
28.3 : Superphylum Lophotrochozoa
- Page ID
- 189956

Compétences à développer
- Décrire les caractéristiques anatomiques et morphologiques uniques des vers plats, des rotifères, des Nemertea, des mollusques et des annélides
- Décrire le développement d'une cavité extracoléomique
- Discutez des avantages d'une véritable segmentation corporelle
- Expliquer les principales caractéristiques des platyhelminthes et leur importance en tant que parasites
- Décrire les caractéristiques des animaux classés dans le phylum Annelida
Les animaux appartenant au superphylum Lophotrochozoa sont des protostomes, dans lesquels le blastopore, ou point d'involution de l'ectoderme ou de la couche germinale externe, devient la bouche qui donne accès au tube digestif. C'est ce que l'on appelle la protostomie ou « première bouche ». Lors de la protostomie, des groupes solides de cellules se séparent de l'endoderme ou de la couche germinale interne pour former une couche mésodermique centrale de cellules. Cette couche se multiplie en une bande puis se divise en interne pour former le coelome ; ce coelome protostomique est donc appelé schizocoelom.
En tant que lophotrochozoaires, les organismes de ce superphylum possèdent soit un lophophore, soit des larves de trochophores. Les lophophores comprennent des groupes qui sont unis par la présence du lophophore, un ensemble de tentacules ciliés entourant la bouche. Les Lophophorata incluent les ascarides et plusieurs autres phylums. Ces clades sont maintenus lorsque les séquences d'ARN sont comparées. Les larves de trochophores sont caractérisées par deux bandes de cils autour du corps.
Les lophotrochozoaires sont triploblastiques et possèdent un mésoderme embryonnaire intercalé entre l'ectoderme et l'endoderme trouvés chez les cnidaires diploblastiques. Ces phylums sont également symétriques bilatéralement, ce qui signifie qu'une coupe longitudinale les divisera en côtés droit et gauche symétriques. Cela signifie également le début de la céphalisation, l'évolution d'une concentration de tissus nerveux et d'organes sensoriels dans la tête de l'organisme, où il rencontre pour la première fois son environnement.
Phylum Platyhelminthes
Les ascarides sont des organismes acélomés qui comprennent de nombreuses formes libres et parasitaires. La plupart des vers plats sont classés dans le superphylum Lophotrochozoa, qui comprend également les mollusques et les annélides. Les Platyhelminthes se composent de deux lignées : les Catenulides et les Rhabditophora. Les caténulides, ou « vers à chaîne », sont un petit clade d'un peu plus de 100 espèces. Ces vers se reproduisent généralement de manière asexuée par bourgeonnement. Cependant, la progéniture ne s'attache pas complètement aux parents et ressemble à une chaîne en apparence. Tous les vers plats restants dont il est question ici font partie de la Rhabditophora. De nombreux vers plats sont parasitaires, y compris d'importants parasites humains. Les ascaris possèdent trois couches de tissu embryonnaire qui donnent naissance à des surfaces qui recouvrent les tissus (provenant de l'ectoderme), les tissus internes (du mésoderme) et tapissent le système digestif (de l'endoderme). Le tissu épidermique est constitué de cellules monocouches ou d'une couche de cellules fusionnées (syncytium) qui recouvre une couche de muscle circulaire au-dessus d'une couche de muscle longitudinal. Les tissus mésodermiques comprennent des cellules mésenchymateuses qui contiennent du collagène et soutiennent les cellules sécrétoires qui sécrètent du mucus et d'autres substances à la surface. Les ascaris sont des acéléomates, de sorte que leur corps est solide entre la surface extérieure et la cavité du système digestif.
Processus physiologiques des vers plats
Les espèces libres de vers plats sont des prédateurs ou des charognards. Les formes parasitaires se nourrissent des tissus de leurs hôtes. La plupart des vers plats, tels que le planaire illustré à la figure\(\PageIndex{1}\), ont une cavité gastrovasculaire plutôt qu'un système digestif complet. Chez ces animaux, la « bouche » est également utilisée pour expulser les déchets du système digestif. Certaines espèces ont également une ouverture anale. L'intestin peut être un sac simple ou très ramifié. La digestion est extracellulaire, les matières digérées étant absorbées par phagocytose dans les cellules de la paroi intestinale. Un groupe, les cestodes, n'a pas de système digestif. Les ascaris ont un système excréteur constitué d'un réseau de tubules répartis dans tout le corps, avec des ouvertures donnant sur l'environnement et des cellules de flamme voisines, dont les cils battent pour diriger les liquides résiduaires concentrés dans les tubules hors du corps. Le système est responsable de la régulation des sels dissous et de l'excrétion des déchets azotés. Le système nerveux est constitué d'une paire de cordons nerveux qui parcourent le corps et qui sont reliés à un gros ganglion ou à une concentration de nerfs à l'extrémité antérieure du ver, où peuvent également se trouver une concentration de cellules photosensorielles et chimiosensorielles.
Il n'y a ni système circulatoire ni système respiratoire, les échanges de gaz et de nutriments dépendant de la diffusion et des jonctions cellules-cellules. Cela limite nécessairement l'épaisseur du corps de ces organismes, les obligeant à être des vers « plats ».
La plupart des espèces de vers plats sont monoïques et la fertilisation est généralement interne. La reproduction asexuée est courante dans certains groupes.
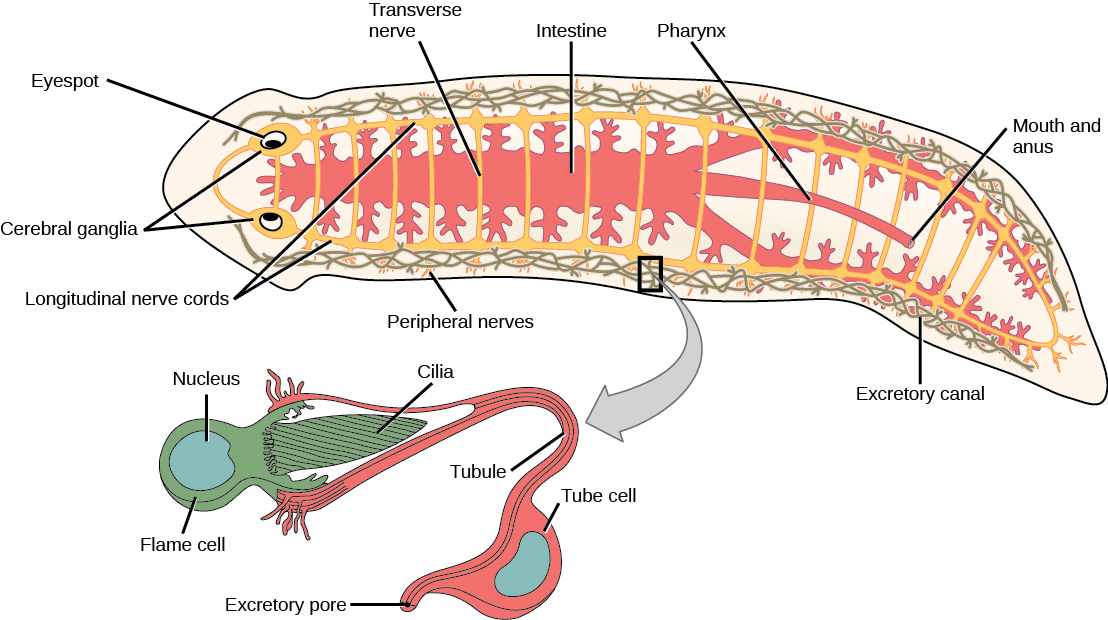
Diversité des vers plats
Les platyhelminthes sont traditionnellement divisés en quatre classes : les turbellaires, les monogènes, les trématodes et les cestodes (Figure\(\PageIndex{2}\)). Comme indiqué ci-dessus, les relations entre les membres de ces classes sont en cours de réévaluation, les turbellaires en particulier étant désormais considérés comme un groupe paraphylétique, un groupe qui n'a pas d'ancêtre commun.
