
23.1 : Origines eucaryotes
- Page ID
- 189849

Compétences à développer
- Énumérer les caractéristiques unificatrices des eucaryotes
- Décrivez ce que les scientifiques savent sur l'origine des eucaryotes en se basant sur le dernier ancêtre commun
- Expliquer la théorie endosymbiotique
Les êtres vivants se répartissent en trois grands groupes : les archées, les bactéries et les eucaryas. Les deux premiers contiennent des cellules procaryotes et le troisième contient tous les eucaryotes. Des fossiles relativement rares sont disponibles pour aider à discerner à quoi ressemblaient les premiers membres de chacune de ces lignées. Il est donc possible que tous les événements qui ont mené au dernier ancêtre commun des eucaryotes existants restent inconnus. Cependant, la biologie comparée des organismes existants et le nombre limité de fossiles fournissent un aperçu de l'histoire de l'Eucarya.
Les premiers fossiles découverts semblent être des bactéries, probablement des cyanobactéries. Ils ont environ 3,5 milliards d'années et sont reconnaissables en raison de leur structure relativement complexe et, pour les procaryotes, de leurs cellules relativement grandes. La plupart des autres procaryotes possèdent de petites cellules de 1 ou 2 µm et seraient difficiles à identifier sous forme de fossiles. La plupart des eucaryotes vivants ont des cellules mesurant 10 µm ou plus. Des structures de cette taille, qui peuvent être des fossiles, apparaissent dans les archives géologiques il y a environ 2,1 milliards d'années.
Caractéristiques des eucaryotes
Les données provenant de ces fossiles ont conduit des biologistes comparés à la conclusion que les eucaryotes vivants sont tous les descendants d'un seul ancêtre commun. La cartographie des caractéristiques trouvées dans tous les principaux groupes d'eucaryotes révèle que les caractéristiques suivantes devaient être présentes chez le dernier ancêtre commun, car ces caractéristiques sont présentes chez au moins certains des membres de chaque lignée principale.
- Cellules dont les noyaux sont entourés d'une enveloppe nucléaire avec des pores nucléaires. C'est la seule caractéristique qui est à la fois nécessaire et suffisante pour définir un organisme comme un eucaryote. Tous les eucaryotes existants possèdent des cellules dotées de noyaux.
- Mitochondries. Certains eucaryotes existants ont des restes de mitochondries très réduits dans leurs cellules, tandis que d'autres membres de leur lignée ont des mitochondries « typiques ».
- Cytosquelette contenant les composants structuraux et de motilité appelés microfilaments et microtubules d'actine. Tous les eucaryotes existants possèdent ces éléments cytosquelettiques.
- Flagelles et cils, organites associés à la motilité cellulaire. Certains eucaryotes existants n'ont pas de flagelles et/ou de cils, mais ils descendent d'ancêtres qui les possédaient.
- Chromosomes, chacun étant constitué d'une molécule d'ADN linéaire enroulée autour de protéines basiques (alcalines) appelées histones. Les quelques eucaryotes dont les chromosomes sont dépourvus d'histones ont clairement évolué à partir d'ancêtres qui en étaient porteurs.
- La mitose est un processus de division nucléaire par lequel les chromosomes répliqués sont divisés et séparés à l'aide d'éléments du cytosquelette. La mitose est universellement présente chez les eucaryotes.
- Le sexe est un processus de recombinaison génétique propre aux eucaryotes au cours duquel les noyaux diploïdes à un stade du cycle vital subissent une méiose pour produire des noyaux haploïdes et une caryogamie subséquente, stade où deux noyaux haploïdes fusionnent pour créer un noyau zygote diploïde.
- Les membres de toutes les lignées principales possèdent des parois cellulaires, et il serait raisonnable de conclure que le dernier ancêtre commun pourrait fabriquer des parois cellulaires à un stade ou à un autre de son cycle de vie. Cependant, on ne connaît pas suffisamment les parois cellulaires des eucaryotes et leur développement pour connaître le degré d'homologie qui existe entre eux. Si le dernier ancêtre commun a pu créer des parois cellulaires, il est clair que cette capacité a dû être perdue dans de nombreux groupes.
