
11.4: 物种形成
- Page ID
- 199138

物种的生物学定义适用于有性繁殖的生物,是一组实际或可能杂交的个体。 根据这个定义,一个物种与另一个物种的区别在于每个物种的个体之间有可能交配以产生可育的后代。 此规则有例外。 许多物种非常相似,因此杂交后代是可能的,并且可能经常出现在自然界中,但是对于大多数物种来说,这条规则通常成立。 实际上,相似物种之间存在杂交表明它们可能是单一杂交物种的后代,物种形成过程可能尚未完成。
鉴于地球上生命的非凡多样性,必须有物种形成机制:从一个原始物种中形成两个物种。 达尔文将这个过程设想为分支事件,并在《论物种起源》(图\(\PageIndex{1}\) a)中找到的唯一插图中描绘了这个过程。 要实现物种形成,必须从一个原始种群中形成两个新种群,并且它们的进化方式必须使来自两个新种群的个体无法杂交。 生物学家提出了可能发生这种情况的机制,分为两大类。 Allopatric 物种形成,意思是 “其他家园” 中的物种形成,涉及种群与亲本物种的地理分离以及随后的进化。 Sympatric 物种形成,意思是 “同一个家园” 中的物种形成,涉及在亲本物种内部发生的物种形成,同时停留在一个地点。
生物学家认为物种形成事件是将一个祖先物种分裂成两个后代物种。 没有理由为什么一次形成的物种不超过两个,只是这种可能性较小,而且这种多重事件也可以概念化为近距离发生的单一分裂。

通过地理分离形成形态
地理上连续的人群拥有相对同质的基因库。 基因流,即等位基因在物种范围内的移动,相对自由,因为个体可以移动,然后在新的位置与个体交配。 因此,分布一端的等位基因的频率将与另一端等位基因的频率相似。 当种群在地理上变得不连续时,等位基因的自由流动就会受到阻碍。 当这种分离持续一段时间时,这两个种群就能够沿着不同的轨迹进化。 因此,随着每个群体中的突变独立产生新的等位基因,它们在众多遗传基因座上的等位基因频率逐渐变得越来越不同。 通常,这两个种群的气候、资源、捕食者和竞争对手等环境条件会有所不同,导致自然选择倾向于每个群体的不同适应方式。 不同的遗传漂移史也将导致分歧,因为种群小于亲本种群,因此会得到加强。
如果有足够的时间,种群之间的遗传和表型差异可能会影响到足够影响繁殖的特征,以至于两个种群中的个体聚集在一起,交配的可能性就会降低,或者如果发生交配,后代将无法存活或不育。 许多类型的不同角色可能会影响两个种群的生殖隔离(无法杂交)。 这些生殖隔离机制可分为预合子机制(受精前起作用的机制)和合子后机制(受精后起作用的机制)。 Prezygotic 机制包括允许个体找到彼此的特征,例如交配时间、对费洛蒙的敏感性或交配地点的选择。 如果个人能够相遇,性格分歧可能会阻止求爱仪式导致交配,这要么是因为女性的偏好发生了变化,要么是男性行为发生了变化。 如果能够交配,生理变化可能会干扰成功受精。 Postzygotic 机制包括阻碍后代正常发育的遗传不相容性,或者如果后代活着,它们自己可能无法产生可行的配子,例如骡子、雌马和雄驴的不育后代。