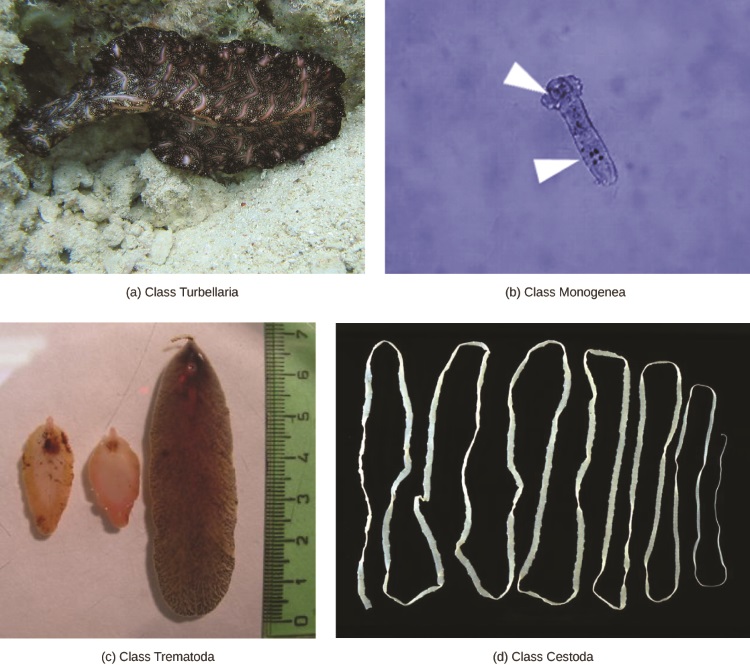
La classe Turbellaria comprend principalement des espèces marines libres, bien que certaines espèces vivent en eau douce ou dans des environnements terrestres humides. L'épiderme ventral des turbellaires est cilié et facilite leur locomotion. Certains turbellaires sont capables de réaliser de remarquables prouesses de régénération au cours desquelles leur corps peut se régénérer, même à partir d'un petit fragment.
Les monogènes sont des ectoparasites, principalement des poissons, dont le cycle vital est simple : une larve nageant librement se fixe au poisson pour commencer à se transformer en une forme adulte parasitaire. Le parasite n'a qu'un seul hôte et cet hôte n'est généralement qu'une seule espèce. Les vers peuvent produire des enzymes qui digèrent les tissus de l'hôte ou qui se contentent de brouter le mucus de surface et les particules de peau. La plupart des monogéniques sont hermaphrodites, mais les gamètes mâles se développent en premier, de sorte que la fécondation croisée est assez courante.
Les trématodes, ou douves, sont des parasites internes des mollusques et de nombreux autres groupes, y compris les humains. Les trématodes ont des cycles de vie complexes qui impliquent un hôte principal dans lequel se produit la reproduction sexuée et un ou plusieurs hôtes secondaires dans lesquels se produit une reproduction asexuée. L'hôte principal est presque toujours un mollusque. Les trématodes sont responsables de maladies humaines graves, notamment la schistosomiase, une douve sanguine. La maladie infecte environ 200 millions de personnes sous les tropiques, provoquant des lésions organiques et des symptômes chroniques tels que la fatigue. L'infection se produit lorsque l'être humain entre dans l'eau et qu'une larve, libérée par le principal escargot hôte, localise et pénètre dans la peau. Le parasite infecte divers organes du corps et se nourrit de globules rouges avant de se reproduire. De nombreux œufs sont libérés dans les matières fécales et se retrouvent dans un cours d'eau, où ils peuvent réinfecter l'escargot hôte principal.