Endosymbiose et évolution des eucaryotes
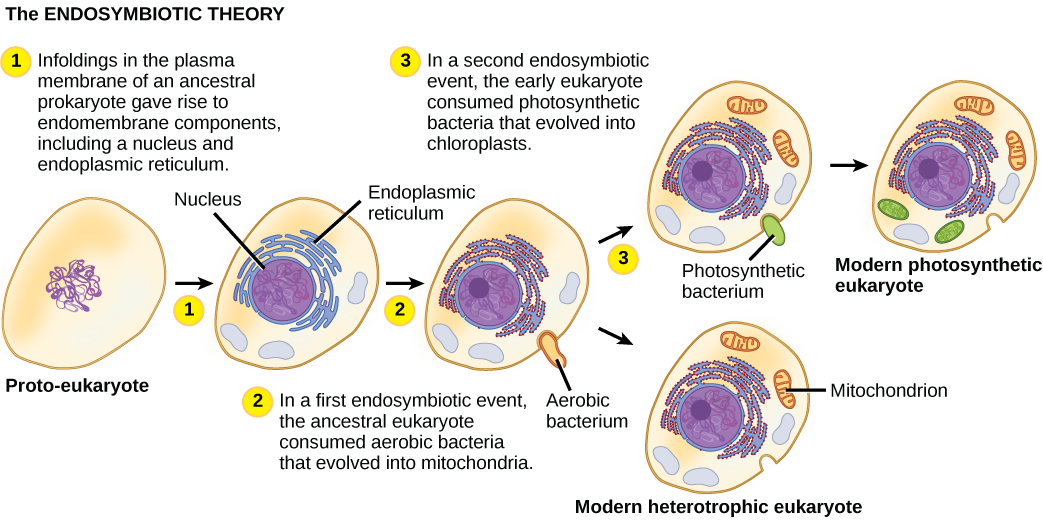
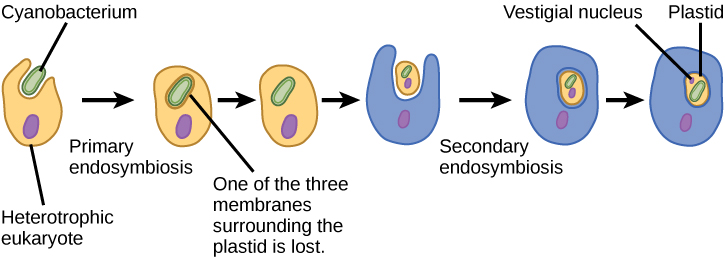
Pour bien comprendre les organismes eucaryotes, il faut comprendre que tous les eucaryotes existants sont les descendants d'un organisme chimère composé d'une cellule hôte et de la ou des cellules d'une alpha-protéobactérie qui a « élu domicile » à l'intérieur de celui-ci. Ce thème majeur à l'origine des eucaryotes est connu sous le nom d'endosymbiose, une cellule en engloutissant une autre de telle sorte que la cellule engloutie survit et que les deux cellules en bénéficient. Sur de nombreuses générations, une relation symbiotique peut donner naissance à deux organismes qui dépendent tellement l'un de l'autre qu'aucun d'entre eux ne pourrait survivre seul. Les événements endosymbiotiques ont probablement contribué à l'origine du dernier ancêtre commun des eucaryotes actuels et à la diversification ultérieure de certaines lignées d'eucaryotes (Figure\(\PageIndex{4}\)). Avant d'expliquer cela plus en détail, il est nécessaire de considérer le métabolisme des procaryotes.
Métabolisme procaryote
De nombreux processus métaboliques importants sont apparus chez les procaryotes, et certains d'entre eux, tels que la fixation de l'azote, ne sont jamais observés chez les eucaryotes. Le processus de respiration aérobie se retrouve dans toutes les principales lignées d'eucaryotes et est localisé dans les mitochondries. La respiration aérobie est également présente dans de nombreuses lignées de procaryotes, mais elle n'est pas présente chez toutes, et de nombreuses preuves suggèrent que ces procaryotes anaérobies n'ont jamais exercé de respiration aérobie, pas plus que leurs ancêtres.
