
17.2:化学防御
- Page ID
- 200034

学习目标
- 描述体液中的酶如何提供预防感染或疾病的保护
- 列出并描述抗菌肽、补体成分、细胞因子和急性期蛋白的功能
- 描述经典、替代和凝集素补体途径之间的相似之处和不同之处
除了物理防御外,与生俱来的非特异性免疫系统还使用许多抑制微生物入侵者的化学介质。 “化学介质” 一词涵盖了在全身各种体液和组织中发现的各种物质。 化学介质可以单独起作用,也可以相互作用以抑制微生物的定植和感染。
有些化学介质是内源产生的,这意味着它们是由人体细胞产生的;另一些则是外源产生的,这意味着它们是由属于微生物组的某些微生物产生的。 有些介质是持续产生的,在抗微生物物质中沐浴该区域;其他介质主要是在应对某些刺激(例如微生物的存在)时产生或激活的。
体液中发现的化学和酶介质
皮肤产生的液体包括内源和外源介质的例子。 真皮中的皮脂腺分泌一种叫做皮脂的油,这种油通过毛囊释放到皮肤表面。 这种皮脂是一种内源性介质,通过帮助封闭毛囊的毛孔,防止皮肤表面的细菌侵入汗腺和周围组织,从而提供额外的防御层(图\(\PageIndex{1}\))。 微生物组的某些成员,例如痤疮丙酸杆菌和马拉色菌等,可以使用脂肪酶来降解皮脂,将其用作食物来源。 这会产生油酸,油酸会在皮肤表面形成温和的酸性环境,对许多致病微生物不友好。 油酸是外源产生的介质的一个例子,因为它是由常驻微生物产生的,而不是由人体细胞直接产生的。
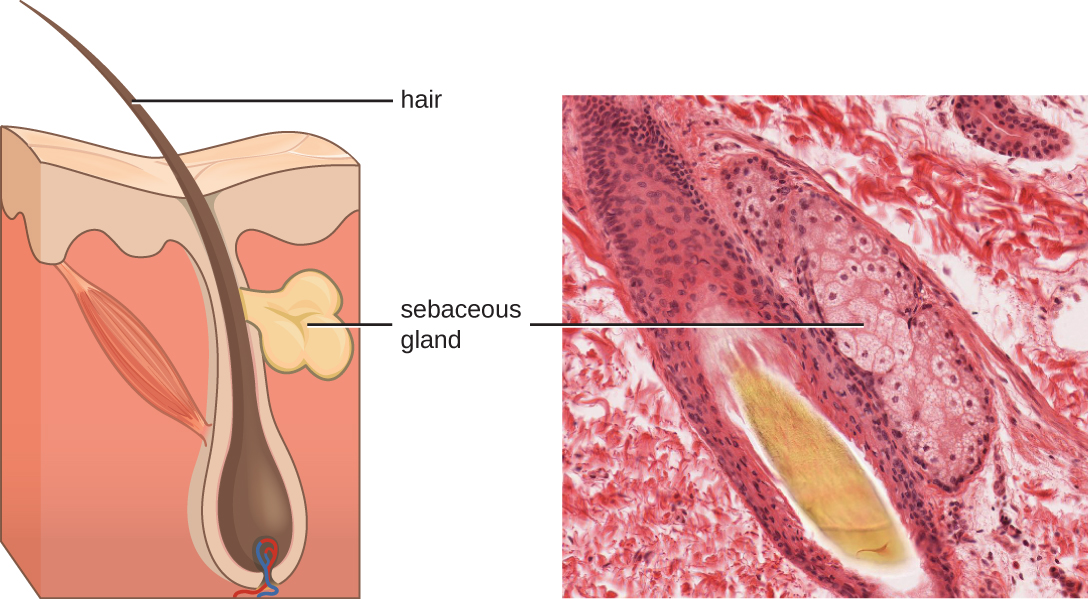
影响皮肤微生物群的环境因素会直接影响化学介质的产生。 例如,低湿度或皮脂分泌减少会使产生油酸的微生物对皮肤不宜居住,从而使皮肤更容易受到通常受皮肤低pH值抑制的病原体的影响。 许多皮肤保湿剂的配方旨在通过恢复皮肤的水分和精油来抵消这种影响。
消化道还会产生大量抑制或杀死微生物的化学介质。 在口腔中,唾液含有乳过氧化物酶等介质,食道分泌的粘液含有抗菌酶溶菌酶。 在胃中,高酸性胃液会杀死大多数微生物。 在下消化道中,肠道含有胰腺和肠道酶、抗菌肽(隐形肽)、肝脏产生的胆汁和产生溶菌酶的特殊Paneth细胞。 这些介质共同消灭了大多数能够在胃酸性环境中存活的病原体。
在尿路中,尿液会在排尿时将微生物排出体外。 此外,尿液的轻微酸度(平均 pH 值约为 6)会抑制尿路中许多微生物和潜在病原体的生长。
雌性生殖系统使用乳酸(一种外源产生的化学介质)来抑制微生物的生长。 构成阴道的细胞和组织层会产生糖原,糖原是一种分支且更复杂的葡萄糖聚合物。 该地区的乳酸杆菌发酵糖原产生乳酸,降低阴道中的pH值并抑制短暂的微生物群、念珠菌(一种与阴道感染相关的酵母菌)等机会性病原体以及其他导致性传播疾病的病原体。
