
13.3: Protists
- Page ID
- 199381

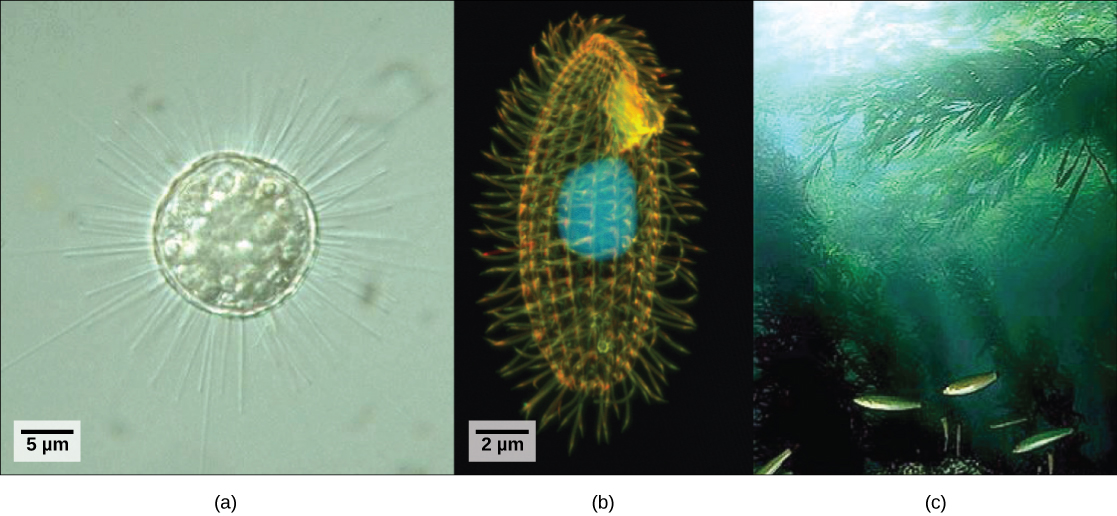
历史上不符合 Animalia、Fungi 或 Plantae 王国标准的真核生物被称为原生物,被归类为普罗蒂斯塔王国。 原生生物包括生活在池塘水中的单细胞真核生物(图\(\PageIndex{1}\)),尽管原生生物生活在各种其他水生和陆地环境中,并且占据了许多不同的利基市场。 并非所有的原生生物都是微观的单细胞物种;有一些非常大的多细胞物种,例如海带。 在过去的二十年中,分子遗传学领域已经证明,一些原生生物与动物、植物或真菌的关系比与其他原生生物的关系更密切。 出于这个原因,最初归类为普罗蒂斯塔王国的抗议者血统已被重新分配到新的王国或其他现有王国。 原生生物的进化谱系仍在研究和辩论中。 同时,“protist” 一词仍然被非正式地用来描述这个极其多样化的真核生物群体。 作为一个集体群体,原生生物表现出惊人的形态、生理和生态多样性。
原生动物的特征
有超过100,000种描述的原生生物活物种,目前尚不清楚可能存在多少未描述的物种。 由于许多原生生物与其他生物生活在共生关系中,而且这些关系通常是特定物种的,因此存在与宿主多样性相匹配的未描述的原生生物多样性的巨大潜力。 作为非动物、植物、真菌或任何单一系统发育相关群体的真核生物的统称,所有原生生物几乎没有共同的特征也就不足为奇了。
几乎所有原生生物都存在于某种类型的水生环境中,包括淡水和海洋环境、潮湿的土壤甚至雪。 有几种抗议物种是感染动物或植物的寄生虫。 寄生虫是一种生活在另一个生物体之上或之内并以其为食的生物,通常不会杀死它。 一些抗议物种靠死亡的生物或其废物为生,并促成了它们的腐烂。
Protist 结构
原生生物的细胞是所有细胞中最精细的细胞之一。 大多数原生生物是微观和单细胞的,但存在一些真正的多细胞形式。 一些原生生物以菌落的形式生活,它们在某些方面表现为一组自由生活的细胞,而在其他方面则表现为多细胞生物。 还有其他原生生物是由巨大的、多核的单细胞组成的,它们看起来像无定形的粘液斑点,或者在其他情况下像蕨类植物。 实际上,许多原生细胞是多核化的;在某些物种中,细胞核的大小不同,在原生细胞功能中起着不同的作用。
单原细胞的大小从小于一微米到海藻 C aulerpa 的多核细胞长度不等。 Protist 细胞可能被动物样细胞膜或植物样细胞壁所包围。 其他的则被包裹在玻璃状的二氧化硅基外壳中,或者用互锁的蛋白质条薄膜包裹。 pellicle 的功能就像一层柔韧的盔甲,在不影响其活动范围的情况下防止抗议者被撕裂或刺穿。