如果将两个孤立的种群重新聚集在一起,并且由两个种群的个体交配形成的杂交后代存活率较低或生育能力降低,那么选择将有利于能够区分自己种群中潜在伴侣的个体和其他人群。 这种选择将增强生殖隔离。
分离导致同种异体物种形成的种群可以通过多种方式发生:从河流形成新的分支,侵蚀形成新的山谷,或者一群无法返回的新地点的生物,例如漂浮在海洋上空的种子到岛屿。 隔离种群所必需的地理分离的性质完全取决于生物体的生物学及其传播潜力。 如果两个飞行昆虫种群居住在附近的不同山谷中,则每个种群中的个体很可能会来回飞行,继续基因流动。 但是,如果两个啮齿动物种群因新湖的形成而分开,则不太可能持续进行基因流动;因此,物种形成的可能性更大。
生物学家将同种异体过程分为两类。 如果一个物种的几个成员迁移到新的地理区域,这称为分散。 如果出现物理分裂生物的自然情况,这就是所谓的替代现象。
科学家记录了许多同种异体物种形成的案例。 例如,在美国西海岸,存在两个单独的斑点猫头鹰亚种。 北方斑点猫头鹰与生活在南方的近亲墨西哥斑点猫头鹰有遗传和表型差异(图\(\PageIndex{2}\))。 它们最初分离的原因尚不清楚,但这可能是由冰河时代的冰川将初始种群一分为二造成的。 1
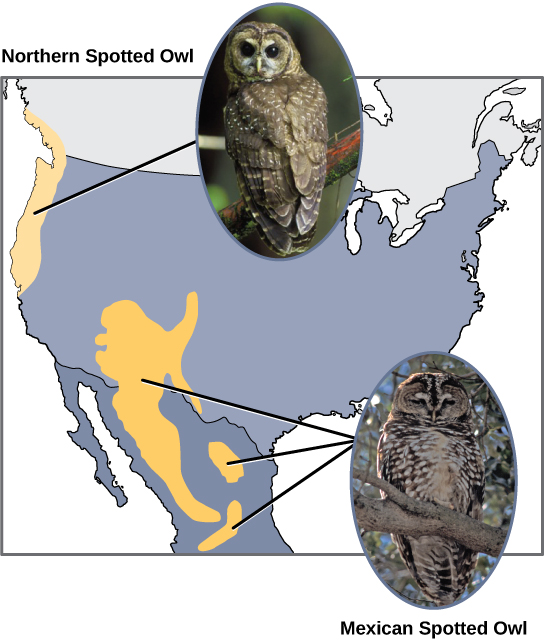
此外,科学家们发现,曾经是同一个物种的两个群体之间的距离越远,物种形成的可能性就越大。 这似乎是合乎逻辑的,因为随着距离的增加,各种环境因素的共同点可能不如近距离的地点那么共同点。 以两只猫头鹰为例;在北方,气候比南方凉爽;每个生态系统中的其他类型的生物不同,它们的行为和习惯也不同;此外,南方猫头鹰的狩猎习惯和猎物选择与北方猫头鹰不同。 这些差异可能导致猫头鹰的进化差异,随着时间的推移,除非种群之间的基因流得以恢复,否则很可能会出现物种形成。
在某些情况下,一个物种的种群分散在整个区域,每个物种都会找到不同的利基或孤立的栖息地。 随着时间的推移,他们新生活方式的不同需求导致了源自单一物种的多种物种形成事件,这就是所谓的适应性辐射。 从一个起源点开始,许多适应方法不断演变,导致该物种辐射成几个新物种。 像夏威夷群岛这样的岛屿群岛为适应性辐射事件提供了理想的环境,因为水环绕着每个岛屿,这导致许多生物在地理上处于隔离状态(图\(\PageIndex{3}\))。 夏威夷蜜爬虫举例说明了自适应辐射的一个例子。 从一个称为创始物种的单一物种中进化出许多物种,包括图中所示的八个物种\(\PageIndex{3}\)。
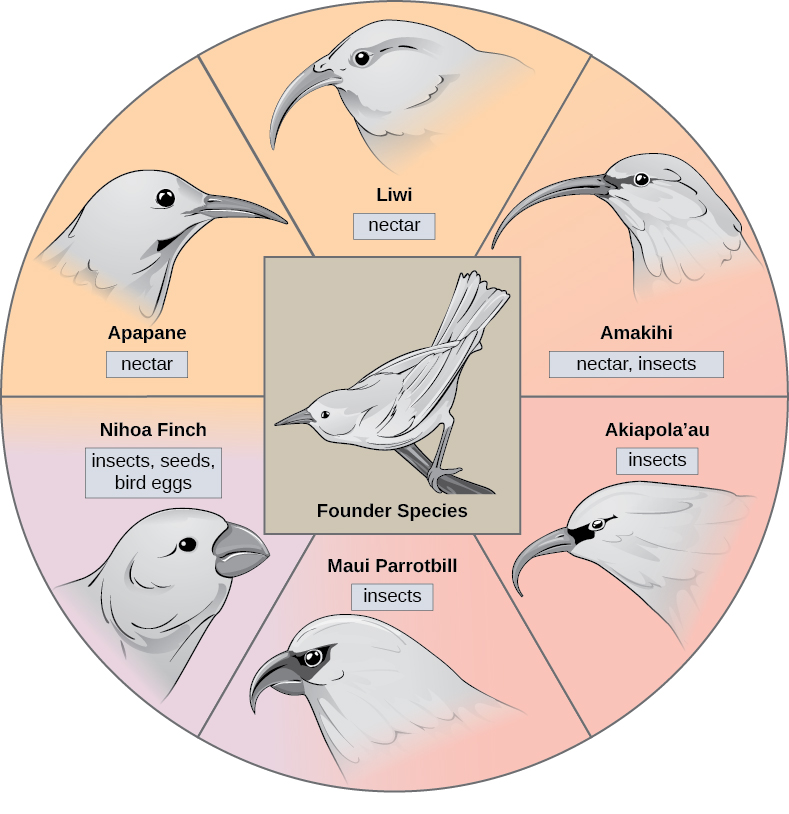
