
11.3:进化的证据
- Page ID
- 199123

进化的证据令人信服且广泛。 从生命系统的各个组织层面来看,生物学家看到了过去和现在进化的标志。 达尔文在他的著作《论物种起源》中用了很大一部分,确定了自然界中与进化相一致的模式,自达尔文以来,我们的理解变得更加清晰和广泛。
化石
化石提供了确凿的证据,表明过去的生物与今天发现的生物不同;化石显示出进化的进展。 科学家确定化石的年龄,并将其在世界各地进行分类,以确定这些生物何时相对于彼此生活。 由此产生的化石记录讲述了过去的故事,并显示了数百万年来形态的演变(图\(\PageIndex{1}\))。 例如,已经找到了鲸鱼和现代马进化过程中物种序列的非常详细的化石记录。 北美马的化石记录特别丰富,其中许多都含有过渡化石:这些化石显示出介于早期和晚期形态之间的中间解剖结构。 化石记录可以追溯到大约5500万年前的狗状祖先,该祖先在5500万至4200万年前在 Eohippus 属中产生了第一个类似马的物种。 这一系列化石追踪了解剖结构的变化,这种变化源于逐渐干燥的趋势,这种趋势将景观从森林覆盖的环境变成了草原。 连续的化石显示了牙齿形状以及足部和腿部解剖结构向放牧习惯的演变,并适应了逃离捕食者的需求,例如在4000万至3000万年前发现的 Mesohippus 物种中。 后来的物种体积有所增加,例如 Hipparion 的物种,它存在于大约2,300万至200万年前。 化石记录显示,马系中有几种适应性辐射,现在已经大大减少到只有一个属,即马属,有几个物种。

解剖学和胚胎学
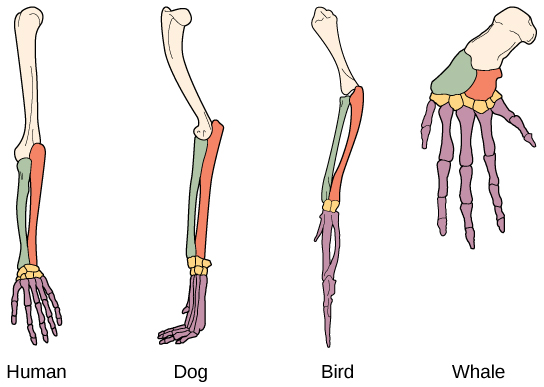
进化的另一种证据是生物体中存在具有相同基本形态的结构。 例如,人、狗、鸟和鲸的附属物中的骨头都具有相同的整体结构(图\(\PageIndex{2}\))。 这种相似之处源于它们起源于共同祖先的附属物。 随着时间的推移,进化导致不同物种中这些骨骼的形状和大小发生变化,但它们的总体布局保持不变,这证明了它们是共同祖先的血统。 科学家称这些同义部分为同源结构。 有些结构存在于完全没有明显功能的生物体中,似乎是过去祖先的残留部分。 例如,有些蛇尽管没有腿,但还是有骨盆骨,因为它们是有腿的爬行动物的后代。 这些没有功能的未使用结构称为残余结构。 其他残留结构的例子包括不会飞的鸟的翅膀(可能有其他功能)、某些仙人掌上的树叶、鲸鱼骨盆骨的痕迹以及洞穴动物看不见的眼睛。
进化的另一个证据是共享相似环境的生物的形态趋同。 例如,生活在北极地区的无关动物物种,例如北极狐和雷鸟(一只鸟),在冬季会有临时的白色覆盖物,以便与冰雪融为一体(图\(\PageIndex{3}\)). The similarity occurs not because of common ancestry, indeed one covering is of fur and the other of feathers, but because of similar selection pressures—the benefits of not being seen by predators.

