
12.5 : Communication entre les neurones
- Page ID
- 195312

Objectifs d'apprentissage
- Expliquer les différences entre les types de potentiels gradués
- Classer les principaux neurotransmetteurs par type chimique et par effet
Les changements électriques qui se produisent dans un neurone, tels que décrits dans la section précédente, sont similaires à ceux d'un interrupteur allumé. Un stimulus déclenche la dépolarisation, mais le potentiel d'action s'exécute de lui-même une fois qu'un seuil est atteint. La question qui se pose maintenant est la suivante : « Qu'est-ce qui fait tourner l'interrupteur ? » Les variations temporaires de la tension de la membrane cellulaire peuvent résulter de la réception par les neurones d'informations provenant de l'environnement ou de l'action d'un neurone sur un autre. Ces types particuliers de potentiels influencent un neurone et déterminent si un potentiel d'action se produira ou non. Bon nombre de ces signaux transitoires proviennent de la synapse.
Potentiels gradués
Les variations locales du potentiel membranaire sont appelées potentiels gradués et sont généralement associées aux dendrites d'un neurone. L'ampleur de la modification du potentiel de la membrane est déterminée par la taille du stimulus qui la provoque. Dans l'exemple du test de la température de la douche, de l'eau légèrement chaude ne provoquerait qu'une légère modification d'un thermorécepteur, tandis que l'eau chaude provoquerait une modification importante du potentiel de la membrane.
Les potentiels gradués peuvent être de deux sortes, soit ils sont dépolarisants, soit hyperpolarisants (Figure\(\PageIndex{1}\)). Pour une membrane au potentiel de repos, un potentiel gradué représente une variation de cette tension supérieure à -70 mV ou inférieure à -70 mV. Les potentiels graduels dépolarisants sont souvent le résultat de l'entrée de Na + ou de Ca 2+ dans la cellule. Ces deux ions ont des concentrations plus élevées à l'extérieur de la cellule qu'à l'intérieur ; comme ils ont une charge positive, ils se déplacent dans la cellule, ce qui la rend moins négative par rapport à l'extérieur. Des potentiels gradués hyperpolarisants peuvent être provoqués par le fait que K + quitte la cellule ou que du Cl pénètre dans la cellule. Si une charge positive sort d'une cellule, la cellule devient plus négative ; si une charge négative entre dans la cellule, la même chose se produit.

Figure\(\PageIndex{1}\) : Potentiels gradués. Les potentiels gradués sont des variations temporaires de la tension de la membrane, dont les caractéristiques dépendent de la taille du stimulus. Certains types de stimuli provoquent une dépolarisation de la membrane, tandis que d'autres provoquent une hyperpolarisation. Cela dépend des canaux ioniques spécifiques qui sont activés dans la membrane cellulaire.
Types de potentiels gradués
Pour les cellules unipolaires des neurones sensoriels, qu'elles soient dotées de terminaisons nerveuses libres ou encapsulées, des potentiels gradués se développent dans les dendrites qui influencent la génération d'un potentiel d'action dans l'axone de la même cellule. C'est ce que l'on appelle un potentiel générateur. Pour d'autres cellules réceptrices sensorielles, telles que les cellules gustatives ou les photorécepteurs de la rétine, des potentiels gradués dans leurs membranes entraînent la libération de neurotransmetteurs au niveau des synapses des neurones sensoriels. C'est ce que l'on appelle le potentiel récepteur.
Un potentiel post-synaptique (PSP) est le potentiel graduel des dendrites d'un neurone qui reçoit des synapses provenant d'autres cellules. Les potentiels postsynaptiques peuvent être dépolarisants ou hyperpolarisants. La dépolarisation dans un potentiel postsynaptique est appelée potentiel post-synaptique excitateur (EPSP) car elle amène le potentiel membranaire à se rapprocher du seuil. L'hyperpolarisation dans un potentiel postsynaptique est un potentiel postsynaptique inhibiteur (IPSP) car elle provoque l'éloignement du potentiel membranaire par rapport au seuil.
Sommation
Tous les types de potentiels gradués entraîneront de petites variations de dépolarisation ou d'hyperpolarisation de la tension d'une membrane. Ces changements peuvent amener le neurone à atteindre le seuil si les changements s'additionnent ou s'additionnent. Les effets combinés de différents types de potentiels gradués sont illustrés dans la figure\(\PageIndex{2}\). Si la variation totale de tension dans la membrane est positive de 15 mV, ce qui signifie que la membrane se dépolarise de -70 mV à -55 mV, les potentiels gradués feront en sorte que la membrane atteigne le seuil.