Alors que l'atmosphère actuelle contient environ un cinquième de la molécule d'oxygène (O 2), des preuves géologiques montrent qu'elle était initialement dépourvue d'O 2. Sans oxygène, on ne s'attendrait pas à une respiration aérobie, et les êtres vivants auraient plutôt compté sur la fermentation. À un moment donné, il y a environ 3,5 milliards d'années, certains procaryotes ont commencé à utiliser l'énergie du soleil pour activer des processus anabolisants qui réduisent le dioxyde de carbone pour former des composés organiques. C'est-à-dire qu'ils ont développé la capacité de photosynthèse. L'hydrogène, dérivé de diverses sources, a été capturé à l'aide de réactions alimentées par la lumière afin de réduire le dioxyde de carbone fixe dans le cycle de Calvin. Le groupe de bactéries à Gram négatif à l'origine des cyanobactéries a utilisé de l'eau comme source d'hydrogène et a libéré de l'O 2 sous forme de déchets.
Finalement, la quantité d'oxygène photosynthétique s'est accumulée dans certains environnements à des niveaux présentant un risque pour les organismes vivants, car elle peut endommager de nombreux composés organiques. Divers processus métaboliques se sont développés pour protéger les organismes de l'oxygène, dont l'un, la respiration aérobie, a également généré des niveaux élevés d'ATP. Elle est devenue largement présente chez les procaryotes, y compris dans un groupe que nous appelons aujourd'hui les alpha-protéobactéries. Les organismes qui n'acquéraient pas de respiration aérobie devaient rester dans des environnements exempts d'oxygène. À l'origine, les environnements riches en oxygène étaient probablement localisés autour d'endroits où les cyanobactéries étaient actives, mais il y a environ 2 milliards d'années, des preuves géologiques montrent que l'oxygène s'accumulait à des concentrations plus élevées dans l'atmosphère. Des niveaux d'oxygène similaires aux niveaux actuels n'ont été observés qu'au cours des 700 derniers millions d'années.
Rappelons que les premiers fossiles que nous croyons être des eucaryotes datent d'environ 2 milliards d'années, ils sont donc apparus alors que les niveaux d'oxygène augmentaient. Rappelons également que tous les eucaryotes existants descendaient d'un ancêtre doté de mitochondries. Ces organites ont été observés pour la première fois par des microscopistes optiques à la fin des années 1800, où ils semblaient être des structures en forme de vers qui semblaient se déplacer dans la cellule. Certains des premiers observateurs ont suggéré qu'il s'agissait peut-être de bactéries vivant à l'intérieur des cellules hôtes, mais ces hypothèses sont restées inconnues ou rejetées dans la plupart des communautés scientifiques.
Théorie endosymbiotique
Au fur et à mesure que la biologie cellulaire s'est développée au XXe siècle, il est devenu évident que les mitochondries étaient les organites responsables de la production d'ATP par respiration aérobie. Dans les années 1960, la biologiste américaine Lynn Margulis a développé une théorie endosymbiotique, selon laquelle les eucaryotes peuvent être le produit d'une cellule qui en engloutit une autre, d'une vie dans une autre et d'une évolution au fil du temps jusqu'à ce que les cellules séparées ne soient plus reconnaissables en tant que telles. En 1967, Margulis a introduit de nouveaux travaux sur la théorie et a étayé ses découvertes par des preuves microbiologiques. Bien que le travail de Margulis ait d'abord rencontré des résistances, cette hypothèse autrefois révolutionnaire est aujourd'hui largement (mais pas complètement) acceptée, les travaux progressant pour découvrir les étapes de ce processus évolutif et les principaux acteurs impliqués. Il reste encore beaucoup à découvrir sur l'origine des cellules qui constituent aujourd'hui les cellules de tous les eucaryotes vivants.
Dans l'ensemble, il est devenu évident que bon nombre de nos gènes nucléaires et de la machinerie moléculaire responsable de la réplication et de l'expression semblent étroitement liés à ceux d'Archaea. D'autre part, les organites métaboliques et les gènes responsables de nombreux processus de récupération d'énergie avaient leur origine dans des bactéries. Il reste encore beaucoup à clarifier sur la façon dont cette relation s'est produite ; il s'agit toujours d'un domaine de découverte passionnant en biologie. Par exemple, on ne sait pas si l'événement endosymbiotique à l'origine des mitochondries s'est produit avant ou après que la cellule hôte ait eu un noyau. Ces organismes seraient parmi les précurseurs éteints du dernier ancêtre commun des eucaryotes.