胚胎学,即对生物体解剖结构发展到成体形态的研究,也为现在差异很大的生物群体之间存在关联性提供了证据。 某些群体中不存在的结构通常以其胚胎形式出现,并在达到成体或幼体形态时消失。 例如,所有脊椎动物胚胎,包括人类,在早期发育的某个时候都表现出鳃缝。 它们在陆生群体的成年人中消失,但在鱼类和一些两栖动物等成虫形式的水生群体中得以维持。 大猿胚胎,包括人类,在发育过程中具有尾巴结构,在出生时会消失。 不相关物种的胚胎往往相似的原因是,即使胚胎的相似性得以保留,在胚胎发育过程中影响生物体的突变变化也可能导致成虫体内的扩增差异。
生物地理学
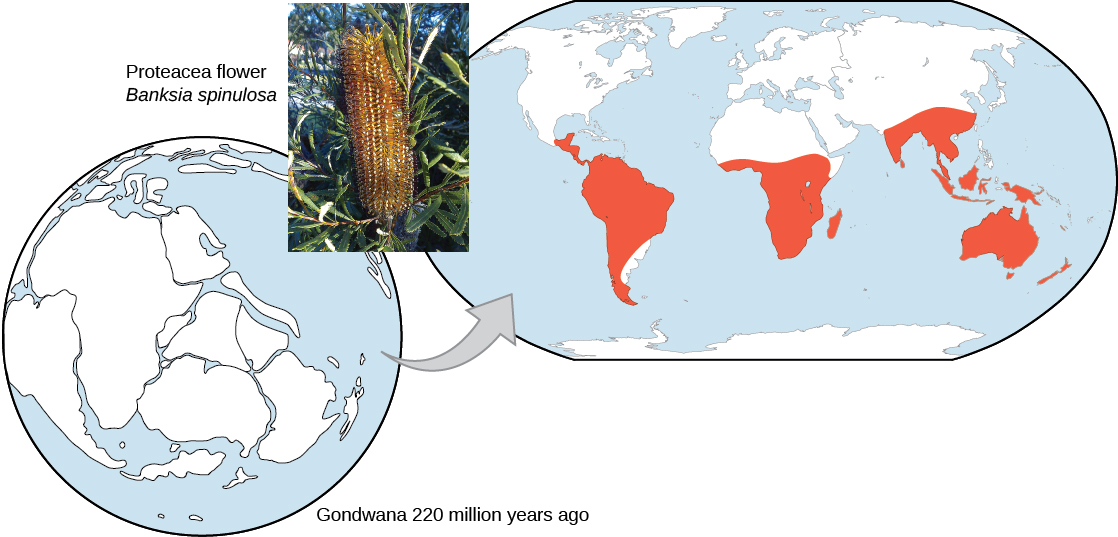
地球上生物的地理分布遵循的模式最能通过演化以及地质时期构造板块的运动来解释。 在超大陆Pangea解体(大约2亿年前)解体之前进化的广泛群体分布在世界各地。 自解体以来进化的群体在地球各区域独一无二地出现,例如由超大陆劳拉西亚形成的北方大陆的独特动植物群和由冈瓦纳超大陆形成的南方大陆的独特动植物。 在南方超大陆冈瓦纳解体之前,Proteaceae 存在于澳大利亚、南部非洲和南美最能解释该植物家族的存在(图\(\PageIndex{4}\))。
澳大利亚有袋动物的巨大多样化以及其他哺乳动物的缺乏反映了这个岛屿大陆的长期孤立。 澳大利亚拥有丰富的特有物种——其他任何地方都找不到的物种——这是岛屿的典型特征,这些岛屿被大片水域隔离,阻碍了物种迁移到其他地区。 随着时间的推移,这些物种在进化上分化为新的物种,这些物种看起来与可能存在于大陆的祖先截然不同。 澳大利亚的有袋动物、加拉帕戈斯群岛上的雀科和夏威夷群岛上的许多物种都只能在岛上找到,但与大陆的祖先物种却有着遥远的关系。
分子生物学
与解剖结构一样,生命分子的结构通过修改反映了血统。 DNA作为遗传物质的普遍性以及遗传密码和DNA复制和表达机制的近乎普遍性反映了所有生命都有共同祖先的证据。 这三个领域之间生命的基本分歧反映在原本保守的结构(例如核糖体成分和膜结构)的重大结构差异上。 总的来说,生物群体的关联性反映在它们的 DNA 序列的相似性上,这正是共同祖先的血统和多样化所期望的模式。
DNA序列也揭示了一些进化机制。 例如,很明显,蛋白质新功能的演变通常发生在基因复制事件之后。 这些重复是一种突变,其中整个基因被添加为基因组中的额外拷贝(或许多拷贝)。 这些复制允许通过突变、选择和漂移自由修改一个拷贝,而第二个拷贝继续产生功能蛋白。 这允许保留蛋白质的原始功能,而进化力量会调整副本,直到它以新的方式发挥作用。
章节摘要
进化的证据存在于生物的各个组织层面以及我们通过化石认识的灭绝物种中。 化石通过现已灭绝的形式为进化变化提供了证据,这些变化导致了现代物种的产生。 例如,有丰富的化石记录显示了从马祖向现代马匹的进化过渡,这些转变记录了中间形态以及对不断变化的生态系统的逐渐适应。 物种的解剖结构和该解剖结构的胚胎学发育揭示了不同谱系中的共同结构,这些结构随着时间的推移被进化而改变。 活物种的地理分布反映了特定地理位置物种的起源和大陆运动的历史。 分子的结构,如解剖结构,反映了活物种之间的关系,并将血统所预期的相似模式与修改相匹配。
词汇表
- 残余结构
- 一种存在于生物体中但没有明显功能的物理结构,似乎来自遥远祖先的功能结构