Pour les potentiels des récepteurs, le seuil n'est pas un facteur, car la modification du potentiel membranaire des cellules réceptrices entraîne directement la libération de neurotransmetteurs. Cependant, les potentiels générateurs peuvent initier des potentiels d'action dans l'axone du neurone sensoriel, et les potentiels postsynaptiques peuvent initier un potentiel d'action dans l'axone d'autres neurones. Les potentiels gradués s'additionnent à un endroit précis au début de l'axone pour initier le potentiel d'action, à savoir le segment initial. Pour les neurones sensoriels, qui n'ont pas de corps cellulaire entre les dendrites et l'axone, le segment initial est directement adjacent aux terminaisons dendritiques. Pour tous les autres neurones, la butte axonale est essentiellement le segment initial de l'axone, et c'est là que se produit la sommation. Ces emplacements présentent une densité élevée de canaux Na + dépendants de la tension qui initient la phase de dépolarisation du potentiel d'action.

Figure\(\PageIndex{2}\) : Sommation du potentiel post-synaptique. Le résultat de la sommation des potentiels postsynaptiques est la variation globale du potentiel membranaire. Au point A, plusieurs potentiels post-synaptiques excitateurs différents s'additionnent pour former une dépolarisation importante. Au point B, une combinaison de potentiels postsynaptiques excitateurs et inhibiteurs donne un résultat final différent pour le potentiel membranaire.
La sommation peut être spatiale ou temporelle, c'est-à-dire qu'elle peut être le résultat de multiples potentiels gradués à différents endroits du neurone, ou tous au même endroit mais séparés dans le temps. La sommation spatiale est liée à l'association de l'activité de plusieurs entrées d'un neurone entre elles. La sommation temporelle est la relation entre les potentiels d'action multiples d'une seule cellule entraînant une modification significative du potentiel membranaire. La sommation spatiale et la sommation temporelle peuvent également agir ensemble.
Synapses
Il existe deux types de connexions entre les cellules électriquement actives, les synapses chimiques et les synapses électriques. Dans une synapse chimique, un signal chimique, à savoir un neurotransmetteur, est libéré par une cellule et affecte l'autre cellule. Dans une synapse électrique, il existe une connexion directe entre les deux cellules afin que les ions puissent passer directement d'une cellule à l'autre. Si une cellule est dépolarisée dans une synapse électrique, la cellule jointe se dépolarise également parce que les ions passent entre les cellules. Les synapses chimiques impliquent la transmission d'informations chimiques d'une cellule à l'autre. Cette section se concentrera sur le type chimique de synapse.
La jonction neuromusculaire (NMJ) décrite dans le chapitre sur le tissu musculaire est un exemple de synapse chimique. Dans le système nerveux, il existe de nombreuses autres synapses qui sont essentiellement les mêmes que celles du NMJ. Toutes les synapses ont des caractéristiques communes, qui peuvent être résumées dans cette liste :
- élément présynaptique
- neurotransmetteur (conditionné dans des vésicules)
- fente synaptique
- protéines réceptrices
- élément postsynaptique
- élimination ou recapture des neurotransmetteurs
Pour le NMJ, ces caractéristiques sont les suivantes : l'élément présynaptique est constitué des terminaisons axonales du motoneurone, le neurotransmetteur est l'acétylcholine, la fente synaptique est l'espace entre les cellules où le neurotransmetteur diffuse, la protéine réceptrice est le récepteur nicotinique de l'acétylcholine, le l'élément postsynaptique est le sarcolemme de la cellule musculaire et le neurotransmetteur est éliminé par l'acétylcholinestérase. D'autres synapses sont similaires et leurs spécificités sont différentes, mais elles contiennent toutes les mêmes caractéristiques.
Libération de neurotrans
Lorsqu'un potentiel d'action atteint les terminaisons axonales, les canaux Ca 2+ dépendants de la tension situés dans la membrane du bulbe synaptique s'ouvrent. La concentration de Ca 2+ augmente à l'intérieur du bulbe terminal, et l'ion Ca 2+ s'associe aux protéines de la surface externe des vésicules des neurotransmetteurs. Le Ca 2+ facilite la fusion de la vésicule avec la membrane présynaptique, de sorte que le neurotransmetteur est libéré par exocytose dans le petit espace entre les cellules, connu sous le nom de fente synaptique.