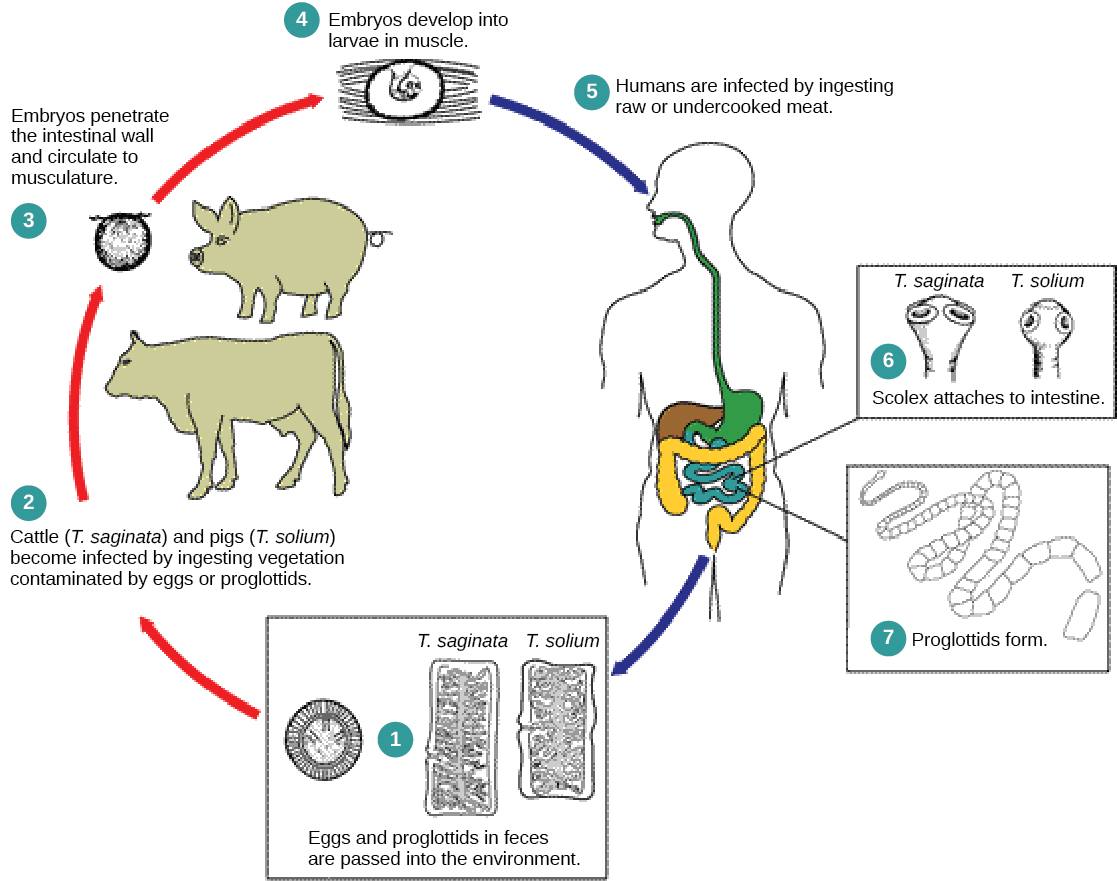
Les cestodes, ou ténias, sont également des parasites internes, principalement des vertébrés (Figure\(\PageIndex{3}\)). Les ténias vivent dans le tractus intestinal de l'hôte principal et restent fixés à l'aide d'une ventouse située à l'extrémité antérieure, ou scolex, du corps du ténia. Le reste du corps du ténia est constitué d'une longue série d'unités appelées proglottides, dont chacune peut contenir un système excréteur avec des cellules flammées, mais contenir des structures reproductrices, mâles et femelles. Les ténias ne possèdent pas de système digestif ; ils absorbent plutôt les nutriments contenus dans les matières alimentaires qui les transitent dans l'intestin de l'hôte.
Les proglottis sont produits au niveau du scolex et migrent graduellement jusqu'à l'extrémité du ténia ; à ce stade, ils sont « matures » et toutes les structures, à l'exception des œufs fécondés, ont dégénéré. La plupart des reproductions se produisent par fécondation croisée. Le proglottide se détache du corps du ver et est libéré dans les matières fécales de l'organisme. Les œufs sont consommés par un hôte intermédiaire. Le ver juvénile infecte l'hôte intermédiaire et s'installe, généralement dans les tissus musculaires. Lorsque le tissu musculaire est mangé par l'hôte principal, le cycle est terminé. Plusieurs ténias parasites des humains se transmettent en mangeant du porc, du bœuf et du poisson crus ou mal cuits.
Phylum Rotifera
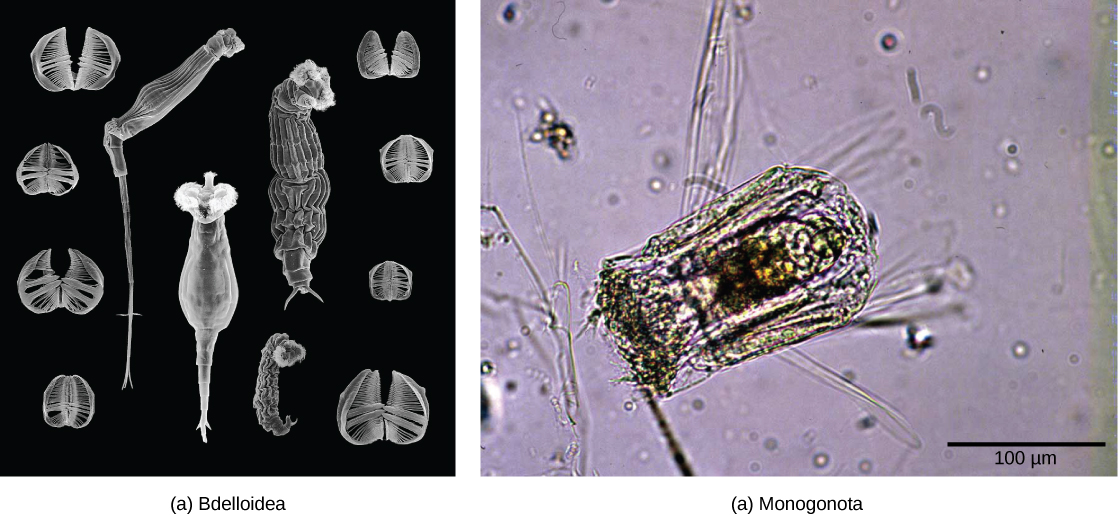
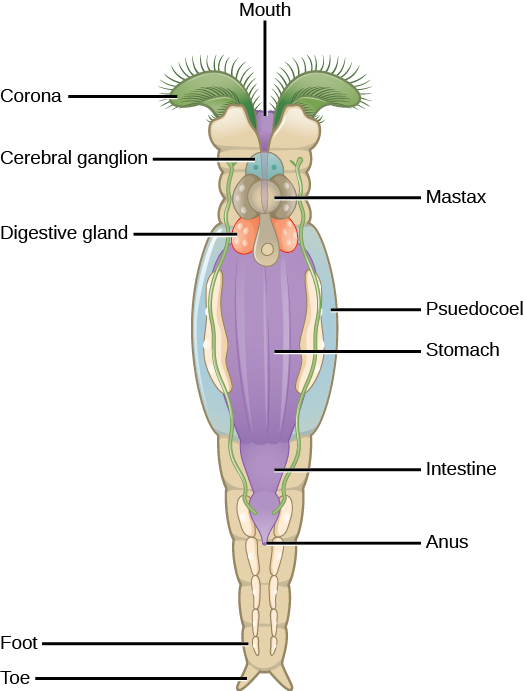
Les rotifères sont un groupe microscopique (environ 100 µm à 30 mm) d'organismes principalement aquatiques qui tirent leur nom de la couronne, une structure rotative en forme de roue recouverte de cils à leur extrémité antérieure (Figure\(\PageIndex{4}\)). Bien que leur taxonomie soit actuellement en évolution, un traitement classe les rotifères en trois classes : Bdelloidea, Monogononta et Seisonidea. La classification du groupe est toutefois en cours de révision, à mesure que de nouvelles preuves phylogénétiques seront disponibles. Il est possible que les « vers à tête épineuse » actuellement présents dans le phylum Acanthocephala soient incorporés à ce groupe à l'avenir.
