
13.2: 真核生物起源
- Page ID
- 199366

化石记录和遗传证据表明,原核细胞是地球上最早的生物。 这些细胞起源于大约35亿年前,也就是地球形成大约10亿年后,在大约21亿年前真核细胞出现之前,它们是地球上唯一的生命形式。 在原核生物统治时期,光合原核生物进化而来,能够利用来自阳光的能量从二氧化碳和电子源(例如氢气、硫化氢或水)中合成有机材料(如碳水化合物)。
使用水作为电子供体的光合作用消耗二氧化碳并释放分子氧(O 2)作为副产物。 数百万年来,光合细菌的功能逐渐使地球的水饱和氧气,然后为大气充氧,而大气层以前所含的二氧化碳浓度要高得多,氧气浓度要低得多。 那个时代的老厌氧原核生物无法在新的有氧环境中发挥作用。 一些物种死亡,而另一些则在地球上剩余的厌氧环境中幸存下来。 还有其他早期的原核生物进化出机制,例如有氧呼吸,通过使用氧气储存有机分子中所含的能量来利用含氧气层。 有氧呼吸是从有机分子中获取能量的更有效方法,这为这些物种的成功做出了贡献(当今生活在地球上的有氧生物的数量和多样性就证明了这一点)。 有氧原核生物的进化是朝着第一个真核生物进化迈出的重要一步,但其他几个显著特征也必须进化。
内共生
在林恩·马古利斯(Lynn Margulis)于1960年代全面研究了一个革命性假设之前,真核细胞的起源在很大程度上还是个谜。 endosymbiotic 理论指出,真核生物是一个原核细胞吞没另一个原核细胞,一个生活在另一个原核细胞中,并随着时间的推移共同进化,直到单独的细胞无法再被识别出来。 这个曾经具有革命性的假设立即具有说服力,现在已被广泛接受,揭示这一进化过程所涉及的步骤以及关键人物的工作正在取得进展。 很明显,许多核真核生物基因和负责复制和表达这些基因的分子机制似乎与古细菌密切相关。 另一方面,代谢细胞器和负责许多能量收集过程的基因起源于细菌。 关于这种关系是如何发生的,还有很多需要澄清的地方;这仍然是生物学中一个令人兴奋的发现领域。 几次内共生事件可能促成了真核细胞的起源。
线粒体
真核细胞可能包含一到几千个线粒体,具体取决于细胞的能量消耗水平。 每个线粒体的长度为 1 到 10 微米,以移动、融合和分裂的长方形球体形式存在于细胞中(图\(\PageIndex{1}\))。 但是,线粒体无法在细胞外存活。 随着光合作用为大气充氧,以及成功的有氧原核生物进化,有证据表明,祖先的细胞吞没了自由生活的有氧原核生物并使其存活。 这使宿主细胞能够利用氧气释放储存在营养物质中的能量。 有几条证据支持线粒体源于这种内共生事件。 线粒体的形状像一组特定的细菌,周围环绕着两个膜,当一个膜结合的生物被另一个膜结合的生物吞没时,就会产生这种情况。 线粒体内膜包含大量的折叠物或 cristae,类似于某些细菌的纹理外表面。