Une fois dans la fente synaptique, le neurotransmetteur diffuse la courte distance jusqu'à la membrane postsynaptique et peut interagir avec les récepteurs des neurotransmetteurs. Les récepteurs sont spécifiques au neurotransmetteur, et les deux s'emboîtent comme une clé et une serrure. Un neurotransmetteur se lie à son récepteur et ne se lie pas aux récepteurs d'autres neurotransmetteurs, faisant de cette liaison un événement chimique spécifique (Figure\(\PageIndex{3}\)).
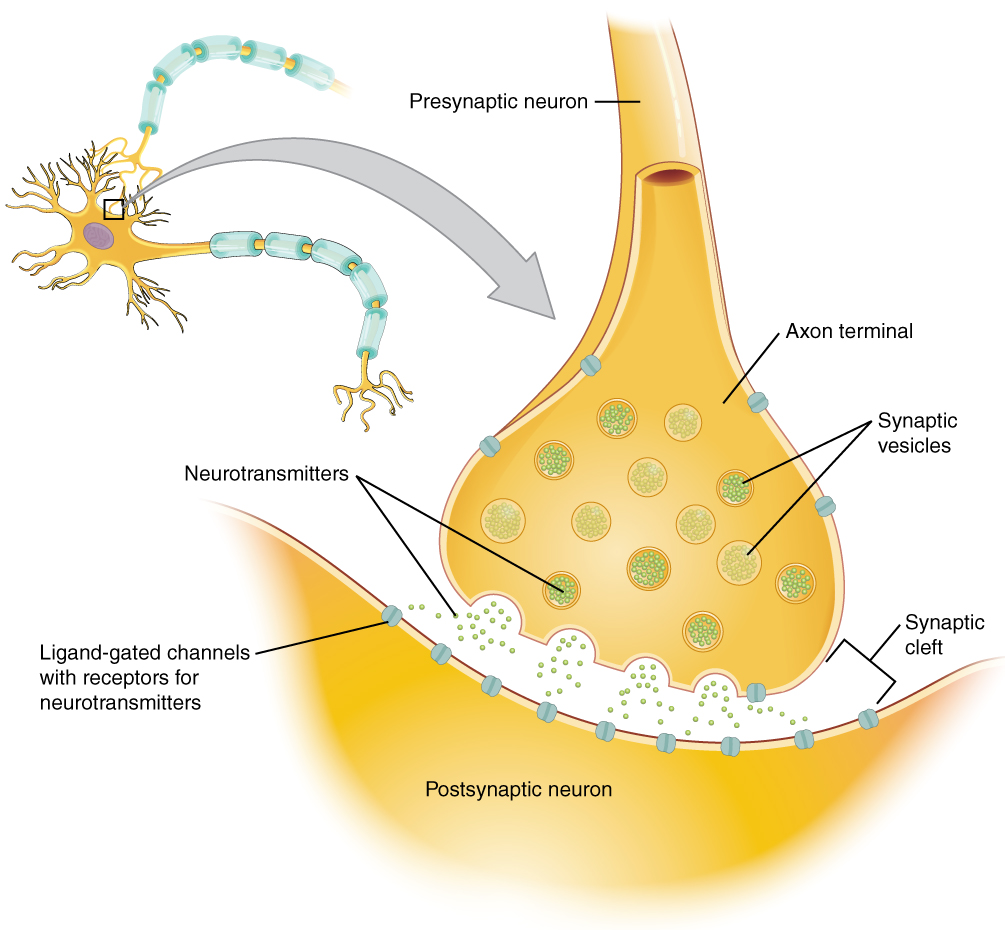
Figure\(\PageIndex{3}\) : La Synapse. La synapse est une connexion entre un neurone et sa cellule cible (qui n'est pas nécessairement un neurone). L'élément présynaptique est le bulbe synaptique terminal de l'axone où le Ca 2+ entre dans le bulbe pour provoquer la fusion des vésicules et la libération des neurotransmetteurs. Le neurotransmetteur diffuse à travers la fente synaptique pour se lier à son récepteur. Le neurotransmetteur est éliminé de la synapse soit par dégradation enzymatique, soit par recapture neuronale, soit par recapture gliale.
Systèmes neurotransmetteurs
Plusieurs systèmes de neurotransmetteurs se trouvent au niveau de diverses synapses du système nerveux. Ces groupes font référence aux substances chimiques qui sont les neurotransmetteurs et, au sein de ces groupes, se trouvent des systèmes spécifiques.