Mitochondries
L'une des principales caractéristiques qui distinguent les procaryotes des eucaryotes est la présence de mitochondries. Les cellules eucaryotes peuvent contenir entre un et plusieurs milliers de mitochondries, selon le niveau de consommation énergétique de la cellule. Chaque mitochondrie mesure 1 à 10 micromètres ou plus de longueur et existe dans la cellule sous la forme d'un organite qui peut être ovoïde, en forme de ver ou de ramification complexe (Figure\(\PageIndex{1}\)). Les mitochondries proviennent de la division des mitochondries existantes ; elles peuvent fusionner entre elles et se déplacer à l'intérieur de la cellule par des interactions avec le cytosquelette. Cependant, les mitochondries ne peuvent pas survivre à l'extérieur de la cellule. Au fur et à mesure que l'atmosphère était oxygénée par la photosynthèse et que des procaryotes aérobies évoluaient avec succès, les preuves suggèrent qu'une cellule ancestrale présentant une certaine compartimentation membranaire a englouti un procaryote aérobie libre, en particulier une alpha-protéobactérie, donnant ainsi à la cellule hôte la capacité d'utiliser de l'oxygène pour libérer l'énergie stockée dans les nutriments. Les alpha-protéobactéries sont un vaste groupe de bactéries qui comprend des espèces symbiotiques avec les plantes, des organismes pathogènes qui peuvent infecter les humains par le biais des tiques et de nombreuses espèces libres qui utilisent la lumière comme source d'énergie. Plusieurs sources de données soutiennent que les mitochondries sont dérivées de cet événement endosymbiotique. La plupart des mitochondries ont la forme d'alpha-protéobactéries et sont entourées de deux membranes, ce qui se produit lorsqu'un organisme lié à la membrane est englouti dans une vacuole par un autre organisme lié à la membrane. La membrane interne mitochondriale est étendue et comporte d'importants repliements appelés crêtes qui ressemblent à la surface externe texturée des alpha-protéobactéries. La matrice et la membrane interne sont riches en enzymes nécessaires à la respiration aérobie.
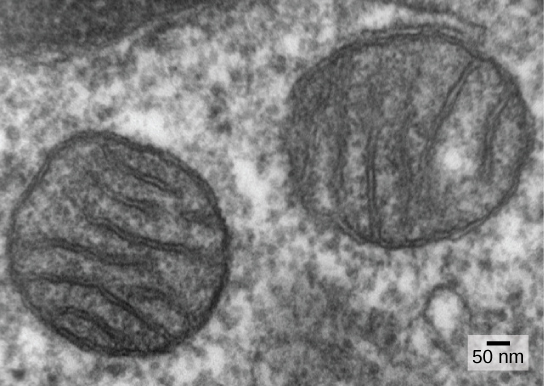
Les mitochondries se divisent indépendamment par un processus qui ressemble à une fission binaire chez les procaryotes. Plus précisément, les mitochondries ne sont pas formées de toutes pièces (de novo) par la cellule eucaryote ; elles s'y reproduisent et se distribuent avec le cytoplasme lorsqu'une cellule se divise ou que deux cellules fusionnent. Par conséquent, bien que ces organites soient hautement intégrés dans la cellule eucaryote, ils se reproduisent toujours comme s'ils étaient des organismes indépendants au sein de la cellule. Cependant, leur reproduction est synchronisée avec l'activité et la division de la cellule. Les mitochondries possèdent leur propre chromosome d'ADN (généralement) circulaire qui est stabilisé par des attaches à la membrane interne et porte des gènes similaires aux gènes exprimés par les alpha-protéobactéries. Les mitochondries possèdent également des ribosomes spéciaux et des ARN de transfert qui ressemblent à ces composants chez les procaryotes. Ces caractéristiques confirment toutes que les mitochondries étaient autrefois des procaryotes libres.
Les mitochondries qui assurent la respiration aérobie possèdent leur propre génome, avec des gènes similaires à ceux des alpha-protéobactéries. Cependant, de nombreux gènes des protéines respiratoires se trouvent dans le noyau. Lorsque ces gènes sont comparés à ceux d'autres organismes, ils semblent être d'origine alpha-protéobactérienne. De plus, dans certains groupes eucaryotes, ces gènes se trouvent dans les mitochondries, tandis que dans d'autres groupes, ils se trouvent dans le noyau. Cela a été interprété comme une preuve que des gènes ont été transférés du chromosome de l'endosymbiote au génome de l'hôte. Cette perte de gènes par l'endosymbiote explique probablement pourquoi les mitochondries ne peuvent pas vivre sans hôte.