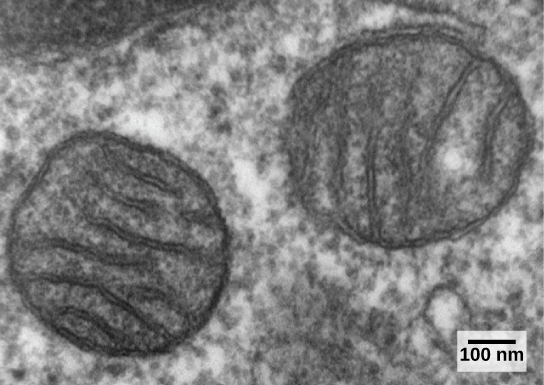
线粒体通过类似于原核生物中二元裂变的过程自行分裂。 线粒体有自己的圆形 DNA 染色体,其携带的基因与细菌表达的基因相似。 线粒体也有特殊的核糖体和转移 RNA,类似于原核生物中的这些成分。 这些特征都支持线粒体曾经是自由生活的原核生物。
叶绿体
叶绿体是一种质体,是植物细胞中的一组相关细胞器,参与淀粉、脂肪、蛋白质和色素的储存。 叶绿体含有绿色色素叶绿素,在光合作用中起作用。 遗传和形态学研究表明,质体是由吞没光合蓝细菌的祖先细胞的内共生关系进化而来的。 质体的大小和形状与蓝细菌相似,被两个或多个膜包裹,对应于蓝细菌的内膜和外膜。 像线粒体一样,质体也含有圆形基因组,其分裂过程让人联想到原核细胞分裂。 红藻和绿藻的叶绿体表现出与光合蓝藻密切相关的DNA序列,这表明红藻和绿藻是这种内共生事件的直接后代。
线粒体很可能在质体之前进化,因为所有真核生物都具有功能性线粒体或线粒体样细胞器。 相比之下,质体仅存在于一部分真核生物中,例如陆地植物和藻类。 图中总结了导致第一个真核生物的进化步骤的一个假设\(\PageIndex{2}\)。
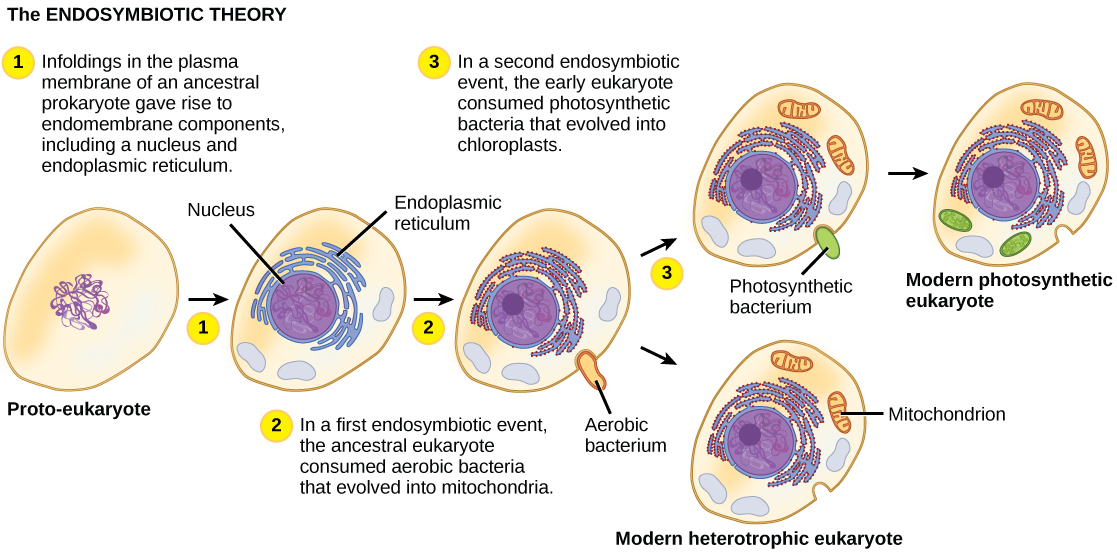
导致第一个真核细胞的确切步骤只能假设,关于哪些事件实际发生以及以什么顺序发生了一些争议。 据推测,螺旋体细菌会产生微管,而鞭毛的原核生物可能是真核鞭毛和纤毛的原材料。 其他科学家认为,导致线粒体和质体的发育的是膜增殖和分隔,而不是内共生事件。 但是,绝大多数研究都支持真核生物进化的内共生假说。
早期的真核生物像今天的大多数原生生物一样是单细胞的,但是随着真核生物变得越来越复杂,多细胞的进化使细胞在保持小巧的同时仍然表现出特殊的功能。 据认为,当今多细胞真核生物的祖先是在大约15亿年前进化出来的。
章节摘要
第一批真核生物是从祖先的原核生物进化而来的,其过程涉及膜增殖、细胞壁的丧失、细胞骨架的进化以及细胞器的获取和进化。 核真核生物基因似乎起源于古细菌,而真核细胞的能量机制似乎起源于细菌。 线粒体和质体起源于内共生事件,当时祖先细胞吞没了有氧细菌(就线粒体而言)和光合细菌(就叶绿体而言)。 线粒体的进化可能早于叶绿体的进化。 有证据表明,继发的内共生事件中,质体似乎是先前的内共生事件之后内共生的结果。
词汇表
- endosymbiosis
- 一个细胞被另一个细胞吞没,使被吞没的细胞存活下来,两个细胞都受益;这是导致真核生物中线粒体和叶绿体进化的过程
- plastid
- 植物细胞中参与淀粉、脂肪、蛋白质和色素储存的一组相关细胞器之一