Le premier groupe, qui est un système neurotransmetteur à part entière, est le système cholinergique. C'est le système basé sur l'acétylcholine. Cela inclut le NMJ comme exemple de synapse cholinergique, mais des synapses cholinergiques se trouvent dans d'autres parties du système nerveux. Ils se trouvent dans le système nerveux autonome et sont répartis dans tout le cerveau.
Le système cholinergique possède deux types de récepteurs, le récepteur nicotinique se trouve dans le NMJ ainsi que dans d'autres synapses. Il existe également un récepteur de l'acétylcholine connu sous le nom de récepteur muscarinique. Ces deux récepteurs sont nommés d'après des médicaments qui interagissent avec le récepteur en plus de l'acétylcholine. La nicotine se lie au récepteur nicotinique et l'active de la même manière que l'acétylcholine. La muscarine, un produit de certains champignons, se lie au récepteur muscarinique. Cependant, la nicotine ne se lie pas au récepteur muscarinique et la muscarine ne se lie pas au récepteur nicotinique.
Les acides aminés constituent un autre groupe de neurotransmetteurs. Cela inclut le glutamate (Glu), le GABA (acide gamma-aminobutyrique, un dérivé du glutamate) et la glycine (Gly). Ces acides aminés possèdent un groupe amino et un groupe carboxyle dans leurs structures chimiques. Le glutamate est l'un des 20 acides aminés utilisés pour fabriquer des protéines. Chaque neurotransmetteur d'acides aminés ferait partie de son propre système, à savoir les systèmes glutamatergique, GABAergique et glycinergique. Ils ont chacun leurs propres récepteurs et n'interagissent pas entre eux. Les neurotransmetteurs d'acides aminés sont éliminés de la synapse par recapture. Une pompe placée dans la membrane cellulaire de l'élément présynaptique, ou parfois d'une cellule gliale voisine, éliminera l'acide aminé de la fente synaptique afin de le recycler, de le reconditionner dans des vésicules et de le libérer à nouveau.
Une autre classe de neurotransmetteurs est l'amine biogénique, un groupe de neurotransmetteurs fabriqués par voie enzymatique à partir d'acides aminés. Ils contiennent des groupes aminés, mais ne contiennent plus de groupes carboxyle et ne sont donc plus classés comme acides aminés. La sérotonine est fabriquée à partir de tryptophane. C'est la base du système sérotoninergique, qui possède ses propres récepteurs spécifiques. La sérotonine est renvoyée dans la cellule présynaptique pour être reconditionnée.
D'autres amines biogéniques sont fabriquées à partir de tyrosine, notamment la dopamine, la norépinéphrine et l'épinéphrine. La dopamine fait partie de son propre système, le système dopaminergique, qui possède des récepteurs de la dopamine. La dopamine est éliminée de la synapse par des protéines de transport dans la membrane cellulaire présynaptique. La norépinéphrine et l'épinéphrine appartiennent au système des neurotransmetteurs adrénergiques. Les deux molécules sont très similaires et se lient aux mêmes récepteurs, appelés récepteurs alpha et bêta. La norépinéphrine et l'épinéphrine sont également renvoyées dans la cellule présynaptique. L'épinéphrine (épi- = « activé » ; « -néphrine » = rein) est également connue sous le nom d'adrénaline (rénal = « rein »), et la norépinéphrine est parfois appelée noradrénaline. La glande surrénale produit de l'épinéphrine et de la noradrénaline qui sont libérées dans la circulation sanguine sous forme d'hormones.
Un neuropeptide est une molécule neurotransmettrice composée de chaînes d'acides aminés reliées par des liaisons peptidiques. C'est ce qu'est une protéine, mais le terme protéine implique une certaine longueur de la molécule. Certains neuropeptides sont assez courts, comme la mét-enképhaline, qui contient cinq acides aminés. D'autres sont longues, comme la bêta-endorphine, qui contient 31 acides aminés. Les neuropeptides sont souvent libérés au niveau des synapses en association avec un autre neurotransmetteur, et ils agissent souvent comme des hormones dans d'autres systèmes de l'organisme, tels que le peptide intestinal vasoactif (VIP) ou la substance P.