Certains eucaryotes vivants sont anaérobies et ne peuvent pas survivre en présence d'une trop grande quantité d'oxygène. Certains semblent dépourvus d'organites qui pourraient être reconnus comme des mitochondries. Entre les années 1970 et le début des années 1990, de nombreux biologistes ont suggéré que certains de ces eucaryotes descendaient d'ancêtres dont les lignées avaient divergé de la lignée des eucaryotes contenant des mitochondries avant l'apparition de l'endosymbiose. Cependant, des résultats ultérieurs suggèrent que des organites réduits se trouvent dans la plupart, sinon dans la totalité, des eucaryotes anaérobies, et que tous les eucaryotes semblent porter certains gènes dans leur noyau qui sont d'origine mitochondriale. Outre la production aérobie d'ATP, les mitochondries ont plusieurs autres fonctions métaboliques. L'une de ces fonctions est de générer des agrégats de fer et de soufre qui sont des cofacteurs importants de nombreuses enzymes. Ces fonctions sont souvent associées à la réduction des organites dérivés des mitochondries chez les eucaryotes anaérobies. Par conséquent, la plupart des biologistes admettent que le dernier ancêtre commun des eucaryotes était la mitochondrie.
Plastiques
Certains groupes d'eucaryotes sont photosynthétiques. Leurs cellules contiennent, en plus des organites eucaryotes standard, un autre type d'organite appelé plaste. Lorsque ces cellules effectuent la photosynthèse, leurs plastes sont riches en chlorophylle a, un pigment, et en une gamme d'autres pigments, appelés pigments accessoires, qui participent à la récupération de l'énergie de la lumière. Les plastes photosynthétiques sont appelés chloroplastes (Figure\(\PageIndex{2}\)).
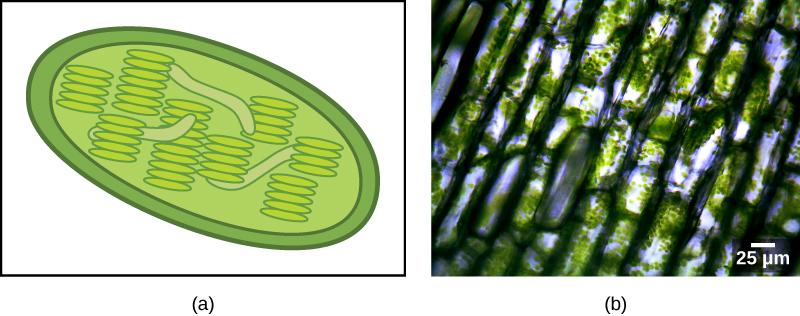
Comme les mitochondries, les plastes semblent avoir une origine endosymbiotique. Cette hypothèse a également été défendue par Lynn Margulis. Les plastes sont dérivés de cyanobactéries qui vivaient à l'intérieur des cellules d'un eucaryote hétérotrophe, aérobie et ancestral. C'est ce que l'on appelle l'endosymbiose primaire, et les plastes d'origine primaire sont entourés de deux membranes. La meilleure preuve est que cela s'est produit deux fois dans l'histoire des eucaryotes. Dans un cas, l'ancêtre commun de la principale lignée/supergroupe des Archaeplastida s'est attaqué à un endosymbiote cyanobactérien ; dans l'autre, l'ancêtre du petit taxon rhizarien amiboïde, Paulinella, s'est attaqué à un endosymbiote cyanobactérien différent. Presque tous les eucaryotes photosynthétiques sont issus du premier événement, et seules quelques espèces sont dérivées de l'autre.
Les cyanobactéries sont un groupe de bactéries à Gram négatif présentant toutes les structures classiques du groupe. Cependant, contrairement à la plupart des procaryotes, ils possèdent de vastes sacs liés à la membrane interne appelés thylakoïdes. La chlorophylle est un composant de ces membranes, tout comme de nombreuses protéines des réactions lumineuses de photosynthèse. Les cyanobactéries possèdent également la paroi du peptidoglycane et la couche de lipopolysaccharides associées aux bactéries à Gram négatif.
