
12.4 : Le potentiel d'action
- Page ID
- 195308

Objectifs d'apprentissage
- Décrire les composants de la membrane qui établissent le potentiel de la membrane au repos
- Décrire les modifications qui se produisent sur la membrane et qui se traduisent par le potentiel d'action
Les fonctions du système nerveux (sensation, intégration et réponse) dépendent des fonctions des neurones qui sous-tendent ces voies. Pour comprendre comment les neurones sont capables de communiquer, il est nécessaire de décrire le rôle d'une membrane excitable dans la génération de ces signaux. La base de cette communication est le potentiel d'action, qui montre comment les modifications de la membrane peuvent constituer un signal. L'examen de la façon dont ces signaux fonctionnent dans des circonstances plus variables implique d'examiner les potentiels gradués, qui seront abordés dans la section suivante.
Membranes cellulaires électriquement actives
La plupart des cellules du corps utilisent des particules chargées, des ions, pour accumuler une charge à travers la membrane cellulaire. Auparavant, il a été démontré que cela faisait partie du fonctionnement des cellules musculaires. La contraction des muscles squelettiques, basée sur le couplage excitation—contraction, nécessite l'intervention d'un neurone. Les deux cellules utilisent la membrane cellulaire pour réguler le mouvement des ions entre le liquide extracellulaire et le cytosol.
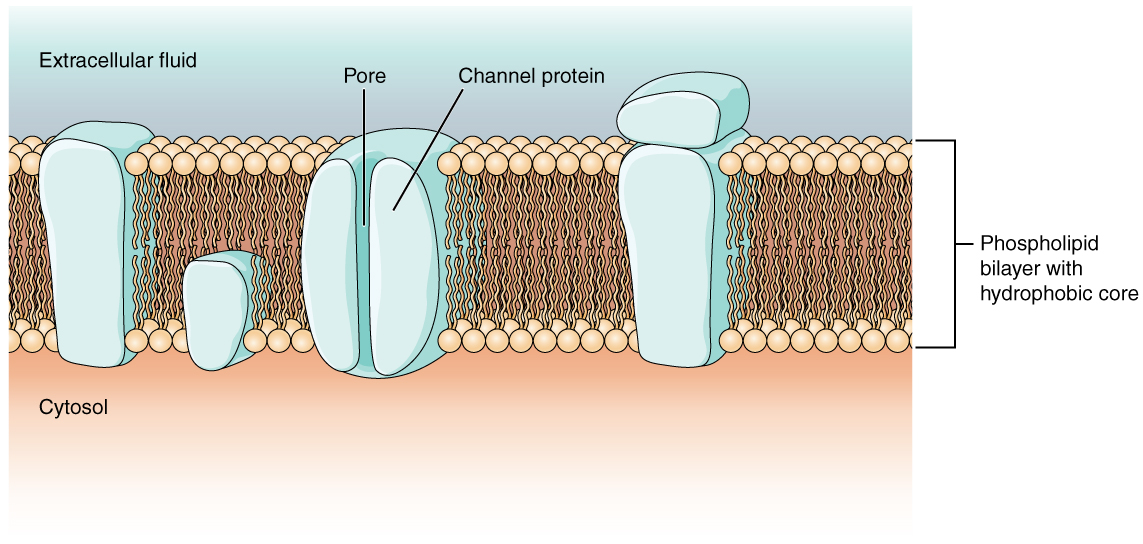
Comme vous l'avez appris dans le chapitre sur les cellules, la membrane cellulaire est principalement responsable de la régulation de ce qui peut traverser la membrane et de ce qui ne reste que d'un seul côté. La membrane cellulaire est une bicouche de phospholipides, de sorte que seules les substances qui peuvent passer directement à travers le noyau hydrophobe peuvent diffuser sans aide. Les particules chargées, qui sont hydrophiles par définition, ne peuvent pas traverser la membrane cellulaire sans assistance (Figure\(\PageIndex{1}\)). Les protéines transmembranaires, en particulier les protéines de canal, rendent cela possible. Plusieurs canaux de transport passifs, ainsi que des pompes de transport actives, sont nécessaires pour générer un potentiel transmembranaire et un potentiel d'action. La protéine porteuse appelée pompe sodium/potassium qui déplace les ions sodium (Na +) d'une cellule et les ions potassium (K +) vers une cellule, régulant ainsi la concentration d'ions des deux côtés de la membrane cellulaire, présente un intérêt particulier.
La pompe sodium/potassium nécessite de l'énergie sous forme d'adénosine triphosphate (ATP), elle est donc également appelée ATPase. Comme cela a été expliqué dans le chapitre sur les cellules, la concentration de Na + est plus élevée à l'extérieur de la cellule qu'à l'intérieur, et la concentration de K + est plus élevée à l'intérieur de la cellule qu'à l'extérieur. Cela signifie que cette pompe déplace les ions à contre-courant des gradients de concentration du sodium et du potassium, raison pour laquelle elle a besoin d'énergie. En fait, la pompe maintient essentiellement ces gradients de concentration.
Les canaux ioniques sont des pores qui permettent à des particules chargées spécifiques de traverser la membrane en réponse à un gradient de concentration existant. Les protéines sont capables de couvrir la membrane cellulaire, y compris son noyau hydrophobe, et peuvent interagir avec la charge des ions en raison des propriétés variées des acides aminés présents dans des domaines ou des régions spécifiques du canal protéique. Les acides aminés hydrophobes se trouvent dans les domaines apposés aux queues hydrocarbonées des phospholipides. Les acides aminés hydrophiles sont exposés aux environnements fluides du liquide extracellulaire et du cytosol. De plus, les ions interagiront avec les acides aminés hydrophiles, qui seront sélectifs en fonction de la charge de l'ion. Les canaux pour les cations (ions positifs) comporteront des chaînes latérales chargées négativement dans le pore. Les canaux pour les anions (ions négatifs) auront des chaînes latérales chargées positivement dans le pore. C'est ce que l'on appelle l'exclusion électrochimique, ce qui signifie que le pore du canal est spécifique à la charge.