L'effet d'un neurotransmetteur sur l'élément postsynaptique dépend entièrement de la protéine réceptrice. Premièrement, s'il n'y a pas de protéine réceptrice dans la membrane de l'élément postsynaptique, le neurotransmetteur n'a aucun effet. L'effet dépolarisant ou hyperpolarisant dépend également du récepteur. Lorsque l'acétylcholine se lie au récepteur nicotinique, la cellule postsynaptique est dépolarisée. En effet, le récepteur est un canal cationique et le Na + chargé positivement se précipite dans la cellule. Cependant, lorsque l'acétylcholine se lie au récepteur muscarinique, dont il existe plusieurs variantes, elle peut provoquer une dépolarisation ou une hyperpolarisation de la cellule cible.
Les neurotransmetteurs d'acides aminés, le glutamate, la glycine et le GABA, sont presque exclusivement associés à un seul effet. Le glutamate est considéré comme un acide aminé excitateur, mais uniquement parce que les récepteurs Glu chez l'adulte provoquent une dépolarisation de la cellule postsynaptique. La glycine et le GABA sont considérés comme des acides aminés inhibiteurs, là encore parce que leurs récepteurs provoquent une hyperpolarisation.
Les amines biogéniques ont des effets mixtes. Par exemple, les récepteurs de la dopamine classés comme récepteurs D1 sont excitateurs alors que les récepteurs de type D2 sont inhibiteurs. Les récepteurs biogènes aux amines et aux neuropeptides peuvent avoir des effets encore plus complexes, car certains peuvent ne pas affecter directement le potentiel membranaire, mais avoir un effet sur la transcription des gènes ou d'autres processus métaboliques dans le neurone. Les caractéristiques des différents systèmes de neurotransmetteurs présentés dans cette section sont organisées dans le tableau\(\PageIndex{1}\).
L'important à retenir à propos des neurotransmetteurs, et des substances chimiques de signalisation en général, est que l'effet dépend entièrement du récepteur. Les neurotransmetteurs se lient à l'une des deux classes de récepteurs situés à la surface de la cellule, ionotropiques ou métabotropiques (Figure\(\PageIndex{4}\)). Les récepteurs ionotropes sont des canaux ioniques dépendants des ligands, tels que le récepteur nicotinique de l'acétylcholine ou le récepteur de la glycine. Un récepteur métabotropique implique un complexe de protéines qui entraînent des modifications métaboliques au sein de la cellule. Le complexe récepteur comprend la protéine réceptrice transmembranaire, une protéine G et une protéine effectrice. Le neurotransmetteur, appelé premier messager, se lie à la protéine réceptrice située à la surface extracellulaire de la cellule, et le côté intracellulaire de la protéine déclenche l'activité de la protéine G. La protéine G est une guanosine triphosphate (GTP) hydrolase qui passe physiquement de la protéine réceptrice à la protéine effectrice pour activer cette dernière. Une protéine effectrice est une enzyme qui catalyse la génération d'une nouvelle molécule, qui agit en tant que médiateur intracellulaire du signal qui se lie au récepteur. Ce médiateur intracellulaire est appelé le second messager.
Différents récepteurs utilisent différents seconds messagers. Deux exemples courants de seconds messagers sont l'adénosine monophosphate cyclique (AMPc) et l'inositol triphosphate (IP 3). L'enzyme adénylate cyclase (exemple de protéine effectrice) produit l'AMPc, et la phospholipase C est l'enzyme qui fabrique l'IP 3. Les seconds messagers, une fois produits par la protéine effectrice, provoquent des modifications métaboliques au sein de la cellule. Ces changements sont très probablement l'activation d'autres enzymes de la cellule. Dans les neurones, ils modifient souvent les canaux ioniques, en les ouvrant ou en les fermant. Ces enzymes peuvent également provoquer des modifications de la cellule, telles que l'activation de gènes dans le noyau, et donc l'augmentation de la synthèse des protéines. Dans les neurones, ces types de changements sont souvent à la base de connexions plus fortes entre les cellules de la synapse et peuvent être à la base de l'apprentissage et de la mémoire.