La forme corporelle des rotifères se compose d'une tête (qui contient la couronne), d'un tronc (qui contient les organes) et du pied. Les rotifères sont généralement des organismes nageant librement et véritablement planctoniques, mais les orteils ou les extensions du pied peuvent sécréter une matière collante qui les aide à adhérer aux surfaces. La tête contient des organes sensoriels sous la forme d'un cerveau bilobé et de petits points oculaires situés près de la couronne.
Les rotifères sont des filtreurs qui mangent des matières mortes, des algues et d'autres organismes vivants microscopiques. Ils constituent donc des éléments très importants des réseaux alimentaires aquatiques. Les rotifères obtiennent de la nourriture dirigée vers la bouche par le courant créé par le mouvement de la couronne. Les particules alimentaires pénètrent dans la bouche et se dirigent vers la mastax (pharynx aux structures semblables à des mâchoires). Les aliments passent ensuite par les glandes digestives et salivaires, puis dans l'estomac, puis dans les intestins. Les déchets digestifs et excréteurs sont collectés dans une vessie cloacale avant d'être libérés par l'anus.
Les rotifères sont des pseudocoélomates que l'on trouve couramment dans l'eau douce et dans certains environnements d'eau salée à travers le monde. La figure\(\PageIndex{5}\) montre l'anatomie d'un rotifère appartenant à la classe des Bdelloidea. Environ 2 200 espèces de rotifères ont été identifiées. Les rotifères sont des organismes dioïques (dotés d'organes génitaux masculins ou féminins) et présentent un dimorphisme sexuel (les mâles et les femelles ont des formes différentes). De nombreuses espèces sont parthénogènes et présentent une haplodiploïdie, une méthode de détermination du sexe selon laquelle un ovule fécondé devient une femelle et un ovule non fécondé devient un mâle. Chez de nombreuses espèces dioïques, les mâles vivent peu de temps et sont plus petits, sans système digestif et ne possèdent qu'un seul testicule. Les femelles peuvent produire des œufs capables de dormance pour se protéger dans des conditions environnementales difficiles.
Phylum Nemertea
Les Nemertea sont communément appelés vers rubans. La plupart des espèces du phylum Nemertea sont marines, principalement benthiques ou vivant au fond, et environ 900 espèces sont connues. Cependant, des nemertini ont également été signalés dans des habitats d'eau douce et terrestres. La plupart des némertiens sont des carnivores qui se nourrissent de vers, de palourdes et de crustacés. Certaines espèces sont des charognards, et certaines espèces de nemertini, comme Malacobdella grossa, ont également développé des relations commensalistes avec certains mollusques. Certaines espèces ont dévasté la pêche commerciale des palourdes et des crabes. Les Némertes n'ont pratiquement aucun prédateur et deux espèces sont vendues comme appâts pour poissons.
Morphologie
La taille des vers rubans varie de 1 cm à plusieurs mètres. Ils présentent une symétrie bilatérale et des propriétés contractiles remarquables. En raison de leur contractilité, ils peuvent modifier leur présentation morphologique en réponse à des signaux environnementaux. Les animaux du phylum Nemertea présentent une morphologie aplatie, c'est-à-dire qu'ils sont plats de l'avant vers l'arrière, comme un tube aplati. Les Nemertea sont des animaux mous et non segmentés (Figure\(\PageIndex{6}\)).

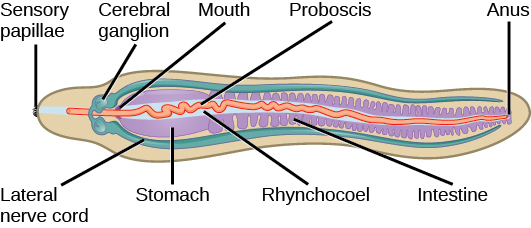
Une caractéristique unique de ce phylum est la présence d'une trompe enfermée dans un rhynchocèle. La trompe sert à capturer de la nourriture et peut être ornée de barbes chez certaines espèces. Le rhynchocoel est une cavité remplie de liquide qui s'étend de la tête jusqu'à près des deux tiers de la longueur de l'intestin chez ces animaux (Figure\(\PageIndex{7}\)). La trompe peut être étendue ou rétractée par le muscle rétracteur fixé à la paroi du rhynchocèle.
Système digestif
Les nemertini présentent un système digestif très développé. Une ouverture buccale ventrale par rapport au rhynchocèle mène à l'intestin antérieur, puis à l'intestin. L'intestin se présente sous forme de poches diverticulaires et se termine par un rectum qui s'ouvre par un anus. Les gonades sont entrecoupées des poches diverticulaires intestinales et s'ouvrent vers l'extérieur par les pores génitaux. Un système circulatoire consiste en une boucle fermée d'une paire de vaisseaux sanguins latéraux. Le système circulatoire est dérivé de la cavité coelomique de l'embryon. Certains animaux peuvent également avoir des vaisseaux interconnectés en plus des vaisseaux latéraux. Bien que ceux-ci soient appelés vaisseaux sanguins, puisqu'ils sont d'origine coelomique, le liquide circulatoire est incolore. Certaines espèces contiennent de l'hémoglobine ainsi que d'autres pigments jaunes ou verts. Les vaisseaux sanguins sont reliés au rhynchocèle. La circulation du liquide dans ces vaisseaux est facilitée par la contraction des muscles de la paroi corporelle. Une paire de protonéphridies, ou reins primitifs, est présente chez ces animaux pour faciliter l'osmorégulation. L'échange gazeux se produit à travers la peau des nemertini.
Système nerveux
Les nemertini ont un ganglion ou « cerveau » situé à l'extrémité antérieure entre la bouche et l'intestin antérieur, entourant le système digestif ainsi que le rhynchocèle. Un anneau de quatre masses nerveuses appelé « ganglions » compose le cerveau de ces animaux. Des cordons nerveux longitudinaux appariés émergent des ganglions cérébraux et s'étendent jusqu'à l'extrémité postérieure. Les ocelles ou points oculaires sont présents par paires, par multiples de deux dans la partie antérieure du corps. On suppose que les taches oculaires proviennent du tissu neural et non de l'épiderme.
Reproduction
Les animaux du phylum Nemertea présentent un dimorphisme sexuel, bien que les espèces d'eau douce puissent être hermaphrodites. Les ovules et les spermatozoïdes sont libérés dans l'eau et la fécondation se fait à l'extérieur. Le zygote se développe ensuite en larve planuliforme. Chez certaines espèces de némertines, une larve de pilidium peut se développer à l'intérieur du jeune ver, à partir d'une série de disques imaginaux. Cette forme larvaire, qui a la forme caractéristique d'une calotte de chevreuil, dévore les tissus du jeune ver pour survivre avant de se métamorphoser en une morphologie semblable à celle d'un adulte.