Les chloroplastes d'origine primaire possèdent des thylakoïdes, un chromosome d'ADN circulaire et des ribosomes similaires à ceux des cyanobactéries. Chaque chloroplaste est entouré de deux membranes. Dans le groupe des Archaeplastides appelé glaucophytes et chez Paulinella, une fine couche de peptidoglycane est présente entre les membranes externe et interne des plastides. Tous les autres plastes sont dépourvus de cette paroi cyanobactérienne relique. On pense que la membrane externe entourant le plaste provient de la vacuole de l'hôte, et que la membrane interne est dérivée de la membrane plasmique du symbiote.
Comme dans le cas des mitochondries, il existe également des preuves solides que de nombreux gènes de l'endosymbiote ont été transférés au noyau. Les plastes, comme les mitochondries, ne peuvent pas vivre indépendamment en dehors de l'hôte. De plus, comme les mitochondries, les plastes sont dérivés de la division d'autres plastes et ne sont jamais construits à partir de zéro. Les chercheurs ont suggéré que l'événement endosymbiotique à l'origine d'Archaeplastida s'est produit il y a 1 à 1,5 milliard d'années, au moins 5 cents millions d'années après que les fossiles suggèrent la présence d'eucaryotes.
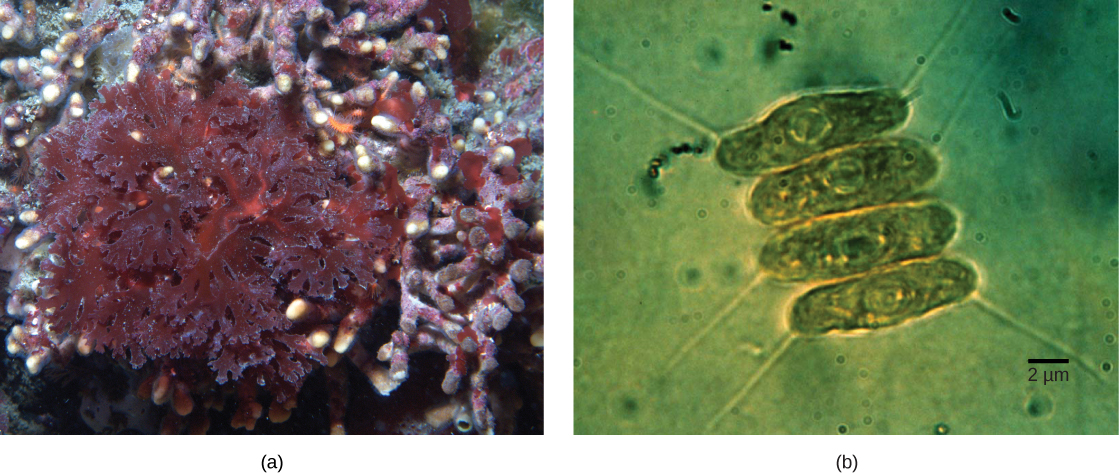
Les plastes des eucaryotes ne proviennent pas tous directement de l'endosymbiose primaire. Certains des principaux groupes d'algues sont devenus photosynthétiques par endosymbiose secondaire, c'est-à-dire en absorbant des algues vertes ou des algues rouges (toutes deux issues d'Archaeplastida) comme endosymbiotes (Figure\(\PageIndex{3}\)). De nombreuses études microscopiques et génétiques ont confirmé cette conclusion. Les plastes secondaires sont entourés de trois membranes ou plus, et certains plastes secondaires contiennent même des restes clairs du noyau d'une algue endosymbiotique. D'autres n'ont « conservé » aucun vestige. Dans certains cas, les événements endosymbiotiques d'ordre tertiaire ou supérieur sont la meilleure explication de la présence de plastes chez certains eucaryotes.
Art Connection
Quelles sont les preuves que les mitochondries ont été incorporées dans la cellule eucaryote ancestrale avant les chloroplastes ?
Lien évolutif : endosymbiose secondaire chez les chlorarachniophytes
L'endosymbiose implique qu'une cellule en engloutit une autre pour produire, au fil du temps, une relation co-évolutive dans laquelle aucune des cellules ne pourrait survivre seule. Les chloroplastes des algues rouges et vertes, par exemple, proviennent de l'engloutissement d'une cyanobactérie photosynthétique par un procaryote précoce.