Figure\(\PageIndex{4}\) : Types de récepteurs. (a) Un récepteur ionotropique est un canal qui s'ouvre lorsque le neurotransmetteur s'y lie. (b) Un récepteur métabotropique est un complexe qui provoque des modifications métaboliques dans la cellule lorsque le neurotransmetteur s'y lie (1). Après liaison, la protéine G hydrolyse le GTP et se déplace vers la protéine effectrice (2). Lorsque la protéine G entre en contact avec la protéine effectrice, un second messager est généré, tel que l'AMPc (3). Le second messager peut ensuite provoquer des modifications du neurone, telles que l'ouverture ou la fermeture de canaux ioniques, des modifications métaboliques et des modifications de la transcription génique.
| Système | Cholinergique | Acides aminés | Amines biogéniques | Neuropeptides |
|---|---|---|---|---|
| neurotransmetteurs | Acétylcholine | Glutamate, glycine, GABA | Sérotonine (5-HT), dopamine, norépinéphrine, (épinéphrine) | Met-enképhaline, bêta-endorphine, VIP, substance P, etc. |
| récepteurs | Récepteurs nicotiniques et muscariniques | Récepteurs Glu, récepteurs Gly, récepteurs GABA | Récepteurs 5-HT, récepteurs D1 et D2, récepteurs α-adrénergiques et β-adrénergiques | Les récepteurs sont trop nombreux pour être listés, mais ils sont spécifiques aux peptides. |
| Élimination | Dégradation par l'acétylcholinestérase | Recapture par les neurones ou les cellules gliales | Recapture par les neurones | Dégradation par des enzymes appelées peptidases |
| Effet post-synaptique | Le récepteur nicotinique provoque une dépolarisation. Les récepteurs muscariniques peuvent provoquer à la fois une dépolarisation ou une hyperpolarisation selon le sous-type. | Les récepteurs Glu provoquent une dépolarisation. Les récepteurs Gly et GABA provoquent une hyperpolarisation. | La dépolarisation ou l'hyperpolarisation dépendent du récepteur spécifique. Par exemple, les récepteurs D1 provoquent une dépolarisation et les récepteurs D2 provoquent une hyperpolarisation. | La dépolarisation ou l'hyperpolarisation dépendent du récepteur spécifique. |
TROUBLES DE LA... Système nerveux
La cause sous-jacente de certaines maladies neurodégénératives, telles que la maladie d'Alzheimer et la maladie de Parkinson, semble être liée aux protéines, en particulier au mauvais comportement des protéines. L'une des théories les plus solides sur les causes de la maladie d'Alzheimer repose sur l'accumulation de plaques bêta-amyloïdes, des conglomérations denses d'une protéine qui ne fonctionne pas correctement. La maladie de Parkinson est liée à une augmentation de la concentration d'une protéine appelée alpha-synucléine qui est toxique pour les cellules du noyau de la substance noire du mésencéphale.
Pour que les protéines fonctionnent correctement, elles dépendent de leur forme tridimensionnelle. La séquence linéaire d'acides aminés se plie en une forme tridimensionnelle basée sur les interactions entre et parmi ces acides aminés. Lorsque le repliement est perturbé et que les protéines prennent une forme différente, elles cessent de fonctionner correctement. Mais la maladie n'est pas nécessairement le résultat d'une perte fonctionnelle de ces protéines ; ces protéines altérées commencent plutôt à s'accumuler et peuvent devenir toxiques. Par exemple, dans la maladie d'Alzheimer, la caractéristique de la maladie est l'accumulation de ces plaques amyloïdes dans le cortex cérébral. Le terme inventé pour décrire ce type de maladie est « protéopathie » et il inclut d'autres maladies. La maladie de Creutzfeld-Jacob, la variante humaine de la maladie à prion connue sous le nom de maladie de la vache folle chez les bovins, implique également l'accumulation de plaques amyloïdes, similaires à la maladie d'Alzheimer. Des maladies d'autres systèmes organiques peuvent également appartenir à ce groupe, telles que la mucoviscidose ou le diabète de type 2. La reconnaissance de la relation entre ces maladies a suggéré de nouvelles possibilités thérapeutiques. Interférer avec l'accumulation des protéines, et peut-être dès leur production initiale dans la cellule, pourrait permettre de découvrir de nouvelles façons de soulager ces maladies dévastatrices.
Révision du chapitre
La base du signal électrique à l'intérieur d'un neurone est le potentiel d'action qui se propage le long de l'axone. Pour qu'un neurone génère un potentiel d'action, il doit recevoir une entrée provenant d'une autre source, soit d'un autre neurone, soit d'un stimulus sensoriel. Cette entrée se traduira par l'ouverture de canaux ioniques dans le neurone, ce qui se traduira par un potentiel gradué en fonction de la force du stimulus. Les potentiels gradués peuvent être dépolarisants ou hyperpolarisants et peuvent s'additionner pour affecter la probabilité que le neurone atteigne le seuil.