Phylum Mollusca
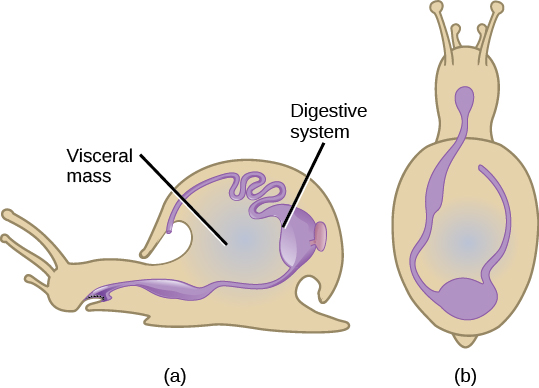
Le phylum Mollusca est le phylum prédominant dans les environnements marins. On estime que 23 pour cent de toutes les espèces marines connues sont des mollusques ; plus de 75 000 espèces ont été décrites, ce qui en fait le deuxième phylum d'animaux le plus diversifié. Le nom « mollusque » signifie un corps mou, puisque les premières descriptions des mollusques proviennent de l'observation de seiches non décortiquées. Les mollusques sont principalement un groupe d'animaux marins, mais on sait qu'ils habitent aussi bien les milieux d'eau douce que les milieux terrestres. Les mollusques présentent un large éventail de morphologies dans chaque classe et sous-classe, mais partagent quelques caractéristiques clés, notamment un pied musclé, une masse viscérale contenant des organes internes et un manteau qui peut ou non sécréter une coquille de carbonate de calcium (Figure\(\PageIndex{8}\)).

Exercice\(\PageIndex{1}\)
Laquelle des affirmations suivantes concernant l'anatomie d'un mollusque est fausse ?
- Les mollusques ont une radula pour broyer les aliments.
- Une glande digestive est connectée à l'estomac.
- Le tissu situé sous la coquille s'appelle le manteau.
- Le système digestif comprend un gésier, un estomac, une glande digestive et l'intestin.
- Réponse
-
d
Les mollusques ont un pied musclé, qui sert à la locomotion et à l'ancrage, et dont la forme et la fonction varient en fonction du type de mollusque étudié. Chez les mollusques décortiqués, ce pied est généralement de la même taille que l'ouverture de la coquille. Le pied est à la fois un organe rétractable et un organe extensible. Le pied est l'organe le plus ventral, tandis que le manteau est l'organe dorsal limite. Les mollusques sont des eucélomates, mais la cavité cœlomique est limitée à une cavité entourant le cœur chez les animaux adultes. La cavité du manteau se développe indépendamment de la cavité coelomique.
La masse viscérale est présente au-dessus du pied, dans la bosse viscérale. Cela inclut les systèmes digestif, nerveux, excréteur, reproducteur et respiratoire. Les espèces de mollusques exclusivement aquatiques ont des branchies pour respirer, tandis que certaines espèces terrestres ont des poumons pour respirer. De plus, un organe ressemblant à une langue appelé radula, qui porte une ornementation semblable à une dent chitineuse, est présent chez de nombreuses espèces et sert à déchiqueter ou à gratter les aliments. Le manteau (également connu sous le nom de pallium) est l'épiderme dorsal des mollusques ; les mollusques à coque sont spécialisés dans la sécrétion d'une coquille calcaire chitineuse et dure.
La plupart des mollusques sont des animaux dioïques et la fertilisation se fait de manière externe, bien que ce ne soit pas le cas chez les mollusques terrestres, tels que les escargots et les limaces, ou chez les céphalopodes. Chez certains mollusques, le zygote éclot et subit deux stades larvaires, trochophore et véligère, avant de devenir un jeune adulte ; les bivalves peuvent présenter un troisième stade larvaire, les glochidies.
Classification du phylum des mollusques
Le phylum Mollusca est un groupe très diversifié (85 000 espèces) composé principalement d'espèces marines. Les mollusques se présentent sous des formes très variées, allant de grands calmars et poulpes prédateurs, dont certains font preuve d'une grande intelligence, à des formes de pâturage aux coquilles finement sculptées et colorées. Ce phylum peut être divisé en sept classes : Aplacophora, monoplacophora, polyplacophora, bivalvia, gastropodes, céphalopodes et scaphopodes.
La classe Aplacophora (« ne portant aucune plaque ») comprend les animaux ressemblant à des vers que l'on trouve principalement dans les habitats marins benthiques. Ces animaux n'ont pas de carapace calcaire mais possèdent des spicules d'aragonite sur leur épiderme. Ils ont une cavité mantelale rudimentaire et sont dépourvus d'yeux, de tentacules et de néphridies (organes excréteurs). Les membres de la classe des monoplacophores (« portant une plaque ») possèdent une seule coque en forme de capuchon qui entoure le corps. La morphologie de la carapace et de l'animal sous-jacent peut varier de circulaire à ovale. Un système digestif en boucle, de multiples paires d'organes excréteurs, de nombreuses branchies et une paire de gonades sont présents chez ces animaux. Les monoplacophores étaient considérés comme éteints et n'étaient connus que par des fossiles jusqu'à la découverte de Neopilina galathaea en 1952. Aujourd'hui, les scientifiques ont identifié près de deux douzaines d'espèces existantes.
Les animaux de la classe des Polyplacophores (« porteurs de nombreuses plaques ») sont communément appelés « chitons » et portent une carapace plaquée en huit en forme d'armure (Figure\(\PageIndex{9}\)). Ces animaux ont un large pied ventral adapté à la succion des roches et d'autres substrats, et un manteau qui s'étend au-delà de la carapace sous la forme d'une ceinture. Des épines calcaires peuvent être présentes sur la ceinture pour se protéger des prédateurs. La respiration est facilitée par les cténidies (branchies) présentes sur la face ventrale. Ces animaux possèdent une radula modifiée pour être grattée. Le système nerveux est rudimentaire et seuls des ganglions buccaux ou « des joues » sont présents à l'extrémité antérieure. Les taches oculaires sont absentes chez ces animaux. Une seule paire de néphridies destinées à l'excrétion est présente.

La classe Bivalvia (« deux coquilles ») comprend les palourdes, les huîtres, les moules, les pétoncles et les panopes. Les membres de cette classe vivent dans des habitats marins et d'eau douce. Comme leur nom l'indique, les bivalves sont enfermés dans une paire de coquilles (les valves sont communément appelées « coquilles ») qui sont articulées à l'extrémité dorsale par des ligaments de la coquille ainsi que par des dents de la coquille (Figure\(\PageIndex{10}\)). La morphologie générale est aplatie latéralement et la région de la tête est peu développée. Les taches oculaires et les statocystes peuvent être absents chez certaines espèces. Comme ces animaux se nourrissent en suspension, il n'y a pas de radula dans cette classe de mollusques. La respiration est facilitée par une paire de cténidies, tandis que l'excrétion et l'osmorégulation sont provoquées par une paire de néphridies. Les bivalves possèdent souvent une grande cavité du manteau. Chez certaines espèces, les bords postérieurs du manteau peuvent fusionner pour former deux siphons qui servent à aspirer et à exsuder l'eau.

