les agents pathogènes. Si la première ligne de défense est violée, la plante doit recourir à un ensemble différent de mécanismes de défense, tels que des toxines et des enzymes.
Les métabolites secondaires sont des composés qui ne sont pas directement dérivés de la photosynthèse et qui ne sont pas nécessaires à la respiration ou à la croissance et au développement des plantes. De nombreux métabolites sont toxiques et peuvent même être mortels pour les animaux qui les ingèrent. Certains métabolites sont des alcaloïdes qui découragent les prédateurs grâce à des odeurs nocives (comme les huiles volatiles de menthe et de sauge) ou à des goûts répulsifs (comme l'amertume de la quinine). D'autres alcaloïdes affectent les herbivores en provoquant soit une stimulation excessive (la caféine, par exemple), soit la léthargie associée aux opioïdes. Certains composés deviennent toxiques après ingestion ; par exemple, le cyanure de glycol contenu dans la racine de manioc libère du cyanure uniquement lorsqu'il est ingéré par l'herbivore.
Les blessures mécaniques et les attaques de prédateurs activent les mécanismes de défense et de protection à la fois dans les tissus endommagés et sur des sites plus éloignés du lieu de la lésion. Certaines réactions de défense se produisent en quelques minutes, d'autres en plusieurs heures. Les cellules infectées et environnantes peuvent mourir, arrêtant ainsi la propagation de l'infection.
La signalisation à longue distance déclenche une réponse systémique visant à dissuader le prédateur. Lorsque les tissus sont endommagés, les jasmonates peuvent favoriser la synthèse de composés toxiques pour les prédateurs. Les jasmonates provoquent également la synthèse de composés volatils qui attirent les parasitoïdes, des insectes qui passent leur stade de développement dans ou sur un autre insecte et finissent par tuer leur hôte. La plante peut activer l'abscission des tissus lésés s'ils sont irrémédiablement endommagés.
Résumé
Les plantes réagissent à la lumière par des changements de morphologie et d'activité. L'irradiation par la lumière rouge convertit le phytochrome photorécepteur en sa forme d'absorption de la lumière rouge lointain, le PFR. Cette forme contrôle la germination et la floraison en fonction de la durée du jour et déclenche la photosynthèse chez les plantes dormantes ou celles qui viennent de sortir du sol. Les récepteurs de lumière bleue, les cryptochromes et les phototropines sont responsables du phototropisme. Les amyloplastes, qui contiennent de lourds granules d'amidon, détectent la gravité. Les pousses présentent un gravitropisme négatif, tandis que les racines présentent un gravitropisme positif. Les hormones végétales, des composés naturels synthétisés en petites quantités, peuvent agir à la fois dans les cellules qui les produisent et dans les tissus et organes distants. Les auxines sont responsables de la dominance apicale, de la croissance des racines, de la croissance dirigée vers la lumière et de nombreuses autres réponses de croissance. Les cytokinines stimulent la division cellulaire et contrecarrent la dominance apicale dans les pousses. Les gibbérellines inhibent la dormance des graines et favorisent la croissance des tiges. L'acide abscissique induit la dormance des graines et des bourgeons et protège les plantes d'une perte excessive d'eau en favorisant la fermeture des stomates. L'éthylène accélère la maturation des fruits et la chute des feuilles. Les plantes réagissent au toucher par des mouvements rapides (thigmotropie et thigmonathie) et par une croissance différentielle lente (thigmomorphogenèse). Les plantes ont développé des mécanismes de défense contre les prédateurs et les agents pathogènes. Les barrières physiques telles que l'écorce et les épines protègent les tissus sensibles. Les plantes possèdent également des défenses chimiques, notamment des métabolites secondaires toxiques et des hormones, qui déclenchent des mécanismes de défense supplémentaires.
Lexique
- acide abscisique (ABA)
- hormone végétale qui induit la dormance des graines et d'autres organes
- abscission
- processus physiologique qui entraîne la chute d'un organe végétal (comme la chute d'une feuille ou d'un pétale)
- auxine
- hormone végétale qui influence l'élongation cellulaire (dans le phototropisme), le gravitropisme, la dominance apicale et la croissance des racines
- chromophore
- molécule qui absorbe la lumière
- cryptochrome
- protéine qui absorbe la lumière dans les régions bleues et ultraviolettes du spectre lumineux
- cytokinine
- hormone végétale qui favorise la division cellulaire
- éthylène
- hormone végétale volatile associée à la maturation des fruits, au flétrissement des fleurs et à la chute des feuilles
- gibbérelline (GA)
- hormone végétale qui stimule l'élongation des pousses, la germination des graines ainsi que la maturation et la chute des fruits et des fleurs
- jasmonates
- petite famille de composés dérivés de l'acide gras acide linoléique
- gravitropisme négatif
- croissance loin de la gravité de la Terre
- oligosaccharine
- hormone importante dans les défenses des plantes contre les infections bactériennes et fongiques
- photomorphogenèse
- croissance et développement des plantes en réponse à la lumière
- photopériodisme
- apparition de processus végétaux, tels que la germination et la floraison, selon la période de l'année
- phototropine
- récepteur de lumière bleue qui favorise le phototropisme, l'ouverture et la fermeture des stomates et d'autres réponses qui favorisent la photosynthèse
- phototropisme
- flexion directionnelle d'une plante vers une source de lumière
- phytochrome
- protéine pigmentaire végétale qui existe sous deux formes réversibles (Pr et Pfr) et qui médie les changements morphologiques en réponse à la lumière rouge
- gravitropisme positif
- croissance vers le centre gravitationnel de la Terre
- statolithe
- Organite végétal (également amyloplaste) contenant de lourds granules d'amidon
- strigolactone
- hormone qui favorise la germination des graines chez certaines espèces et inhibe le développement apical latéral en l'absence d'auxines
- thigmomorphogenèse
- réponse développementale au toucher
- thigmonastique
- croissance directionnelle d'une plante indépendamment de la direction dans laquelle le contact est appliqué
- thigmotropisme
- croissance directionnelle d'une plante en réponse à un contact constant