大多数原生生物都是活动性的,但不同类型的原生生物进化出不同的运动模式。 有些原生生物有一根或多根鞭毛,它们会旋转或鞭打。 其他人则被成排或一簇的小纤毛覆盖,他们协调打这些纤毛才能游泳。 还有一些人从细胞的任何地方发出类似叶片的伪足虫,将伪足类固定在基质上,然后将细胞的其余部分拉向锚点。 一些原生生物可以通过将他们的运动策略与光感器官相结合来向光移动。
原生生物如何获得能量
原生动物表现出多种形式的营养,可能是有氧或厌氧的。 光合原生物(photoautotrophs)的特征是叶绿体的存在。 其他原生生物是异养生物,它们消耗有机物质(例如其他生物)来获取营养。 变形虫和其他一些异养原生物通过一种称为吞噬作用的过程摄取颗粒,在这种过程中,细胞膜会吞没食物颗粒并将其带入内部,挤出称为食物液泡的细胞内膜囊或囊泡(图\(\PageIndex{2}\))。 然后,这个囊泡与溶酶体融合,食物颗粒被分解成小分子,这些小分子可以扩散到细胞质中并用于细胞新陈代谢。 未消化的残留物最终会通过 exocytosis 从细胞中排出。
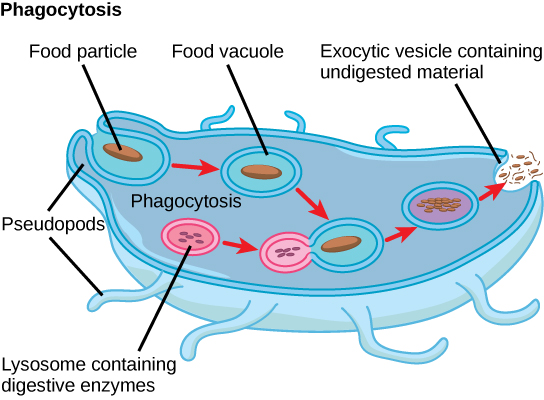
一些异养生物从死亡生物或其有机废物中吸收营养,而另一些异养生物则能够根据条件使用光合作用或以有机物为食。
繁殖
原生生物通过多种机制繁殖。 大多数都能够进行某种形式的无性繁殖,例如二元裂变产生两个子细胞,或者多次裂变同时分裂成许多子细胞。 其他人会产生微小的芽,这些芽会继续分裂并长到亲本抗议者的大小。 有性生殖,包括减数分裂和受精,在原生生物中很常见,许多原生生物可以在必要时从无性生殖转为有性生殖。 有性生殖通常与养分耗尽或环境发生变化的时期有关。 有性生殖可能使原生生物重组基因,产生新的后代变异,这些变异可能更适合在新环境中生存。 但是,有性生殖也通常与处于保护性休息阶段的囊肿有关。 根据其栖息地的不同,囊肿可能特别耐受极端温度、干燥或低 pH 值。 这种策略还允许某些原生生物 “等待” 压力源,直到他们的环境变得更有利于生存,或者直到他们被带到不同的环境(例如通过风、水或通过更大的生物体运输),因为囊肿几乎没有细胞新陈代谢。
Protist 多样性
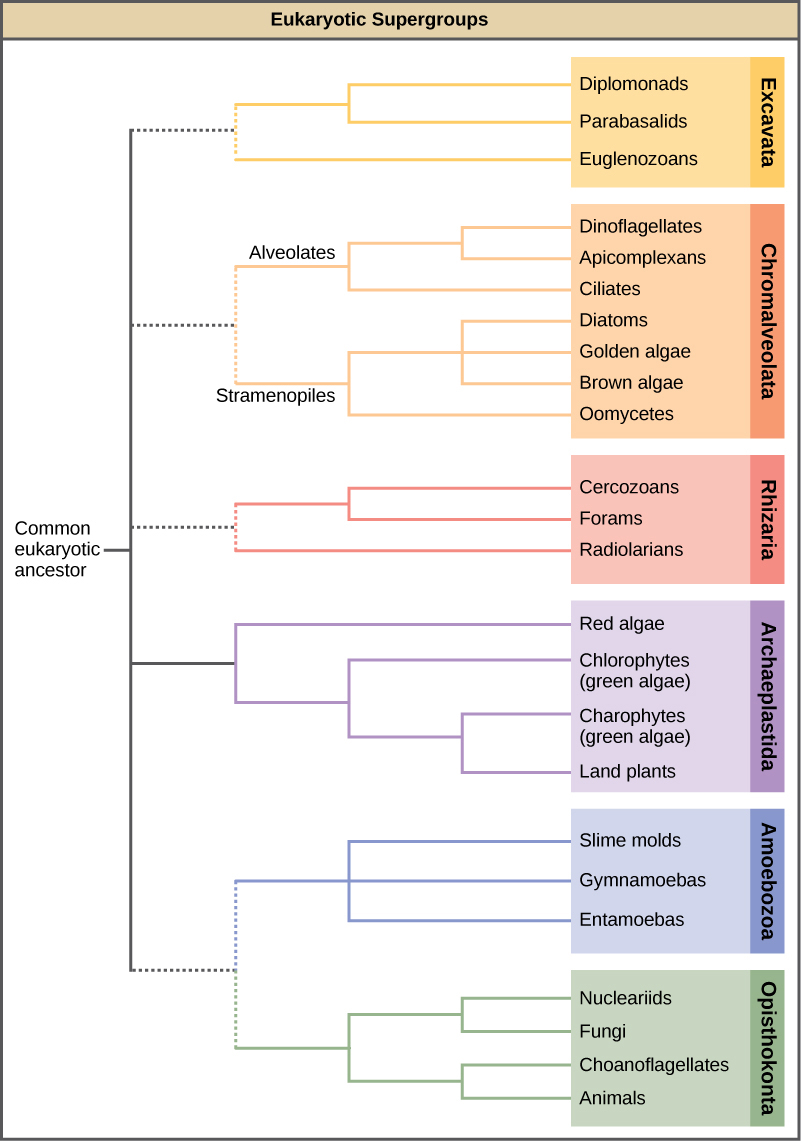