L'une des fonctions du manteau est de sécréter la coque. Certains bivalves, comme les huîtres et les moules, possèdent la capacité unique de sécréter et de déposer une nacre calcaire ou « nacre » autour de particules étrangères qui peuvent pénétrer dans la cavité du manteau. Cette propriété a été exploitée commercialement pour produire des perles.
Les animaux de la classe des gastropodes (« pied ventre ») comprennent des mollusques bien connus tels que les escargots, les limaces, les conques, les lièvres de mer et les papillons de mer. Les gastropodes comprennent des espèces à coquillages ainsi que des espèces à carapace réduite. Ces animaux sont asymétriques et présentent généralement une carapace enroulée (Figure\(\PageIndex{11}\)). Les coquilles peuvent être planospirales (comme un tuyau d'arrosage enroulé), communément observées chez les escargots de jardin, ou conispirales (comme un escalier en colimaçon), communément observées chez les conques marines.

La masse viscérale des espèces à coque présente une torsion autour de l'axe perpendiculaire au centre du pied, qui est la principale caractéristique de ce groupe, ainsi qu'un pied modifié pour ramper (Figure\(\PageIndex{12}\)). La plupart des gastéropodes ont une tête avec des tentacules, des yeux et un style. Une radula complexe est utilisée par le système digestif et facilite l'ingestion de nourriture. Les yeux peuvent être absents chez certaines espèces de gastéropodes. La cavité du manteau renferme les cténidies ainsi qu'une paire de néphridies.
Connexion quotidienne : le venin d'escargot peut-il être utilisé comme analgésique pharmacologique ?
Les escargots marins du genre Conus (Figure\(\PageIndex{13}\)) attaquent leurs proies avec une piqûre venimeuse. La toxine libérée, connue sous le nom de conotoxine, est un peptide doté de liaisons disulfures internes. Les conotoxines peuvent provoquer la paralysie chez l'homme, ce qui indique que cette toxine attaque des cibles neurologiques. Il a été démontré que certaines conotoxines bloquent les canaux ioniques neuronaux. Ces résultats ont amené les chercheurs à étudier les conotoxines pour d'éventuelles applications médicales.
Les conotoxines constituent un domaine passionnant de développement pharmacologique potentiel, car ces peptides peuvent être modifiés et utilisés dans des conditions médicales spécifiques pour inhiber l'activité de neurones spécifiques. Par exemple, ces toxines peuvent être utilisées pour provoquer la paralysie des muscles dans des applications de santé spécifiques, de la même manière que l'utilisation de la toxine botulique. L'ensemble du spectre des conotoxines, ainsi que leurs mécanismes d'action, n'étant pas complètement connus, l'étude de leurs applications potentielles en est encore à ses balbutiements. À ce jour, la plupart des recherches se sont concentrées sur leur utilisation pour traiter les maladies neurologiques. Ils se sont également révélés efficaces pour soulager la douleur chronique et la douleur associée à des affections telles que la sciatique et le zona. L'étude et l'utilisation de biotoxines, des toxines dérivées d'organismes vivants, constituent un excellent exemple de l'application de la science biologique à la médecine moderne.

La classe des céphalopodes (animaux « pieds de tête ») comprend les poulpes, les calmars, les seiches et les nautilus. Les céphalopodes sont une classe d'animaux porteurs de coquillages ainsi que de mollusques à carapace réduite. Ils présentent une coloration vive, généralement observée dans les calmars et les poulpes, qui sont utilisés pour le camouflage. Tous les animaux de cette classe sont des prédateurs carnivores et ont des mâchoires en forme de bec à l'extrémité antérieure. Tous les céphalopodes présentent la présence d'un système nerveux très développé ainsi que d'yeux, ainsi que d'un système circulatoire fermé. Le pied est lobé et transformé en tentacules et en entonnoir, qui leur sert de mode de locomotion. Des drageons sont présents sur les tentacules des poulpes et des calmars. Les cténidies sont enfermées dans une grande cavité du manteau et sont desservies par de gros vaisseaux sanguins, chacun étant associé à son propre cœur ; le manteau possède des siphonophores qui facilitent l'échange d'eau.
La locomotion des céphalopodes est facilitée par l'éjection d'un jet d'eau pour la propulsion. C'est ce qu'on appelle la propulsion « à réaction ». Une paire de néphridies est présente dans la cavité du manteau. Un dimorphisme sexuel est observé chez cette classe d'animaux. Les membres d'une espèce s'accouplent et la femelle pond ensuite ses œufs dans une niche isolée et protégée. Les femelles de certaines espèces s'occupent des œufs pendant une période prolongée et peuvent finir par mourir pendant cette période. Les céphalopodes tels que les calmars et les poulpes produisent également du sépia ou une encre foncée, que l'on projette sur un prédateur pour l'aider à s'échapper rapidement.
La reproduction chez les céphalopodes est différente de celle des autres mollusques en ce sens que l'œuf éclot pour donner naissance à un adulte juvénile sans passer par les stades larvaires trochophores et véligères.
Chez le Nautilus spp., qui porte des coquillages, la coque en spirale est à chambres multiples. Ces chambres sont remplies de gaz ou d'eau pour réguler la flottabilité. La structure de la coquille des calmars et des seiches est réduite et se présente à l'intérieur sous la forme d'un enclos à calmars et d'un os de seiche, respectivement. Des exemples sont présentés dans la figure\(\PageIndex{14}\).

Figure\(\PageIndex{14}\) : Le (a) nautilus, (b) la seiche géante, (c) le calmar de récif et (d) le poulpe à anneaux bleus appartiennent tous à la classe des céphalopodes. (crédit a : modification d'une œuvre par J. Baecker ; crédit b : modification d'une œuvre par Adrian Mohedano ; crédit c : modification d'une œuvre par Silke Baron ; crédit d : modification d'une œuvre par Angell Williams)
Les membres de la classe des scaphopodes (« pieds de bateau ») sont communément appelés « défenses » ou « coquilles dentaires », comme en témoigne l'examen de Dentalium, l'un des rares genres de scaphopodes restants (Figure\(\PageIndex{15}\)). Les scaphopodes sont généralement enfouis dans le sable, l'ouverture antérieure étant exposée à l'eau. Ces animaux portent une seule carapace conique dont les deux extrémités sont ouvertes. La tête est rudimentaire et dépasse de l'extrémité postérieure de la coquille. Ces animaux ne possèdent pas d'yeux, mais ils ont une radula, ainsi qu'un pied modifié en tentacules à extrémité bulbeuse, appelés captacles. Les captacules servent à attraper et à manipuler les proies. Les cténidies sont absentes chez ces animaux.