Les canaux ioniques peuvent également être spécifiés par le diamètre du pore. La distance entre les acides aminés sera spécifique au diamètre de l'ion lorsqu'il se dissociera des molécules d'eau qui l'entourent. En raison des molécules d'eau environnantes, les pores plus grands ne sont pas idéaux pour les ions plus petits, car les molécules d'eau interagissent, par des liaisons hydrogène, plus facilement que les chaînes latérales des acides aminés. C'est ce qu'on appelle l'exclusion de taille Certains canaux ioniques sont sélectifs en termes de charge, mais pas nécessairement en termes de taille, et sont donc appelés canaux non spécifiques. Ces canaux non spécifiques permettent aux cations, en particulier au Na +, au K + et au Ca 2+, de traverser la membrane, mais excluent les anions.
Les canaux ioniques ne permettent pas toujours aux ions de se diffuser librement à travers la membrane. Certaines sont ouvertes par certains événements, ce qui signifie que les chaînes sont fermées. Une autre façon de catégoriser les chaînes consiste donc à se baser sur la façon dont elles sont fermées. Bien que ces classes de canaux ioniques se trouvent principalement dans les cellules du tissu nerveux ou musculaire, elles peuvent également être trouvées dans les cellules des tissus épithéliaux et conjonctifs.
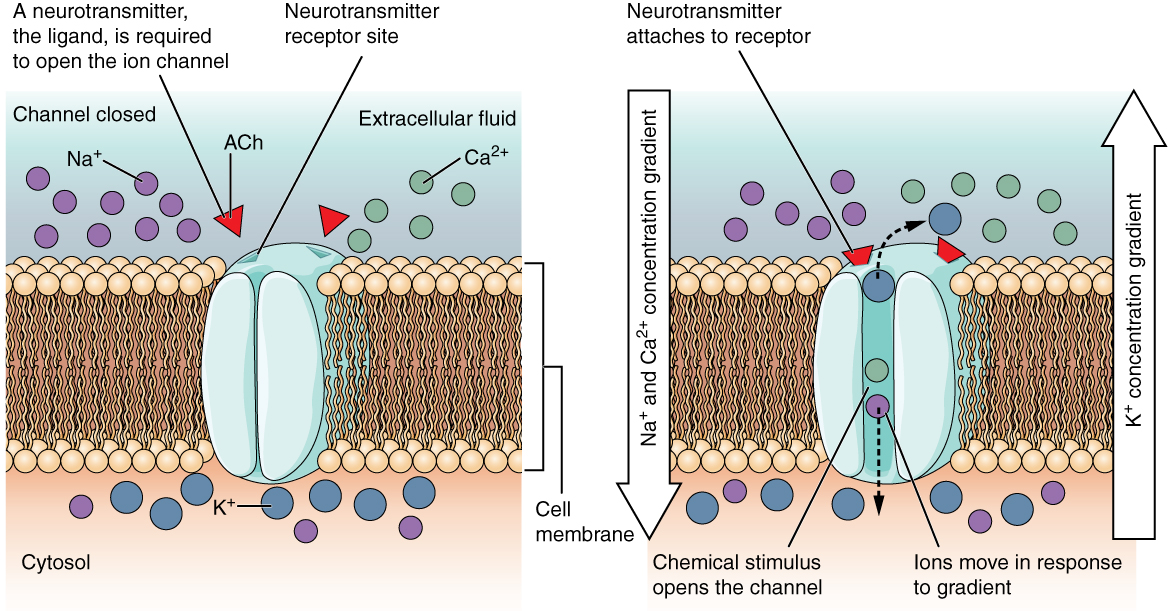
Un canal dépendant d'un ligand s'ouvre parce qu'une molécule de signalisation, un ligand, se lie à la région extracellulaire du canal. Ce type de canal est également connu sous le nom de récepteur ionotropique car lorsque le ligand, connu sous le nom de neurotransmetteur du système nerveux, se lie à la protéine, les ions traversent la membrane pour modifier sa charge (Figure\(\PageIndex{2}\)).
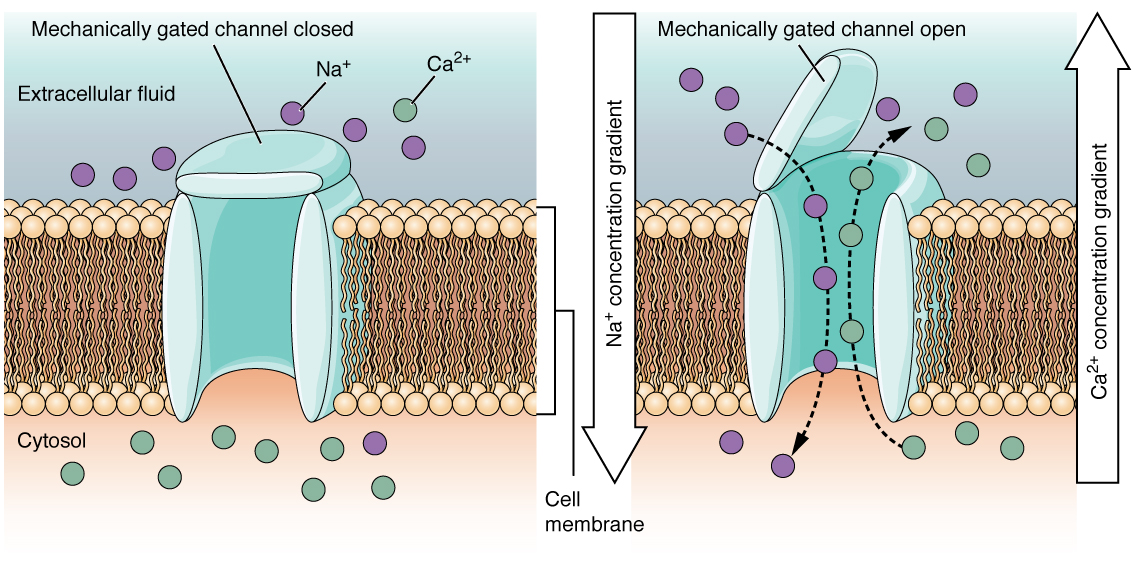
Un canal à commande mécanique s'ouvre en raison d'une distorsion physique de la membrane cellulaire. De nombreux canaux associés au sens du toucher (somatosensation) sont contrôlés mécaniquement. Par exemple, lorsque la pression est appliquée sur la peau, ces canaux s'ouvrent et permettent aux ions de pénétrer dans la cellule. Le canal qui s'ouvre en fonction des changements de température, comme lors du test de l'eau dans la douche, serait similaire à ce type de canal (Figure\(\PageIndex{3}\)).