在眼睛中,眼泪含有溶菌酶和乳铁蛋白的化学介质,两者都能够消灭进入眼睛表面的微生物。 Lysozyme 在 peptidoglycan 中分离 NAG 和 NAM 之间的键,肽聚糖是细菌细胞壁的成分。 它对革兰氏阳性细菌更有效,革兰氏阳性细菌缺乏与革兰氏阴性细菌相关的保护性外膜。 乳铁蛋白通过化学结合和封存铁来抑制微生物的生长。 这实际上使许多需要铁才能生长的微生物挨饿。
在耳朵中,由于存在脂肪酸,耳垢(耳垢)具有抗菌特性,脂肪酸会将pH值降至3到5之间。
呼吸道在鼻道、气管和肺部使用各种化学介质。 鼻腔产生的粘液含有与泪液和唾液中发现的抗微生物分子相似的混合物(例如溶菌酶、乳铁蛋白、乳过氧化物酶)。 气管和肺部的分泌物还含有溶菌酶和乳铁蛋白,以及各种其他化学介质,例如具有抗菌特性的称为表面活性剂的脂蛋白复合物。
练习\(\PageIndex{1}\)
- 解释内源性和外源性介质之间的区别
- 描述 pH 值如何影响抗微生物防御能力
抗菌肽
抗菌肽 (AMP) 是一类特殊的非特异性细胞衍生介质,具有广谱抗菌特性。 一些 AMP 通常由人体产生,而另一些则主要是在出现入侵病原体时产生(或大量生产)。 已经开始研究如何将 AMP 用于疾病的诊断和治疗。
AMP 可能通过多种方式诱发微生物中的细胞损伤,包括对膜造成损伤、破坏 DNA 和 RNA 或干扰细胞壁合成。 根据特定的抗微生物机制,特定的 AMP 可能仅抑制某些微生物群体(例如革兰氏阳性菌或革兰氏阴性细菌),或者它可能对细菌、真菌、原生动物和病毒更广泛有效。 在皮肤上发现了许多 AMP,但也可以在身体的其他部位找到。
一种叫做防御素的 AMP 家族可以由全身的上皮细胞产生,也可以由巨噬细胞和中性粒细胞等细胞防御产生(参见 Cell ul ar Defensins)。 防御素可能在宿主细胞内分泌或起作用;它们通过破坏质膜来对抗微生物。 被称为 bacteriocins 的 AMP 是由胃肠道内某些常驻微生物群的成员通过外源产生的。 编码这些类型AMP的基因通常携带在质粒上,并且可以通过横向或水平基因转移在常驻微生物群中的不同物种之间传递。
全身还有许多其他放大器。 表中总结了几个比较重要的 AMP 的特征\(\PageIndex{1}\)。
| 放大器 | 秘密者 | 身体部位 | 病原体受到抑制 | 行动模式 |
|---|---|---|---|---|
| Bacteriocins | 常住微生物群 | 胃肠道 | 细菌 | 破坏膜 |
| Cathelicidin | 上皮细胞、巨噬细胞和其他细胞类型 | 皮肤 | 细菌和真菌 | 破坏膜 |
| 防御 | 上皮细胞、巨噬细胞、中性粒细胞 | 贯穿全身 | 真菌、细菌和许多病毒 | 破坏膜 |
| Dermicidin | 汗腺 | 皮肤 | 细菌和真菌 | 破坏膜完整性和离子通道 |
| 组胺类药物 | 唾液腺 | 口腔 | 真菌 | 破坏细胞内功能 |
练习\(\PageIndex{2}\)
为什么抗微生物肽 (AMP) 被视为非特异性防御?