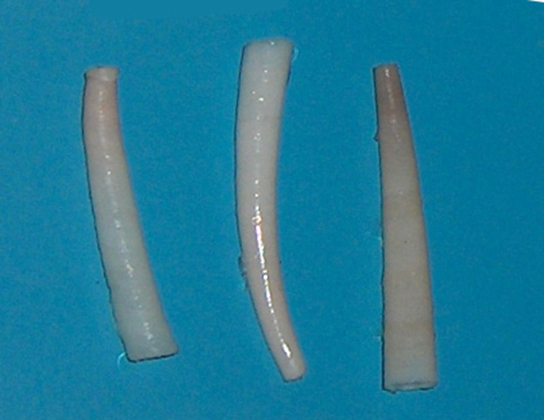
Figure\(\PageIndex{15}\) : Antalis vulgaris montre la forme classique des Dentaliidae qui donne à ces animaux leur nom commun de « coquille de défense ». (crédit : Georges Jansoone)
Phylum Annelida
Le phylum Annelida comprend des vers segmentés. Ces animaux vivent dans des habitats marins, terrestres et d'eau douce, mais la présence d'eau ou d'humidité est un facteur critique pour leur survie, en particulier dans les habitats terrestres. Le nom du phylum est dérivé du mot latin annellus, qui signifie un petit anneau. Les animaux de ce phylum présentent des symbioses parasitaires et commensales avec d'autres espèces de leur habitat. Environ 16 500 espèces ont été décrites dans le phylum Annelida. Le phylum comprend des vers de terre, des vers polychètes et des sangsues. Les annélides présentent un développement protostomique au stade embryonnaire et sont souvent appelées « vers segmentés » en raison de leur caractéristique clé, le métamérisme, ou véritable segmentation.
Morphologie
Les annélides présentent une symétrie bilatérale et leur morphologie générale ressemble à des vers. Les annélides ont un plan corporel segmenté dans lequel les caractéristiques morphologiques internes et externes se répètent dans chaque segment du corps. Le métamérisme permet aux animaux de grossir en ajoutant des « compartiments » tout en rendant leurs mouvements plus efficaces. On pense que ce métamérisme provient de cellules téloblastiques identiques au stade embryonnaire, qui donnent naissance à des structures mésodermiques identiques. L'ensemble du corps peut être divisé en tête, corps et pygidium (ou queue). Le clitellum est une structure reproductrice qui produit du mucus qui facilite le transfert des spermatozoïdes et donne naissance à un cochon dans lequel se produit la fécondation ; il apparaît sous la forme d'une bande fusionnée dans le tiers antérieur de l'animal (Figure\(\PageIndex{16}\)).

Figure\(\PageIndex{16}\) : Le clitellum, vu ici comme un segment protubérant de coloration différente de celle du reste du corps, est une structure qui favorise la reproduction des annélides. (crédit : Rob Hille)
anatomie
L'épiderme est protégé par une cuticule externe acellulaire, mais celle-ci est beaucoup plus fine que la cuticule des ecdysozoaires et ne nécessite pas de perte périodique pour sa croissance. Les muscles circulaires et longitudinaux sont situés à l'intérieur de l'épiderme. Des extensions chitineuses ressemblant à des poils, ancrées dans l'épiderme et faisant saillie à partir de la cuticule, appelées soetes/chètes, sont présentes dans chaque segment. Les annélides montrent la présence d'un véritable coelome, dérivé du mésoderme embryonnaire et de la protostomie. Ce sont donc les vers les plus avancés. Les vers de terre (oligochètes) ont un système digestif bien développé et complet, avec une bouche, un pharynx musculaire, un œsophage, une plante et un gésier. Le gésier mène à l'intestin et se termine par une ouverture anale. Une vue en coupe transversale d'un segment du corps d'un ver de terre (un annélide de type terrestre) est présentée sur la figure\(\PageIndex{17}\) ; chaque segment est limité par un septum membraneux qui divise la cavité coelomique en une série de compartiments.
Les annélides possèdent un système circulatoire fermé composé de vaisseaux sanguins dorsaux et ventraux parallèles au tube digestif ainsi que de capillaires qui desservent les tissus individuels. De plus, ces vaisseaux sont reliés par des boucles transversales dans chaque segment. Ces animaux n'ont pas de système respiratoire bien développé et les échanges gazeux se produisent à travers la surface humide du corps. L'excrétion est facilitée par une paire de métanéphridies (type de « rein » primitif constitué d'un tubule alambiqué et d'un entonnoir cilié ouvert) présentes dans tous les segments situés vers la face ventrale. Les annélides présentent un système nerveux bien développé avec un anneau nerveux composé de ganglions fusionnés autour du pharynx. Le cordon nerveux est en position ventrale et porte des ganglions ou des ganglions élargis dans chaque segment.
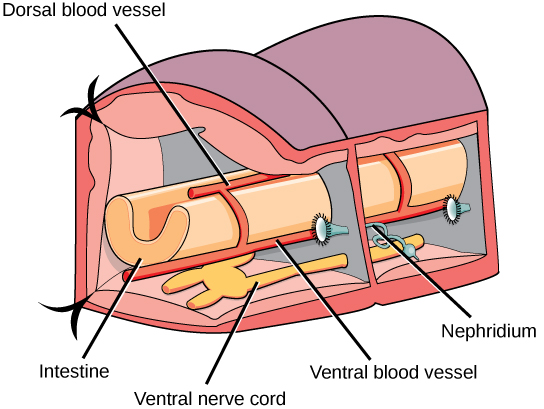
Les annélides peuvent être monoïques avec des gonades permanentes (comme chez les vers de terre et les sangsues) ou dioïques avec des gonades temporaires ou saisonnières qui se développent (comme chez les polychètes). Cependant, la fertilisation croisée est préférée chez les animaux hermaphrodites. Ces animaux peuvent également présenter un hermaphrodisme simultané et participer à un échange simultané de spermatozoïdes lorsqu'ils sont alignés pour la copulation.
Classification du phylum annélida
Le phylum Annelida contient la classe des Polychètes (les polychètes) et la classe des Oligochètes (les vers de terre, les sangsues et les espèces apparentées).