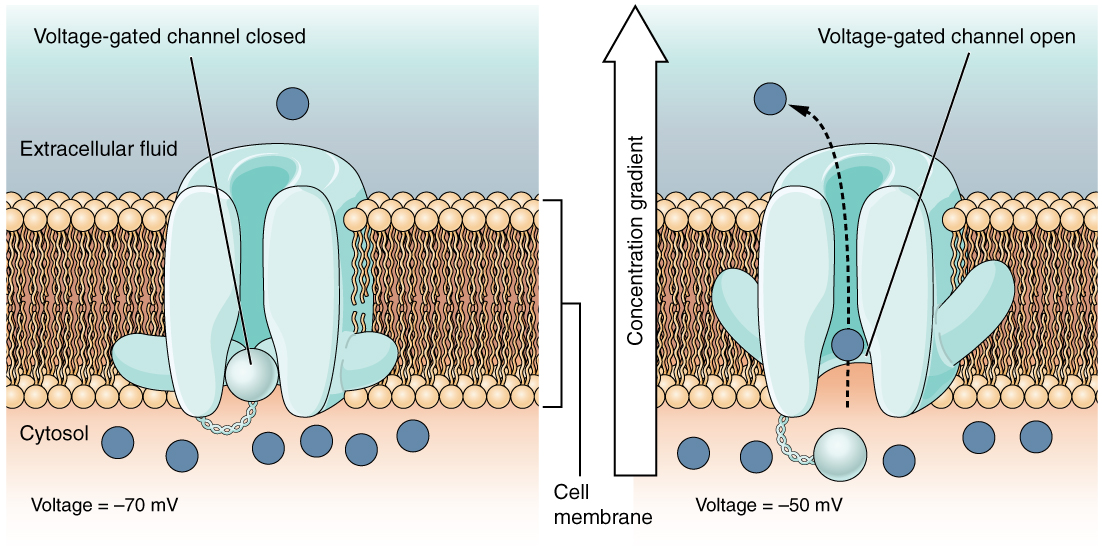
Un canal dépendant de la tension est un canal qui répond aux modifications des propriétés électriques de la membrane dans laquelle il est intégré. Normalement, la partie interne de la membrane est à une tension négative. Lorsque cette tension devient moins négative, le canal commence à permettre aux ions de traverser la membrane (Figure\(\PageIndex{4}\)).
Un canal de fuite est fermé de manière aléatoire, ce qui signifie qu'il s'ouvre et se ferme de manière aléatoire, d'où la référence à une fuite. Il n'y a aucun événement réel qui ouvre le canal ; au contraire, celui-ci a un taux intrinsèque de commutation entre les états ouvert et fermé. Les canaux de fuite contribuent à la tension transmembranaire de repos de la membrane excitable (Figure\(\PageIndex{5}\)).
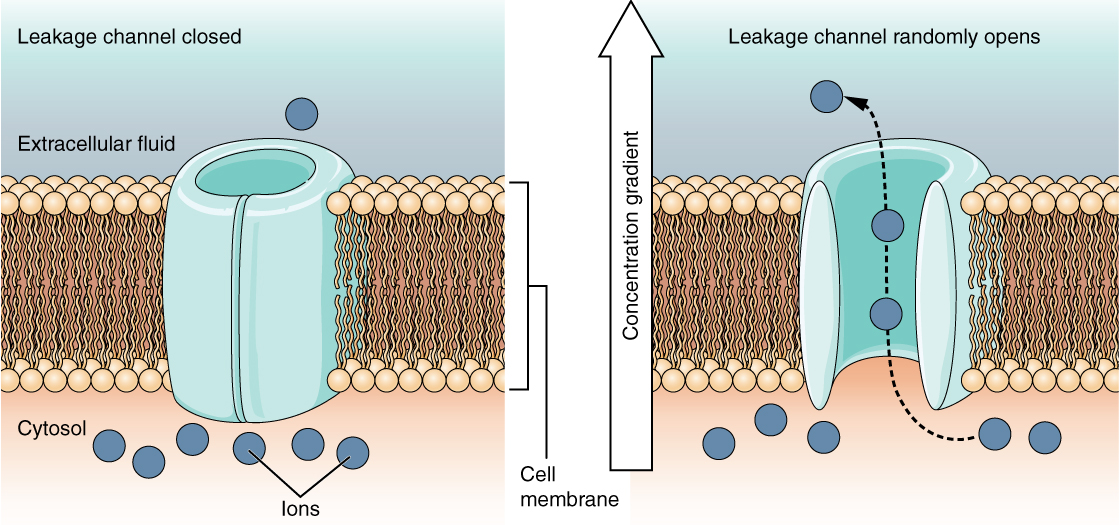
Le potentiel de la membrane
L'état électrique de la membrane cellulaire peut présenter plusieurs variations. Ce sont toutes des variations du potentiel de la membrane. Un potentiel est une distribution de charge à travers la membrane cellulaire, mesurée en millivolts (mV). La norme consiste à comparer l'intérieur de la cellule par rapport à l'extérieur, de sorte que le potentiel de la membrane est une valeur représentant la charge du côté intracellulaire de la membrane sur la base de la valeur nulle relative de l'extérieur (Figure\(\PageIndex{6}\)).