注意图中物种的喙的差异\(\PageIndex{3}\)。 根据每个新栖息地的特定食物来源进行自然选择,喙的遗传变异发生了变化,导致了适合特定食物来源的不同喙的进化。 吃水果和种子的鸟的喙更厚、更强壮,适合打破坚果。 吃花蜜的鸟有长嘴可以浸入花中才能得到花蜜。 吃昆虫的鸟有像剑一样的嘴,适合刺伤和刺穿昆虫。 达尔文的雀科是另一个经过充分研究的群岛中适应性辐射的例子。
没有地理分离的形态形成
如果没有物理屏障来隔离继续在同一栖息地生活和繁殖的个体,会发生分歧吗? 已经提出并研究了许多同情物种形成的机制。
一种形式的同情物种形成可能始于减数分裂期间的染色体错误或染色体过多的杂交个体的形成。 多倍体是一种细胞或生物体拥有一组或多组额外染色体的情况。 科学家已经确定了两种主要的多倍体类型,它们可能导致处于多倍体状态的个体的生殖隔离。 在某些情况下,多倍体个体将拥有来自自身物种的两组或更多完整的染色体,这种情况称为自多倍体(图\(\PageIndex{4}\))。 前缀 “auto” 表示自我,因此该术语表示来自自己物种的多条染色体。 多倍体是由减数分裂中的一个错误造成的,在该错误中,所有染色体都移入一个细胞,而不是分离。
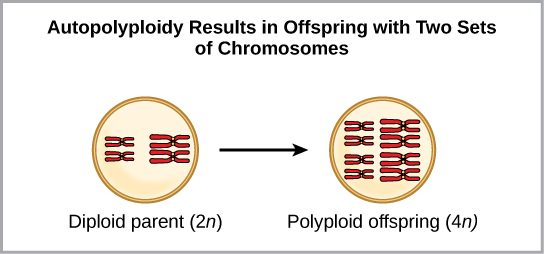
例如,如果 2 n = 6 的植物物种产生的自多倍体配子也是二倍体(2 n = 6,当它们应该为 n = 3 时),则配子现在的染色体数量是应有的两倍。 这些新配子将与该植物物种产生的普通配子不相容。 但是它们既可以自授粉,也可以与其他配子具有相同二倍体数的自多倍体植物一起繁殖。 这样,通过形成 4 n 的后代,称为四倍体,可以快速形成同情物种形态。 这些个体只能用这种新物种而不是祖先物种的个体立即繁殖。 另一种形式的多倍体发生在两个不同物种的个体繁殖形成称为同种异体多倍体的可存活后代时。 前缀 “allo” 的意思是 “其他”(从同种异体中召回);因此,当来自两个不同物种的配子结合时,就会出现同种异体多倍体。 该图\(\PageIndex{5}\)说明了同种多倍体形成的一种可能方式。 请注意,需要两代人或两代人的繁殖行为才能产生可行的肥沃杂种。
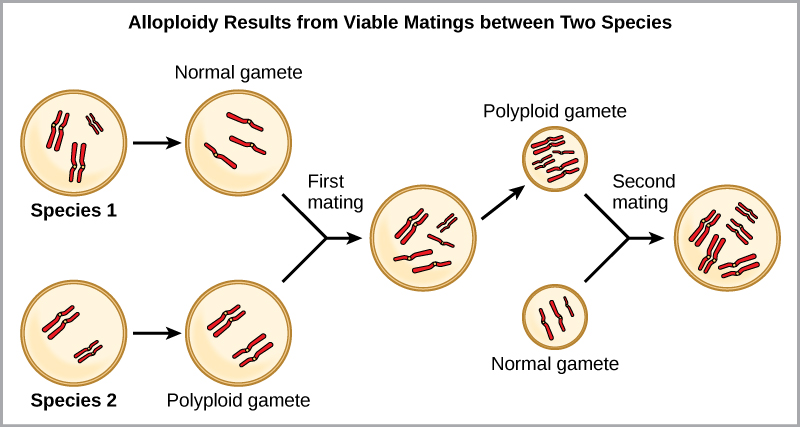
小麦、棉花和烟草植物的栽培形式都是同种异体类。 尽管多倍体偶尔发生在动物身上,但动物的大多数染色体异常是致命的;它最常见于植物中。 科学家们发现,在所有研究的植物物种中,有超过1/2与通过多倍体进化的物种有关。
Sympatric 物种形成也可能以多倍体以外的其他方式发生。 例如,想象一下生活在湖中的鱼类。 随着人口的增长,对食物的竞争也加剧。 在寻找食物的压力下,假设其中一组鱼具有遗传灵活性,可以发现并以其他鱼类未使用的另一种资源为食。 如果在湖的不同深度发现了这种新的食物来源会怎样? 随着时间的推移,以第二种食物来源为食的人之间的相互作用比其他鱼类的互动更多;因此它们也会一起繁殖。 这些鱼的后代很可能会像他们的父母一样行事,在同一个地区喂食和生活,从而使它们与原始种群分开。 如果这组鱼继续与第一个种群分开,那么随着它们之间积累更多的遗传差异,最终可能会出现同情形态的形成。