随着DNA测序的出现,原生群体之间以及原生群体与其他真核生物之间的关系开始变得更加清晰。 许多基于形态相似性的关系正在被基于遗传相似性的新关系所取代。 表现出相似形态特征的原生生物之所以进化出类似的结构,可能是因为相似的选择压力,而不是因为最近的共同血统。 这种现象被称为趋同进化。 这也是原教徒分类如此具有挑战性的原因之一。 新兴的分类方案将整个域 Eukaryota 分为六个 “超级组”,其中包含所有原生生物以及动物、植物和真菌(图\(\PageIndex{3}\));其中包括 Exculata、Chromalveolata、Rhizaria、Archeplastida、Amoebozoa 和 Opisthokonta。 超级群被认为是单系的;据信每个超级群中的所有生物都是从同一个共同祖先进化而来的,因此,与该群体之外的生物相比,所有成员之间的关系最为密切。 仍然缺乏证据证明某些团体存在垄断地位。
人类病原体
许多原生虫是致病性寄生虫,它们必须感染其他生物才能存活和繁殖。 Protist 寄生虫包括人类疟疾、非洲昏睡病和水传播胃肠炎的病原体。 其他抗议者病原体捕食植物,对粮食作物造成大规模破坏。
疟原虫种类
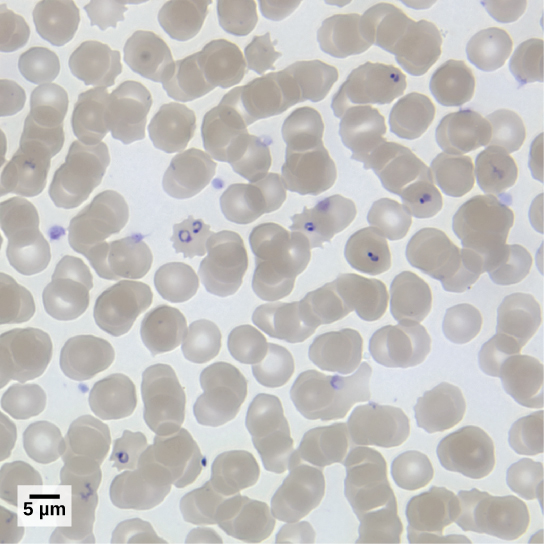
疟原虫属的成员必须感染蚊子和脊椎动物才能完成其生命周期。 在脊椎动物中,寄生虫在肝细胞中发育并继续感染红细胞,在每个无性复制周期中从血细胞中爆裂并破坏血细胞(图\(\PageIndex{4}\))。 在已知会感染人类的四种疟原虫物种中,P. 恶性疟原虫占所有疟疾病例的50%,是世界热带地区与疾病相关的死亡的主要原因。 据估计,2010年,疟疾造成50万至100万人死亡,其中大多数是非洲儿童。 在疟疾过程中,P. 恶性疟原虫可以感染和破坏人类一半以上的循环血细胞,从而导致严重的贫血。 为了应对寄生虫从受感染血细胞中爆发时释放的废物,宿主免疫系统会产生巨大的炎症反应,诱发谵妄的发热,因为寄生虫会破坏红细胞,将寄生虫废物泄漏到血液中。 P。 恶性疟原虫由非洲疟蚊冈比亚按蚊传播给人类。 杀死、消毒或避免接触这种极具侵略性的蚊子的技术,对于控制疟疾至关重要。
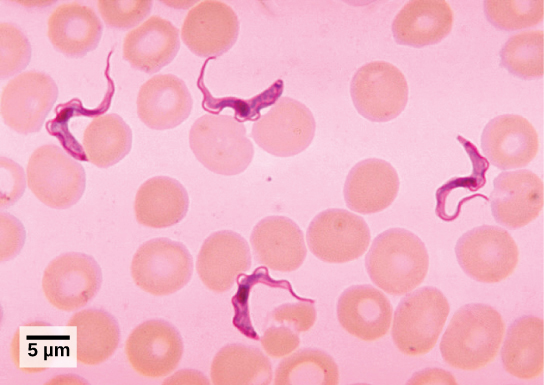
锥虫
T。 布鲁西是导致非洲昏睡病的寄生虫,它在每个传染周期中都会改变其厚厚的表面糖蛋白层,从而混淆人体免疫系统(图\(\PageIndex{5}\))。 糖蛋白被免疫系统识别为异物,并对寄生虫进行特异性抗体防御。 但是,T. 布鲁西有成千上万种可能的抗原,在接下来的每一代中,原生生物都会转向具有不同分子结构的糖蛋白涂层。 通过这种方式,T. brucei 能够在免疫系统无法成功清除寄生虫的情况下持续复制。 如果不进行治疗,非洲昏睡病必然会导致死亡,因为它会损害神经系统。 在疫情期间,该疾病的死亡率可能很高。 加强监测和控制措施导致报告病例减少;自2009年以来,已报告的病例数是50年来最低的(在整个撒哈拉以南非洲地区少于10,000例)。