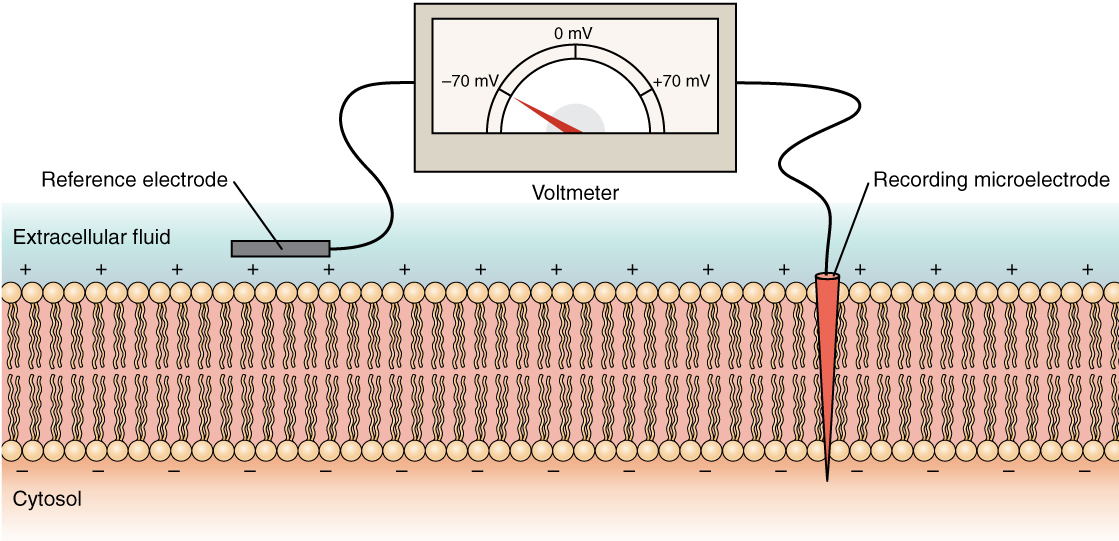
La concentration d'ions dans les fluides extracellulaires et intracellulaires est largement équilibrée, avec une charge neutre nette. Cependant, une légère différence de charge se produit directement à la surface de la membrane, à la fois à l'intérieur et à l'extérieur. C'est la différence dans cette région très limitée qui possède tout le pouvoir des neurones (et des cellules musculaires) pour générer des signaux électriques, y compris des potentiels d'action.
Avant de pouvoir décrire ces signaux électriques, il faut expliquer l'état de repos de la membrane. Lorsque la cellule est au repos et que les canaux ioniques sont fermés (à l'exception des canaux de fuite qui s'ouvrent de manière aléatoire), les ions sont répartis à travers la membrane de manière très prévisible. La concentration de Na + à l'extérieur de la cellule est 10 fois supérieure à la concentration à l'intérieur. De plus, la concentration de K + à l'intérieur de la cellule est supérieure à celle à l'extérieur. Le cytosol contient une forte concentration d'anions, sous forme d'ions phosphate et de protéines chargées négativement. Les gros anions sont un composant de la membrane cellulaire interne, y compris les phospholipides spécialisés et les protéines associées au feuillet interne de la membrane (feuillet est un terme utilisé pour désigner un côté de la membrane bicouche lipidique). La charge négative est localisée dans les gros anions.
Les ions étant répartis à travers la membrane à ces concentrations, la différence de charge est mesurée à -70 mV, la valeur décrite comme le potentiel de la membrane au repos. La valeur exacte mesurée pour le potentiel de la membrane au repos varie d'une cellule à l'autre, mais -70 mV est le plus souvent utilisée comme valeur. Cette tension serait en fait beaucoup plus faible, à l'exception des contributions de certaines protéines importantes dans la membrane. Les canaux de fuite permettent au Na + de pénétrer lentement dans la cellule ou au K + de sortir lentement, et la pompe Na + /K + les rétablit. Cela peut sembler être un gaspillage d'énergie, mais chacun joue un rôle dans le maintien du potentiel de la membrane.
Le potentiel d'action
Le potentiel de membrane au repos décrit l'état stable de la cellule, qui est un processus dynamique équilibré par une fuite d'ions et un pompage d'ions. Sans aucune influence extérieure, cela ne changera pas. Pour déclencher un signal électrique, le potentiel de la membrane doit changer.
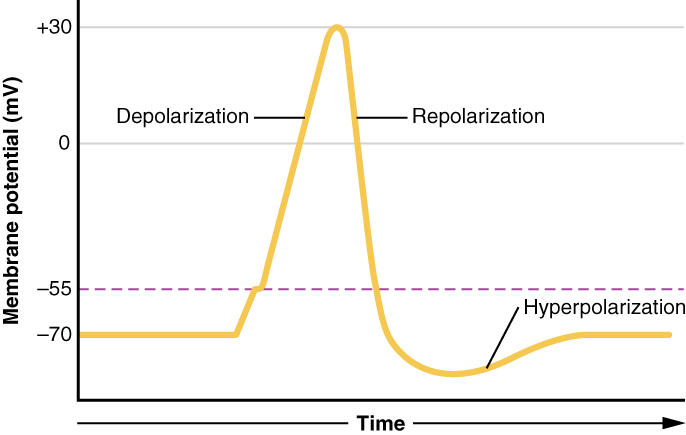
Cela commence par l'ouverture d'un canal pour le Na + dans la membrane. Comme la concentration de Na + est 10 fois plus élevée à l'extérieur de la cellule qu'à l'intérieur de la cellule, des ions se précipitent dans la cellule, principalement sous l'effet du gradient de concentration. Comme le sodium est un ion chargé positivement, il modifiera la tension relative immédiatement à l'intérieur de la cellule par rapport à l'extérieur. Le potentiel de repos est l'état de la membrane à une tension de -70 mV, de sorte que le cation sodium entrant dans la cellule la rendra moins négative. C'est ce qu'on appelle la dépolarisation, ce qui signifie que le potentiel de la membrane se déplace vers zéro
Le gradient de concentration du Na + est si fort qu'il continuera à pénétrer dans la cellule même après que le potentiel de la membrane soit devenu nul, de sorte que la tension immédiatement autour du pore commence à devenir positive. Le gradient électrique joue également un rôle, car les protéines négatives situées sous la membrane attirent l'ion sodium. Le potentiel de la membrane atteindra +30 mV lorsque le sodium aura pénétré dans la cellule.
Lorsque le potentiel de la membrane atteint +30 mV, d'autres canaux dépendants de la tension s'ouvrent dans la membrane. Ces canaux sont spécifiques à l'ion potassium. Un gradient de concentration agit également sur K +. Lorsque K + commence à quitter la cellule, emportant avec elle une charge positive, le potentiel de la membrane commence à revenir à sa tension de repos. C'est ce que l'on appelle la repolarisation, ce qui signifie que la tension de la membrane revient vers la valeur de -70 mV du potentiel de la membrane au repos.