Cela soulève la question de savoir s'il est possible qu'une cellule contenant un endosymbiote pour elle-même soit engloutie, entraînant une endosymbiose secondaire. Des preuves moléculaires et morphologiques suggèrent que les protistes chlorarachniophytes sont issus d'un événement endosymbiotique secondaire. Les chlorarachniophytes sont des algues rares indigènes des mers tropicales et du sable qui peuvent être classées dans le supergroupe des rhizaires. Les chlorarachniophytes étendent de minces brins cytoplasmiques, s'interconnectant avec d'autres chlorarachniophytes, dans un réseau cytoplasmique. On pense que ces protistes sont apparus lorsqu'un eucaryote a englouti une algue verte, cette dernière ayant déjà établi une relation endosymbiotique avec une cyanobactérie photosynthétique (Figure\(\PageIndex{5}\)).
Plusieurs sources de données soutiennent que les chlorarachniophytes ont évolué à partir d'une endosymbiose secondaire. Les chloroplastes contenus dans les endosymbiotes des algues vertes sont toujours capables de photosynthèse, rendant les chlorarachniophytes photosynthétiques. L'endosymbiote de l'algue verte présente également un noyau vestigial rabougri. En fait, il semble que les chlorarachniophytes soient les produits d'un événement endosymbiotique secondaire récent sur le plan de l'évolution. Les plastes des chlorarachniophytes sont entourés de quatre membranes : les deux premières correspondent aux membranes interne et externe de la cyanobactérie photosynthétique, la troisième correspond à l'algue verte et la quatrième correspond à la vacuole qui entourait l'algue verte lorsqu'elle a été engloutie par le ancêtre chlorarachniophyte. Dans d'autres lignées impliquant une endosymbiose secondaire, seules trois membranes peuvent être identifiées autour des plastes. Ceci est actuellement rectifié sous la forme d'une perte séquentielle d'une membrane au cours de l'évolution.
Le processus d'endosymbiose secondaire n'est pas propre aux chlorarachniophytes. En fait, l'endosymbiose secondaire des algues vertes a également entraîné la formation de protistes euglénidés, tandis que l'endosymbiose secondaire des algues rouges a entraîné l'évolution de dinoflagellés, d'apicomplexans et de stramenopiles.
Résumé
La plus ancienne preuve fossile d'eucaryotes date d'environ 2 milliards d'années. Les fossiles plus anciens semblent tous être des procaryotes. Il est probable que les eucaryotes actuels descendent d'un ancêtre qui avait une organisation procaryote. Le dernier ancêtre commun de l'Eucarya d'aujourd'hui présentait plusieurs caractéristiques, notamment des cellules dont les noyaux se divisaient par mitose et contenaient des chromosomes linéaires où l'ADN était associé à des histones, un système cytosquelettique et endomembranaire, et la capacité de fabriquer des cilies/flagelles pendant au moins une partie de sa vie cycle. Il était aérobie parce que ses mitochondries étaient le résultat d'une alpha-protéobactérie aérobie vivant à l'intérieur d'une cellule hôte. On ne sait toujours pas si cet hôte possédait un noyau au moment de la symbiose initiale. Le dernier ancêtre commun a peut-être eu une paroi cellulaire pendant au moins une partie de son cycle de vie, mais d'autres données sont nécessaires pour confirmer cette hypothèse. Les eucaryotes d'aujourd'hui sont très divers dans leurs formes, leur organisation, leurs cycles de vie et le nombre de cellules par individu.
Connexions artistiques
Figure\(\PageIndex{4}\) : Quelles sont les preuves que les mitochondries ont été incorporées dans la cellule eucaryote ancestrale avant les chloroplastes ?
- Réponse
-
Toutes les cellules eucaryotes possèdent des mitochondries, mais toutes les cellules eucaryotes ne possèdent pas de chloroplastes.
Lexique
- endosymbiose
- engloutissement d'une cellule dans une autre de telle sorte que la cellule engloutie survit et que les deux cellules en bénéficient ; le processus responsable de l'évolution des mitochondries et des chloroplastes chez les eucaryotes
- théorie endosymbiotique
- théorie selon laquelle les eucaryotes peuvent avoir été le produit d'une cellule qui en engloutit une autre, de l'une vivant dans une autre et d'une évolution au fil du temps jusqu'à ce que les cellules séparées ne soient plus reconnaissables en tant que telles
- plaste
- membre d'un groupe d'organites apparentés dans les cellules végétales qui participent au stockage des amidons, des graisses, des protéines et des pigments