在拉丁美洲,该属中的另一种物种 T. 克鲁兹,是恰加斯病的原因。 T。 cruzi 感染主要由吸血虫引起。 这种寄生虫在慢性感染阶段栖息在心脏和消化系统组织中,导致心律异常引起的营养不良和心力衰竭。 据估计,有1000万人感染了恰加斯病,该病在2008年造成1万人死亡。
植物寄生虫
陆生植物的Protist 寄生虫包括破坏粮食作物的病原体。 oomycete Plasmopara viticola 寄生葡萄植物,导致一种叫做霜霉病的疾病(图\(\PageIndex{6}\) a)。 感染了磷的葡萄植物 viticola 显得发育迟缓,枯萎的叶子变色。 霜霉病的蔓延导致法国葡萄酒业在十九世纪几乎崩溃。

P@@ hytophthora infestans 是一种导致马铃薯晚疫病的 oomycete,它会导致马铃薯茎和茎腐烂成黑粘液(图\(\PageIndex{6}\) b)。 由 P 引起的广泛马铃薯疫病 在十九世纪,侵扰者引发了众所周知的爱尔兰马铃薯饥荒,这场饥荒夺走了大约100万人的生命,并导致至少100万人从爱尔兰移民。 在美国和俄罗斯的某些地区,晚疫病继续困扰着马铃薯作物,在不使用杀虫剂的情况下,多达70%的农作物被消灭。
有益的原生生物
原生生物作为生产者扮演着至关重要的生态角色,尤其是在世界海洋中。 它们在食物网的另一端与分解剂同样重要。
原生生物作为食物来源
原生生物是许多其他生物的重要营养来源。 在某些情况下,例如浮游生物,原生生物是直接食用的。 或者,光合原生生物通过固碳为其他生物提供营养。 例如,被称为 zooxanthellae 的光合作用 dinoflagellates 将它们的大部分能量传递给容纳它们的珊瑚息肉(图\(\PageIndex{7}\))。 在这种互惠互利的关系中,息肉为虫黄藻提供了保护环境和营养。 息肉分泌形成珊瑚礁的碳酸钙。 如果没有 dinoflagellate 共生体,珊瑚会在一种称为珊瑚漂白的过程中失去藻类色素,最终死亡。 这就解释了为什么造礁珊瑚不生活在深度超过20米的水域中:到达这些深度的光线不足以让二鞭毛虫进行光合作用。