La repolarisation ramène le potentiel de la membrane à la valeur de -70 mV qui indique le potentiel de repos, mais elle dépasse en fait cette valeur. Les ions potassium atteignent l'équilibre lorsque la tension de la membrane est inférieure à -70 mV, de sorte qu'une période d'hyperpolarisation se produit lorsque les canaux K + sont ouverts. La fermeture de ces chaînes K + est légèrement retardée, ce qui explique ce court dépassement.
Ce qui a été décrit ici est le potentiel d'action, qui est présenté sous forme de graphique de la tension dans le temps sur la Figure\(\PageIndex{6}\). C'est le signal électrique que le tissu nerveux génère pour communiquer. La variation de la tension de la membrane de -70 mV au repos à +30 mV à la fin de la dépolarisation est une variation de 100 mV. Cela peut également être écrit sous la forme d'une modification de 0,1 V. Pour mettre cette valeur en perspective, pensez à une batterie. Une pile AA que vous pourriez trouver dans une télécommande de téléviseur a une tension de 1,5 V, ou une pile de 9 V (la batterie rectangulaire avec deux bornes à une extrémité) est évidemment de 9 V. La variation observée dans le potentiel d'action est inférieure d'un ou deux ordres de grandeur à la charge de ces batteries. En fait, le potentiel de la membrane peut être décrit comme une batterie. Une charge est stockée à travers la membrane et peut être libérée dans les bonnes conditions. Une batterie de votre télécommande a stocké une charge qui est « libérée » lorsque vous appuyez sur un bouton.
Exercice\(\PageIndex{1}\)
Ce qui se passe à travers la membrane d'une cellule électriquement active est un processus dynamique difficile à visualiser à l'aide d'images statiques ou de descriptions textuelles. Visionnez cette animation pour en savoir plus sur ce processus. Quelle est la différence entre la force motrice du Na+ et celle du K+ ? Et qu'y a-t-il de similaire dans le mouvement de ces deux ions ?
La question qui se pose maintenant est la suivante : qu'est-ce qui déclenche le potentiel d'action ? La description ci-dessus passe commodément ce point sous silence. Mais c'est essentiel pour comprendre ce qui se passe. Le potentiel de la membrane restera à la tension de repos jusqu'à ce que quelque chose change. La description ci-dessus indique simplement qu'un canal Na + s'ouvre. Or, dire « un canal s'ouvre » ne signifie pas qu'une protéine transmembranaire individuelle change. Au lieu de cela, cela signifie qu'un type de canal s'ouvre. Il existe différents types de canaux qui permettent au Na + de traverser la membrane. Un canal Na+ dépendant d'un ligand s'ouvre lorsqu'un neurotransmetteur s'y lie et un canal Na+ mécaniquement activé s'ouvre lorsqu'un stimulus physique affecte un récepteur sensoriel (comme une pression appliquée sur la peau comprime un récepteur tactile). Qu'il s'agisse d'un neurotransmetteur se liant à sa protéine réceptrice ou d'un stimulus sensoriel activant une cellule réceptrice sensorielle, certains stimuli déclenchent le processus. Le sodium commence à pénétrer dans la cellule et la membrane devient moins négative.
Un troisième type de canal qui joue un rôle important dans la dépolarisation du potentiel d'action est le canal Na + dépendant de la tension. Les canaux qui commencent à dépolariser la membrane à cause d'un stimulus aident la cellule à se dépolariser de -70 mV à -55 mV. Une fois que la membrane atteint cette tension, les canaux Na + dépendants de la tension s'ouvrent. C'est ce que l'on appelle le seuil. Toute dépolarisation qui ne modifie pas le potentiel de la membrane à -55 mV ou plus n'atteindra pas le seuil et n'entraînera donc pas de potentiel d'action. De plus, tout stimulus qui dépolarise la membrane à -55 mV ou au-delà provoquera l'ouverture d'un grand nombre de canaux et un potentiel d'action sera initié.
En raison du seuil, le potentiel d'action peut être assimilé à un événement numérique, qu'il se produise ou non. Si le seuil n'est pas atteint, aucun potentiel d'action ne se produit. Si la dépolarisation atteint -55 mV, le potentiel d'action se poursuit et s'étend jusqu'à +30 mV, date à laquelle K + provoque une repolarisation, y compris le dépassement hyperpolarisant. De plus, ces modifications sont les mêmes pour chaque potentiel d'action, ce qui signifie qu'une fois le seuil atteint, la même chose se produit. Un stimulus plus fort, qui pourrait dépolariser la membrane bien au-delà du seuil, n'augmentera pas le potentiel d'action. Les potentiels d'action sont « tout ou rien ». Soit la membrane atteint le seuil et tout se passe comme décrit ci-dessus, soit la membrane n'atteint pas le seuil et rien d'autre ne se passe. Tous les potentiels d'action culminent à la même tension (+30 mV), de sorte qu'un potentiel d'action n'est pas plus grand qu'un autre. Des stimuli plus puissants déclencheront de multiples potentiels d'action plus rapidement, mais les signaux individuels ne sont pas plus importants. Ainsi, par exemple, vous ne ressentirez pas une plus grande sensation de douleur ou une contraction musculaire plus forte, en raison de l'ampleur du potentiel d'action, car ils ne sont pas de tailles différentes.
Comme nous l'avons vu, la dépolarisation et la repolarisation d'un potentiel d'action dépendent de deux types de canaux (le canal Na + dépendant de la tension et le canal K + dépendant de la tension). Le canal Na + dépendant de la tension possède en fait deux portes. L'une est la porte d'activation, qui s'ouvre lorsque le potentiel de la membrane passe à -55 mV. L'autre porte est la porte d'inactivation, qui se ferme après un certain laps de temps, de l'ordre d'une fraction de milliseconde. Lorsqu'une cellule est au repos, la porte d'activation est fermée et la porte d'inactivation est ouverte. Cependant, lorsque le seuil est atteint, la porte d'activation s'ouvre, permettant au Na + de se précipiter dans la cellule. Chronométrée avec le pic de dépolarisation, la porte d'inactivation se ferme. Lors de la repolarisation, plus aucun sodium ne peut pénétrer dans la cellule. Lorsque le potentiel de la membrane passe à nouveau à -55 mV, la porte d'activation se ferme. Ensuite, la porte d'inactivation s'ouvre à nouveau, rendant le canal prêt à recommencer l'ensemble du processus.