Les potentiels gradués peuvent être le résultat de stimuli sensoriels. Si le stimulus sensoriel est reçu par les dendrites d'un neurone sensoriel unipolaire, tel que le neurone sensoriel se terminant dans la peau, le potentiel gradué est appelé potentiel générateur car il peut générer directement le potentiel d'action dans le segment initial de l'axone. Si le stimulus sensoriel est reçu par une cellule réceptrice sensorielle spécialisée, le potentiel gradué est appelé potentiel récepteur. Les potentiels gradués produits par les interactions entre les neurones au niveau des synapses sont appelés potentiels postsynaptiques (PSP). Un potentiel gradué dépolarisant au niveau d'une synapse est appelé PSP excitatrice, et un potentiel gradué hyperpolarisant au niveau d'une synapse est appelé PSP inhibitrice.
Les synapses sont les contacts entre les neurones, qui peuvent être de nature chimique ou électrique. Les synapses chimiques sont beaucoup plus courantes. Au niveau d'une synapse chimique, le neurotransmetteur est libéré par l'élément présynaptique et diffuse à travers la fente synaptique. Le neurotransmetteur se lie à une protéine réceptrice et provoque une modification de la membrane postsynaptique (PSP). Le neurotransmetteur doit être inactivé ou retiré de la fente synaptique afin que le stimulus soit limité dans le temps.
Les caractéristiques particulières d'une synapse varient en fonction du système de neurotransmetteurs produit par ce neurone. Le système cholinergique se trouve à la jonction neuromusculaire et à certains endroits du système nerveux. Les acides aminés tels que le glutamate, la glycine et l'acide gamma-aminobutyrique (GABA) sont utilisés comme neurotransmetteurs. D'autres neurotransmetteurs sont le résultat de modifications enzymatiques d'acides aminés, comme dans les amines biogéniques, ou de liaisons covalentes, comme dans les neuropeptides.
Questions sur les liens interactifs
Regardez cette vidéo pour en savoir plus sur la sommation. Le processus de conversion des signaux électriques en signaux chimiques et inversement nécessite des modifications subtiles qui peuvent entraîner des augmentations ou des diminutions transitoires de la tension de la membrane. Pour provoquer un changement durable dans la cellule cible, plusieurs signaux sont généralement additionnés ou sommés. La sommation spatiale doit-elle se produire en une seule fois, ou les signaux séparés peuvent-ils arriver sur le neurone postsynaptique à des moments légèrement différents ? Expliquez votre réponse.
Réponse : Un deuxième signal provenant d'un neurone présynaptique distinct peut arriver un peu plus tard, à condition qu'il arrive avant que le premier ne s'éteigne ou ne se dissipe.
Regardez cette vidéo pour en savoir plus sur la libération d'un neurotransmetteur. Le potentiel d'action atteint l'extrémité de l'axone, appelée terminaison axonale, et un signal chimique est émis pour indiquer à la cellule cible de faire quelque chose, soit d'initier un nouveau potentiel d'action, soit de supprimer cette activité. Dans un espace très court, le signal électrique du potentiel d'action est transformé en signal chimique d'un neurotransmetteur, puis en modifications électriques de la membrane cellulaire cible. Quelle est l'importance des canaux calciques dépendants de la tension dans la libération des neurotransmetteurs ?
Réponse : Le potentiel d'action dépolarise la membrane cellulaire du terminal axonal, qui contient le canal Ca 2+ dépendant de la tension. Ce changement de tension ouvre le canal afin que Ca 2+ puisse entrer dans la borne axonale. Les ions calcium permettent aux vésicules synaptiques de libérer leur contenu par exocytose.
Questions de révision
Q. Quelle est l'ampleur de la variation du potentiel membranaire nécessaire pour que la somme des potentiels postsynaptiques entraîne la génération d'un potentiel d'action ?
A. +30 mV
B. +15 mV
C. +10 mV
D. -15 mV
Réponse : B
Q. Un canal s'ouvre sur une membrane postsynaptique et provoque l'entrée d'un ion négatif dans la cellule. De quel type de potentiel gradué s'agit-il ?
A. dépolarisant
B. repolarisant
C. hyperpolarisant
D. non polarisant
Réponse : C
Q. Quel neurotransmetteur est libéré à la jonction neuromusculaire ?
A. norépinéphrine
B. sérotonine
C. dopamine
D. acétylcholine
Réponse : D
Q. Quel type de récepteur nécessite une protéine effectrice pour initier un signal ?
A. amine biogénique
B. récepteur ionotropique
C. système cholinergique
D. récepteur métabotropique
Réponse : D
Q. Lequel des neurotransmetteurs suivants est associé exclusivement à l'inhibition ?
A. GABA
B. acétylcholine
C. glutamate
D. norépinéphrine
Réponse : A
Questions sur la pensée critique
Q. Si une cellule postsynaptique possède des synapses provenant de cinq cellules différentes, que trois provoquent des EPSP et que deux d'entre elles provoquent des IPSP, donnez un exemple d'une série de dépolarisations et d'hyperpolarisations qui amèneraient le neurone à atteindre le seuil.