原生生物本身及其光合作用产物对于从细菌到哺乳动物等生物的生存至关重要,无论是直接还是间接地。 作为初级生产者,原生生物为世界上很大一部分的水生物种提供食物。 (在陆地上,陆地植物是初级生产者。) 事实上,世界上大约四分之一的光合作用是由原生生物进行的,尤其是 dinoflagellates、硅藻和多细胞藻类。
原生生物不会只为海栖生物创造食物来源。 例如,某些厌氧物种存在于白蚁和食木蟑螂的消化道中,它们有助于消化这些昆虫在穿过木材时摄入的纤维素。 用于消化纤维素的实际酶实际上是由生活在原生细胞中的细菌产生的。 白蚁为原生生物及其细菌提供食物来源,而原生生物和细菌通过分解纤维素为白蚁提供营养。
分解剂
许多类真菌的原生生物是腐烂体,以死亡生物或生物产生的废物(腐生菌是等效术语)为食的生物,专门从非生物有机物中吸收营养。 例如,许多类型的 oomycetes 生长在死亡的动物或藻类身上。 Saprobic protists 具有将无机养分返回土壤和水中的基本功能。 这个过程允许新的植物生长,这反过来又为食物链中的其他生物提供生计。 事实上,如果没有原生物、真菌和细菌等腐烂物种,生命将不复存在,因为所有有机碳都被 “捆绑在一起” 在死亡生物体中。
章节摘要
原生生物在生物学和生态学特征方面极其多样化,这在很大程度上是由于它们是系统发育不相关的群体的人工组合。 原生生物表现出高度多样的细胞结构、多种类型的繁殖策略、几乎所有可能的营养类型和不同的栖息地。 大多数单细胞原生生物都是活动性的,但这些生物使用不同的结构进行运输。
将原生生物分为有意义的群体的过程正在进行中,但过去20年的遗传数据澄清了许多以前不清楚或错误的关系。 目前大多数人的观点是将所有真核生物分为六个超级组。 该分类方案的目标是创建所有源自共同祖先的物种群。
词汇表
- Amoebozoa
- 包含变形虫和粘液霉菌的真核生物超级群
- Archeplastida
- 包含陆地植物、绿藻和红藻的真核生物超级群
- Chromalveolata
- 含有 dinoflagellates、纤毛动物、褐藻、硅藻和水霉的真核超级群
- 挖掘塔
- 真核生物超级群体,包含带有喂食槽的鞭毛单细胞生物
- Opisthokonta
- 含有真菌、动物和 choanoflagellates 的真核生物超级群
- 寄生物
- 一种生活在另一个生物体之上或之内并以其为食的生物,通常不会杀死它
- pellicle
- 一种由互锁的蛋白质条组成的外层细胞覆盖物,其功能类似于一层柔韧的盔甲,在不影响细胞活动范围的情况下防止细胞被撕裂或刺穿
- 根瘤
- 真核生物超级群体,包含通过变形虫运动移动的生物
- saprobe
- 一种以死有机物质为食的生物