Le canal K + dépendant de la tension ne possède qu'une seule grille, qui est sensible à une tension de membrane de -50 mV. Cependant, il ne s'ouvre pas aussi rapidement que le canal Na + dépendant de la tension. Cela peut prendre une fraction de milliseconde pour que le canal s'ouvre une fois que cette tension est atteinte. Cela coïncide exactement avec le moment où le débit de Na + atteint son pic, de sorte que les canaux K + dépendants de la tension s'ouvrent juste au moment où les canaux Na + dépendants de la tension sont inactivés. Lorsque le potentiel de la membrane se repolarise et que la tension repasse à -50 mV, le canal se ferme, à nouveau, avec un léger retard. Le potassium continue de quitter la cellule pendant un court moment et le potentiel de la membrane devient plus négatif, ce qui entraîne un dépassement hyperpolarisant. Ensuite, le canal se referme et la membrane peut revenir à son potentiel de repos en raison de l'activité continue des canaux non fermés et de la pompe Na + /K +.
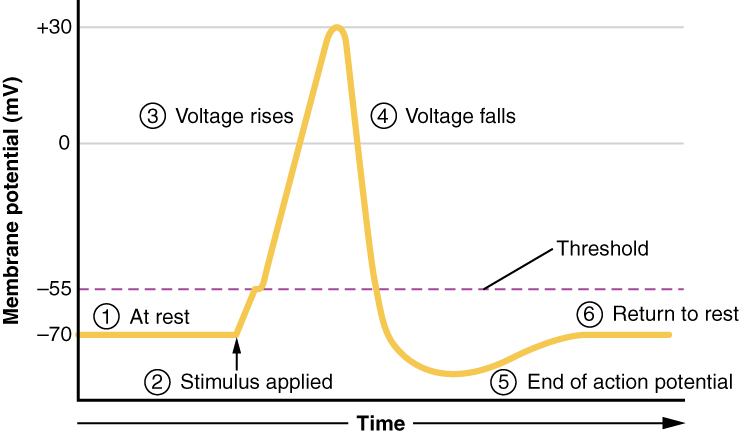
Tout cela se produit en 2 millisecondes environ (Figure\(\PageIndex{7}\)). Pendant qu'un potentiel d'action est en cours, il n'est pas possible d'en initier un autre. Cet effet est appelé période réfractaire. La période réfractaire comporte deux phases : la période réfractaire absolue et la période réfractaire relative. Pendant la phase absolue, aucun autre potentiel d'action ne démarrera. Cela est dû à la porte d'inactivation du canal Na + dépendant de la tension. Une fois que ce canal est revenu à sa conformation de repos (moins de -55 mV), un nouveau potentiel d'action peut être déclenché, mais uniquement par un stimulus plus puissant que celui qui a initié le potentiel d'action actuel. Cela est dû au flux de K + hors de la cellule. Comme cet ion sort précipitamment, tout Na+ qui tente d'entrer ne dépolarisera pas la cellule, mais empêchera simplement la cellule de s'hyperpolariser.
Propagation du potentiel d'action
Le potentiel d'action est initié au début de l'axone, au niveau de ce que l'on appelle le segment initial. Il existe une densité élevée de canaux Na + dépendants de la tension, ce qui permet une dépolarisation rapide. Le potentiel d'action se propage le long de l'axone, car davantage de canaux Na + dépendants de la tension s'ouvrent à mesure que la dépolarisation se propage. Cette propagation se produit parce que le Na + entre par le canal et se déplace le long de l'intérieur de la membrane cellulaire. Lorsque le Na + se déplace ou circule sur une courte distance le long de la membrane cellulaire, sa charge positive dépolarise un peu plus la membrane cellulaire. Au fur et à mesure que cette dépolarisation se propage, de nouveaux canaux Na + dépendants de la tension s'ouvrent et de plus en plus d'ions pénètrent dans la cellule, propageant la dépolarisation un peu plus loin.
Comme les canaux Na + dépendants de la tension sont inactivés au pic de la dépolarisation, ils ne peuvent pas être ouverts à nouveau pendant une courte période, soit la période réfractaire absolue. De ce fait, la propagation de la dépolarisation vers les canaux précédemment ouverts n'a aucun effet. Le potentiel d'action doit se propager vers les terminaisons axonales ; par conséquent, la polarité du neurone est maintenue, comme mentionné ci-dessus.
La propagation, telle que décrite ci-dessus, s'applique aux axones non myélinisés. Lorsque la myélinisation est présente, le potentiel d'action se propage différemment. Les ions sodium qui pénètrent dans la cellule par le segment initial commencent à se propager sur la longueur du segment axonal, mais il n'y a pas de canaux Na + dépendants de la tension avant le premier nœud de Ranvier. Comme il n'y a pas d'ouverture constante de ces canaux le long du segment axonal, la dépolarisation se propage à une vitesse optimale. La distance entre les nœuds est la distance optimale pour maintenir la membrane toujours dépolarisée au-dessus du seuil au nœud suivant. Lorsque le Na + se répand à l'intérieur de la membrane du segment axonal, la charge commence à se dissiper. Si le nœud était plus bas dans l'axone, cette dépolarisation aurait été trop faible pour que les canaux Na + dépendants de la tension puissent être activés au nœud suivant de Ranvier. Si les nœuds étaient plus proches les uns des autres, la vitesse de propagation serait plus lente.