A. EPSP1 = +5 mV, EPSP2 = +7 mV, EPSP 3 = +10 mV, IPSP1 = -4 mV, IPSP2 = -3 mV. 5 + 7 + 10 — 4 — 3 = +15 mV.
Q. Pourquoi le récepteur est-il l'élément important qui détermine l'effet d'un neurotransmetteur sur une cellule cible ?
R. Les différents neurotransmetteurs ont des récepteurs différents. Ainsi, c'est le type de récepteur de la cellule postsynaptique qui détermine quels canaux ioniques s'ouvrent. La liaison de l'acétylcholine au récepteur nicotinique provoque la traversée de la membrane par les cations. La liaison du GABA à son récepteur provoque la traversée de la membrane par le chlorure d'anions.
Lexique
- amine biogénique
- classe de neurotransmetteurs dérivés enzymatiquement d'acides aminés mais ne contenant plus de groupe carboxyle
- synapse chimique
- connexion entre deux neurones, ou entre un neurone et sa cible, où un neurotransmetteur diffuse sur une très courte distance
- système cholinergique
- système neurotransmetteur de l'acétylcholine, qui comprend ses récepteurs et l'enzyme acétylcholinestérase
- protéine effectrice
- enzyme qui catalyse la génération d'une nouvelle molécule, qui agit en tant que médiateur intracellulaire du signal qui se lie au récepteur
- synapse électrique
- connexion entre deux neurones, ou deux cellules électriquement actives quelconques, où les ions circulent directement à travers des canaux recouvrant leurs membranes cellulaires adjacentes
- potentiel post-synaptique excitateur (EPSP)
- potentiel graduel dans la membrane postsynaptique qui résulte d'une dépolarisation et augmente la probabilité qu'un potentiel d'action se produise
- potentiel du générateur
- potentiel gradué provenant des dendrites d'une cellule unipolaire qui génère le potentiel d'action dans le segment initial de l'axone de cette cellule
- Protéine G
- la guanosine triphosphate (GTP) hydrolase qui passe physiquement de la protéine réceptrice à la protéine effectrice pour activer cette dernière
- potentiel postsynaptique inhibiteur (IPSP)
- potentiel graduel dans la membrane postsynaptique qui résulte d'une hyperpolarisation et réduit la probabilité qu'un potentiel d'action se produise
- récepteur métabotropique
- récepteur de neurotransmetteur qui implique un complexe de protéines qui provoquent des changements métaboliques dans une cellule
- récepteur muscarinique
- type de protéine réceptrice de l'acétylcholine qui se caractérise par une liaison avec la muscarine et qui est un récepteur métabotropique
- neuropeptide
- type de neurotransmetteur qui comprend des molécules de protéines et des chaînes plus courtes d'acides aminés
- récepteur nicotinique
- type de protéine réceptrice de l'acétylcholine qui se caractérise par une liaison également à la nicotine et qui est un récepteur ionotropique
- potentiel postsynaptique (PSP)
- potentiel graduel dans la membrane postsynaptique causé par la liaison du neurotransmetteur aux récepteurs protéiques
- potentiel de récepteur
- potentiel gradué dans une cellule sensorielle spécialisée qui provoque directement la libération d'un neurotransmetteur sans potentiel d'action intermédiaire
- sommation spatiale
- combinaison de potentiels graduels à travers la membrane cellulaire neuronale provoquée par des signaux provenant d'éléments présynaptiques distincts qui s'additionnent pour initier un potentiel d'action
- résumer
- à additionner, comme dans le cas de la variation cumulée des potentiels postsynaptiques vers l'atteinte du seuil dans la membrane, soit sur une partie de la membrane, soit sur une certaine période
- fente synaptique
- petit espace entre les cellules d'une synapse chimique où le neurotransmetteur diffuse de l'élément présynaptique à l'élément postsynaptique
- sommation temporelle
- combinaison de potentiels gradués au même endroit sur un neurone, produisant un signal puissant à partir d'une entrée