这种情况确实在自然界中发挥作用,其他导致生殖隔离的情况也是如此。 其中一个地方就是非洲的维多利亚湖,它以慈鲷鱼的同情形态而闻名。 研究人员在这些鱼类中发现了数百个同情物种形成事件,这些事件不仅大量发生,而且发生在很短的时间内。 该图\(\PageIndex{6}\)显示了尼加拉瓜慈鲷鱼种群中的这种物种形态。 在这个地方,两种类型的慈鲷生活在同一个地理位置;但是,它们的形态已经不同,可以吃不同的食物来源。
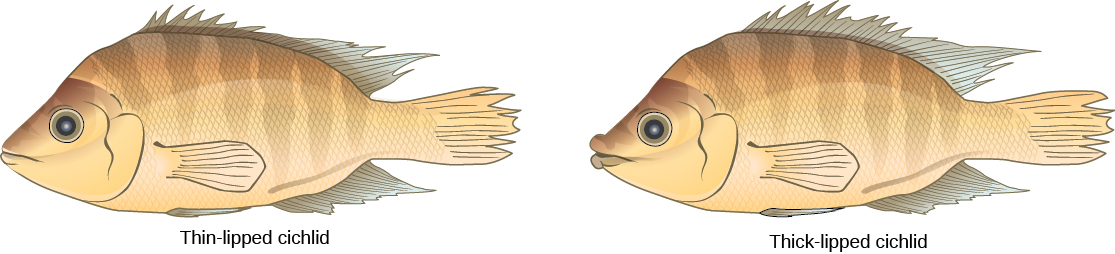
最后,一个有据可查的持续的同情物种形态形态形成的例子发生在苹果蛆 Rhagoletis pomonella 中,它是在苹果引入北美的某个时候作为孤立的种群出现的。 当地的苍蝇种群以山楂物种为食,具有宿主特异性:它只侵扰山楂树。 重要的是,它还使用树木作为交配的交配地点。 据推测,无论是由于突变还是行为错误,苍蝇都会跳过宿主,在苹果树中相遇和交配,然后在苹果果中产卵。 后代成熟并保持了对苹果树的偏爱,实际上将原始种群分为两个新的种群,这些种群由宿主物种分开,而不是按地理位置分开。 宿主跳跃发生在十九世纪,但现在两种苍蝇种群之间存在可衡量的差异。 一般而言,寄生虫的宿主特异性似乎是同情物种形成的常见原因。
章节摘要
物种形成沿着两个主要途径发生:地理分离(同种异体物种形成)和通过共享栖息地内发生的机制(同情物种形成)。 这两种途径都迫使人群之间的生殖隔离。 Sympatric 物种形成可以通过减数分裂的错误而发生,减数分裂会形成带有额外染色体的配子,称为多倍体。 Autopolyploidy 发生在单个物种中,而同种多倍体是由于密切相关的物种之间的交配而发生的。 一旦种群被隔离,就会发生进化分歧,导致生殖隔离特征的演变,如果两个种群再次聚集在一起,这些特征会阻止杂交。 经过一段时间的隔离,杂交后代的生存能力下降有望选择更强的固有隔离机制。
脚注
- 1 Courtney,S.P. 等人,“对北方斑点猫头鹰状况的科学评估”,可持续生态系统研究所(2004),俄勒冈州波特兰。
词汇表
- 自适应辐射
- 当一个物种辐射出来形成其他几个物种时的物种形成
- 同种异体形态形成
- 通过地理分离产生的形态形成
- 扩散的
- 一种同种异体物种形态,发生在物种的少数成员迁移到新的地理区域时
- 形态
- 一个新物种的形成
- 同情形态形成
- 出现在同一个地理空间中的形态形态
- 代教
- 一种同种异体物种形态,发生在环境中的某物将相同物种的生物分成不同的组时