La propagation le long d'un axone non myélinisé est appelée conduction continue ; le long d'un axone myélinisé, il s'agit d'une conduction saltatoire. La conduction continue est lente car des canaux Na + dépendants de la tension s'ouvrent toujours et de plus en plus de Na + se précipitent dans la cellule. La conduction saltatoire est plus rapide car le potentiel d'action saute essentiellement d'un nœud à l'autre (saltare = « sauter »), et le nouvel afflux de Na + renouvelle la membrane dépolarisée. Parallèlement à la myélinisation de l'axone, le diamètre de l'axone peut influencer la vitesse de conduction. Tout comme l'eau coule plus vite dans une large rivière que dans un ruisseau étroit, la dépolarisation basée sur le Na + se propage plus rapidement le long d'un large axone que le long d'un étroit axone. Ce concept est connu sous le nom de résistance et s'applique généralement aux fils électriques ou à la plomberie, tout comme il est vrai pour les axones, bien que les conditions spécifiques soient différentes à l'échelle des électrons ou des ions par rapport à l'eau d'une rivière.
DÉSÉQUILIBRES HOMÉOSTATIQUES : concentration
Les cellules gliales, en particulier les astrocytes, sont responsables du maintien de l'environnement chimique du tissu du SNC. Les concentrations d'ions dans le liquide extracellulaire sont à la base de la façon dont le potentiel de la membrane est établi et des modifications de la signalisation électrochimique. Si l'équilibre des ions est perturbé, des résultats drastiques sont possibles.
Normalement, la concentration de K+ est plus élevée à l'intérieur du neurone qu'à l'extérieur. Après la phase de repolarisation du potentiel d'action, les canaux de fuite de K+ et la pompe Na+/K+ garantissent le retour des ions à leur emplacement d'origine. À la suite d'un accident vasculaire cérébral ou d'un autre événement ischémique, les taux de K+ extracellulaire sont élevés. Les astrocytes de la région sont équipés pour éliminer l'excès de K+ afin de faciliter la pompe. Mais lorsque le niveau est très déséquilibré, les effets peuvent être irréversibles.
Les astrocytes peuvent devenir réactifs dans de tels cas, ce qui nuit à leur capacité à maintenir l'environnement chimique local. Les cellules gliales s'agrandissent et leurs processus se gonflent. Ils perdent leur capacité d'amortissement du K+ et le fonctionnement de la pompe est affecté, voire inversé. L'un des premiers signes d'une maladie cellulaire est cette « fuite » d'ions sodium dans les cellules du corps. Ce déséquilibre sodium/potassium affecte négativement la chimie interne des cellules, les empêchant de fonctionner normalement.
Visitez ce site pour découvrir un laboratoire virtuel de neurophysiologie et pour observer les processus électrophysiologiques du système nerveux, où les scientifiques mesurent directement les signaux électriques produits par les neurones. Souvent, les potentiels d'action apparaissent si rapidement qu'il n'est pas utile de regarder un écran pour les voir apparaître. Un haut-parleur est alimenté par les signaux enregistrés par un neurone et il « surgit » chaque fois que le neurone déclenche un potentiel d'action. Ces potentiels d'action se déclenchent si vite que cela semble statique à la radio. Les électrophysiologistes peuvent reconnaître les modèles à l'intérieur de cette statique pour comprendre ce qui se passe. Pourquoi le modèle de sangsue est-il utilisé pour mesurer l'activité électrique des neurones au lieu d'utiliser des humains ?
Révision
Le système nerveux est caractérisé par des signaux électriques envoyés d'une zone à l'autre. Que ces zones soient proches ou très éloignées les unes des autres, le signal doit se déplacer le long d'un axone. La base du signal électrique est la distribution contrôlée des ions à travers la membrane. Les canaux ioniques transmembranaires régulent le moment où les ions peuvent entrer ou sortir de la cellule, de sorte qu'un signal précis est généré. Ce signal est le potentiel d'action qui a une forme très caractéristique basée sur les variations de tension à travers la membrane au cours d'une période donnée.
La membrane est normalement au repos avec des concentrations établies de Na + et de K + de chaque côté. Un stimulus déclenchera la dépolarisation de la membrane, et les canaux dépendants de la tension entraîneront une dépolarisation supplémentaire suivie d'une repolarisation de la membrane. Un léger dépassement de l'hyperpolarisation marque la fin du potentiel d'action. Lorsqu'un potentiel d'action est en cours, un autre ne peut pas être généré dans les mêmes conditions. Lorsque le canal Na + dépendant de la tension est inactivé, aucun potentiel d'action ne peut être généré. Une fois que ce canal est revenu à son état de repos, un nouveau potentiel d'action est possible, mais il doit être déclenché par un stimulus relativement plus fort pour vaincre le K + quittant la cellule.
Le potentiel d'action se déplace le long de l'axone lorsque des canaux ioniques dépendants de la tension sont ouverts par la dépolarisation étalée. Dans les axones non myélinisés, cela se produit de manière continue car il existe des canaux dépendants de la tension dans toute la membrane. Dans les axones myélinisés, la propagation est qualifiée de saltatoire parce que les canaux dépendants de la tension ne se trouvent qu'aux nœuds de Ranvier et que les événements électriques semblent « sauter » d'un nœud à l'autre. La conduction saltatoire est plus rapide que la conduction continue, ce qui signifie que les axones myélinisés propagent leurs signaux plus rapidement. Le diamètre de l'axone fait également la différence, car les ions qui diffusent à l'intérieur de la cellule ont moins de résistance dans un espace plus large.
Questions sur les liens interactifs
Ce qui se passe à travers la membrane d'une cellule électriquement active est un processus dynamique difficile à visualiser à l'aide d'images statiques ou de descriptions textuelles. Regardez cette animation pour bien comprendre le processus. Quelle est la différence entre la force motrice du Na + et celle du K + ? Et qu'y a-t-il de similaire dans le mouvement de ces deux ions ?
Réponse : Le sodium pénètre dans la cellule en raison de l'immense gradient de concentration, alors que le potassium s'en éloigne à cause de la dépolarisation provoquée par le sodium. Cependant, ils descendent tous les deux leurs gradients respectifs, vers l'équilibre.