Les vers de terre sont les membres les plus abondants de la classe des oligochètes, qui se distinguent par la présence du clitellum et d'un petit nombre de chètes réduites (« oligo- = « peu » ; -chaetae = « poils »). Le nombre et la taille des chètes sont fortement diminués dans les Oligochètes par rapport aux polychètes (polychètes = plusieurs, chètes = poils). Les nombreux chètes des polychètes sont également disposés dans des appendices charnus, plats et appariés qui font saillie à partir de chaque segment appelé parapodie, qui peuvent être spécialisés pour différentes fonctions chez les polychates. La sous-classe Hirudinea comprend des sangsues telles que Hirudo medicinalis et Hemiclepsis marginata. La classe Oligochaeta comprend la sous-classe Hirudinia et la sous-classe Brachiobdella. Une différence significative entre les sangsues et les autres annélides est le développement de drageons aux extrémités antérieure et postérieure et l'absence de chaetes. De plus, la segmentation de la paroi corporelle peut ne pas correspondre à la segmentation interne de la cavité coelomique. Cette adaptation peut aider les sangsues à s'allonger lorsqu'elles ingèrent de grandes quantités de sang provenant de vertébrés hôtes. La sous-classe Brachiobdella comprend des espèces telles que Branchiobdella balcanica sketi et Branchiobdella astaci, des vers qui ressemblent à des sangsues ainsi qu'à des oligochètes.

Figure\(\PageIndex{18}\) : (a) le ver de terre, (b) la sangsue et (c) le plumeau sont tous des annélides. (crédit a : modification d'une œuvre de S. Shepherd ; crédit b : modification d'une œuvre de « Sarah G... ») /Flickr ; crédit c : modification de l'œuvre de Chris Gotschalk, NOAA)
Résumé
Le phylum Annelida comprend des animaux vermiformes segmentés. La segmentation est également observée dans l'anatomie interne, ce que l'on appelle le métamérisme. Les annélides sont des protostomes. Ces animaux ont des systèmes neuronal et digestif bien développés. Certaines espèces portent une bande spécialisée de segments appelée clitellum. Les annélides montrent la présence de nombreuses projections chitineuses appelées chètes, et les polychètes possèdent des parapodies. Les meuniers sont vus dans l'ordre Hirudinea. Les stratégies de reproduction incluent le dimorphisme sexuel, l'hermaphrodisme et l'hermaphrodisme en série. La segmentation interne est absente dans la classe Hirudinea.
Les ascaris sont des animaux acéolomés et triploblastiques. Ils sont dépourvus de systèmes circulatoire et respiratoire et ont un système excréteur rudimentaire. Ce système digestif est incomplet chez la plupart des espèces. Il existe quatre classes traditionnelles de vers plats : les turbellaires vivant en grande partie à l'état libre, les monogéniques ectoparasites et les trématodes et cestodes endoparasites. Les trématodes ont des cycles de vie complexes impliquant un mollusque hôte secondaire et un hôte principal dans lequel se produit la reproduction sexuée. Les cestodes, ou ténias, infectent le système digestif des hôtes vertébrés primaires.
Les rotifères sont des organismes microscopiques, multicellulaires, principalement aquatiques, qui font actuellement l'objet d'une révision taxonomique. Le groupe se caractérise par la structure ciliée rotative en forme de roue, la couronne, qui se trouve sur sa tête. La mastax ou pharynx à mâchoires est une autre structure propre à ce groupe d'organismes.
Les nemertini sont les eucoélomates les plus simples. Ces animaux en forme de ruban portent une trompe spécialisée enfermée dans un rhynchocèle. Le développement d'un système circulatoire fermé dérivé du coelom constitue une différence significative observée chez cette espèce par rapport aux autres phylums pseudocélomates. Les systèmes digestif, nerveux et excréteur sont plus développés chez les nemertini que chez les phylums moins avancés. Le développement embryonnaire des vers némertins se fait par un stade larvaire planuliforme.
Le phylum Mollusca est un vaste groupe d'invertébrés marins. Les mollusques présentent diverses variations morphologiques au sein du phylum. Ce phylum se distingue également par le fait que certains membres présentent une coque calcaire comme moyen de protection externe. Certains mollusques ont développé une carapace réduite. Les mollusques sont des protostomes. L'épiderme dorsal des mollusques est modifié pour former le manteau, qui entoure la cavité du manteau et les organes viscéraux. Cette cavité est bien distincte de la cavité cœlomique qui, chez l'animal adulte, entoure le cœur. La respiration est facilitée par des branchies appelées cténidies. Une langue à dents chitineuses appelée radula est présente chez la plupart des mollusques. Le développement précoce de certaines espèces se fait par l'intermédiaire de deux stades larvaires : trochophore et véligère. Le dimorphisme sexuel est la stratégie sexuelle prédominante dans ce phylum. Les mollusques peuvent être divisés en sept classes, chacune ayant des caractéristiques morphologiques distinctes.
Lexique
- Annélide
- phylum d'animaux vermiformes avec métamérisme
- captacule
- projection semblable à un tentacule présente dans les défenses et les coquilles pour attraper des proies
- clitellum
- bande spécialisée de segments fusionnés, qui facilite la reproduction
- conispiral
- forme de coque enroulée autour d'un axe horizontal
- couronne
- structure en forme de roue sur la partie antérieure du rotifère qui contient les cils et déplace la nourriture et l'eau vers la bouche
- cténidium
- structure branchiale spécialisée chez les mollusques
- manteau
- (également, pallium) épiderme spécialisé qui entoure tous les organes viscéraux et sécrète des coquilles
- mastax
- pharynx à mâchoires propre aux rotifères
- métamérisme
- série de structures corporelles similaires à l'intérieur et à l'extérieur, telles que des segments
- Mollusques
- phylum de protostomes à corps mous et sans segmentation
- nacre
- sécrétion calcaire produite par les bivalves pour recouvrir la face interne des coquilles et pour recouvrir les particules qui s'y introduisent
- Thé Nemertea
- phylum de protostomes aplatis dorsoventralement connus sous le nom de vers rubans
- parapodium
- appendice charnu, plat, qui fait saillie par paires à partir de chaque segment de polychètes
- pilidium
- forme larvaire présente chez certaines espèces de némertines
- planospiral
- forme de coque enroulée autour d'un axe vertical
- planuliforme
- forme larvaire trouvée dans le phylum Nemertea
- radula
- orgue en forme de langue orné d'ornements chitineux
- rhynchocoel
- cavité présente au-dessus de la bouche qui abrite la trompe
- schizocoelom
- coelome formé par des groupes de cellules qui se séparent de la couche endodermique
- seta/chaeta
- projection chitineuse à partir de la cuticule
- trochophore
- premier des deux stades larvaires chez les mollusques
- veliger
- deuxième des deux stades larvaires chez les mollusques