Visitez ce site pour découvrir un laboratoire virtuel de neurophysiologie et pour observer les processus électrophysiologiques du système nerveux, où les scientifiques mesurent directement les signaux électriques produits par les neurones. Souvent, les potentiels d'action apparaissent si rapidement qu'il n'est pas utile de regarder un écran pour les voir apparaître. Un haut-parleur est alimenté par les signaux enregistrés par un neurone et il « surgit » chaque fois que le neurone déclenche un potentiel d'action. Ces potentiels d'action se déclenchent si vite que cela semble statique à la radio. Les électrophysiologistes peuvent reconnaître les modèles à l'intérieur de cette statique pour comprendre ce qui se passe. Pourquoi le modèle de sangsue est-il utilisé pour mesurer l'activité électrique des neurones au lieu d'utiliser des humains ?
Réponse : Les propriétés de l'électrophysiologie étant communes à tous les animaux, l'utilisation de la sangsue est une approche plus simple et plus humaine pour étudier les propriétés de ces cellules. Il existe des différences entre le système nerveux des invertébrés (comme une sangsue) et celui des vertébrés, mais pas pour les fins de l'étude de ces expériences.
Questions de révision
Q. Quel ion entre dans un neurone et provoque la dépolarisation de la membrane cellulaire ?
A. sodium
B. chlorure
C. potassium
D. phosphate
Réponse : A
Q. Les canaux Na + dépendants de la tension s'ouvrent lorsqu'ils atteignent quel état ?
A. potentiel de repos
B. seuil
C. repolarisation
D. dépassement
Réponse : B
Q. De quoi a besoin un canal dépendant d'un ligand pour s'ouvrir ?
A. augmentation de la concentration en ions Na +
B. liaison d'un neurotransmetteur
C. augmentation de la concentration en ions K +
D. dépolarisation de la membrane
Réponse : B
Q. À quoi répond un canal à verrouillage mécanique ?
A. stimulus physique
B. stimulus chimique
C. augmentation de la résistance
D. diminution de la résistance
Réponse : A
Q. Laquelle des tensions suivantes serait la plus susceptible d'être mesurée pendant la période réfractaire relative ?
A. +30 mV
B. 0 mV
C. -45 mV
D. -80 mg
Réponse : D
Q. Lequel des éléments suivants va probablement propager un potentiel d'action le plus rapidement ?
A. un axone fin et non myélinisé
B. un axone fin et myélinisé
C. un axone épais non myélinisé
D. un axone myélinisé épais
Réponse : D
Questions sur la pensée critique
Q. Que signifie le fait qu'une action potentielle soit un événement « tout ou rien » ?
R. La membrane cellulaire doit atteindre le seuil avant que les canaux Na+ dépendants de la tension ne s'ouvrent. Si le seuil n'est pas atteint, si ces canaux ne s'ouvrent pas et si la phase dépolarisante du potentiel d'action ne se produit pas, la membrane cellulaire reviendra simplement à son état de repos.
Q. La perception consciente de la douleur est souvent retardée en raison du temps qu'il faut aux sensations pour atteindre le cortex cérébral. Pourquoi cela serait-il le cas sur la base de la propagation du potentiel axonal ?
R. Les axones des neurones sensoriels sensibles à la douleur sont fins et non myélinisés, de sorte que cette sensation met plus de temps à atteindre le cerveau que les autres sensations.
Lexique
- période réfractaire absolue
- temps pendant une période d'action où un autre potentiel d'action ne peut pas être généré parce que le canal Na + dépendant de la tension est inactivé
- porte d'activation
- partie du canal Na + dépendant de la tension qui s'ouvre lorsque la tension de la membrane atteint le seuil
- conduction continue
- propagation lente d'un potentiel d'action le long d'un axone non myélinisé en raison de canaux Na + dépendants de la tension situés sur toute la longueur de la membrane cellulaire
- dépolarisation
- variation du potentiel de la membrane cellulaire entre le repos et zéro
- exclusion électrochimique
- principe de passage sélectif des ions dans un canal en fonction de leur charge
- membrane excitable
- membrane cellulaire qui régule le mouvement des ions afin de générer un signal électrique
- fermé
- propriété d'un canal qui détermine son mode d'ouverture dans des conditions spécifiques, telles qu'un changement de tension ou une déformation physique
- porte d'inactivation
- partie d'un canal Na + dépendant de la tension qui se ferme lorsque le potentiel de la membrane atteint +30 mV
- récepteur ionotropique
- un récepteur de neurotransmetteur qui agit comme une porte de canal ionique et qui s'ouvre par la liaison du neurotransmetteur
- canal de fuite
- canal ionique qui s'ouvre de manière aléatoire et qui n'est pas lié à un événement spécifique, également appelé canal non fermé
- canaux dépendants d'un ligand
- un autre nom pour un récepteur ionotropique dont un neurotransmetteur est le ligand
- canal fermé mécaniquement
- canal ionique qui s'ouvre lorsqu'un événement physique affecte directement la structure de la protéine
- potentiel membranaire
- distribution de la charge à travers la membrane cellulaire, sur la base des charges d'ions
- canal non spécifique
- canal qui n'est pas spécifique à un ion par rapport à un autre, tel qu'un canal cationique non spécifique qui permet à tout ion chargé positivement de traverser la membrane
- période réfractaire
- temps après l'initiation d'un potentiel d'action lorsqu'un autre potentiel d'action ne peut pas être généré
- période réfractaire relative
- période pendant la période réfractaire où un nouveau potentiel d'action ne peut être initié que par un stimulus plus fort que le potentiel d'action actuel, car les canaux K + dépendants de la tension ne sont pas fermés
- repolarisation
- retour du potentiel de la membrane à sa tension normalement négative à la fin du potentiel d'action
- résistance
- propriété d'un axone liée à la capacité des particules à diffuser à travers le cytoplasme ; cette propriété est inversement proportionnelle au diamètre de la fibre
- potentiel de membrane au repos
- la différence de tension mesurée à travers la membrane d'une cellule dans des conditions stables, généralement -70 mV
- conduction saltatoire
- propagation rapide du potentiel d'action le long d'un axone myélinisé en raison de la présence de canaux Na + dépendants de la tension uniquement aux nœuds de Ranvier
- exclusion de taille
- principe de passage sélectif des ions dans un canal en fonction de leur taille relative
- canal dépendant de la tension
- canal ionique qui s'ouvre en raison d'une modification de la charge distribuée à travers la membrane où il se trouve