血浆蛋白介质
在血浆(血液的液体部分)中发现了许多非特异性先天免疫因子。 血浆含有电解质、糖、脂质和蛋白质,每种都有助于维持稳态(即稳定的体内功能),并且含有参与血液凝结的蛋白质。 血浆中发现的其他蛋白质,例如急性期蛋白、补体蛋白和细胞因子,参与非特异性先天免疫反应。
血浆与血清
血液的液体部分有两个术语:血浆和血清。 如果它们既是液体又缺少细胞,它们有何不同? 凝血(血细胞凝结)发生后剩下的血液部分是血清。 尽管血清中仍然存在许多维生素、电解质、某些糖、补体蛋白和抗体等分子,但凝血因子已基本耗尽。 相反,血浆仍然含有所有的凝血元素。 要从血液中获得血浆,必须使用抗凝剂来防止凝血。 抗凝剂的例子包括肝素和乙二胺四乙酸(EDTA)。 由于凝血受到抑制,样品一旦获得,就必须在离心机中轻轻地向下旋转。 较重、更密集的血细胞在离心管底部形成颗粒,而较轻、密度较低的液体血浆部分则保留在细胞颗粒上方。
急性期蛋白
急性期蛋白质是另一类抗微生物介质。 急性期蛋白主要在肝脏中产生,并因免疫系统中的炎症分子而分泌到血液中。 急性期蛋白的例子包括 C 反应蛋白、血清淀粉样蛋白 A、铁蛋白、转铁蛋白、纤维蛋白原和甘露糖结合凝集素。 这些蛋白质中的每一种都有不同的化学结构,并以某种方式抑制或破坏微生物(表\(\PageIndex{1}\))。
| 一些急性期蛋白及其功能 | |
|---|---|
| C 反应蛋白 | Coats 细菌(opsonization),为吞噬细胞摄入做好准备 |
| 血清淀粉样蛋白 A | |
| 铁蛋白 | 结合并封存铁,从而抑制病原体的生长 |
| 转铁蛋白 | |
| 纤维蛋白原 | 参与形成捕获细菌病原体的血块 |
| 甘露糖结合凝集素 | 激活补充级联 |
补体系统
补体系统是一组血浆蛋白介质,可以起到与生俱来的非特异性防御作用,同时还可以连接先天免疫和适应性免疫(将在下一章中讨论)。 补体系统由30多种蛋白质(包括C1至C9)组成,这些蛋白质通常作为前体蛋白在血液中循环。 这些前体蛋白在受到多种因素(包括微生物的存在)的刺激或触发时会被激活。 补体蛋白被认为是先天非特异性免疫的一部分,因为它们始终存在于血液和组织液中,可以快速被激活。 此外,当通过替代途径(将在本节稍后介绍)激活时,补体蛋白会以非特异性方式靶向病原体。
循环补体前体发挥作用的过程称为补体激活。 这个过程是一个级联,可以由三种不同的机制之一触发,即替代途径、经典途径和凝集素途径。
替代途径由补体蛋白 C3 的自发激活启动。 C3 的水解产生两种产物,即 C3a 和 C3b。 当不存在入侵微生物时,使用血液中的水进行水解反应时,C3b 会很快降解。 但是,如果存在入侵微生物,C3b 会附着在这些微生物的表面。 一旦附着,C3b 将以级联形式招募其他补体蛋白(图\(\PageIndex{2}\))。
传统途径提供了一种更有效的激活补体级联的机制,但它取决于特定的适应性免疫防御系统产生的抗体。 要启动经典途径,特异性抗体必须首先与病原体结合以形成抗体-抗原复合物。 这会激活补体级联中的第一个蛋白质,即 C1 复合物。 C1 复合物是一种多部分蛋白质复合物,每个成分都参与整个复合物的完全激活。 在招募和激活 C1 复合物之后,剩余的经典途径补体蛋白以级联序列被招募和激活(图\(\PageIndex{2}\))。
凝集素激活途径与传统途径类似,但它是由甘露糖结合凝集素(一种急性期蛋白)与微生物表面碳水化合物的结合触发的。 像其他急性期蛋白一样,凝集素是由肝细胞产生的,通常在感染期间人体接收到的炎症信号而上调(图\(\PageIndex{2}\))。
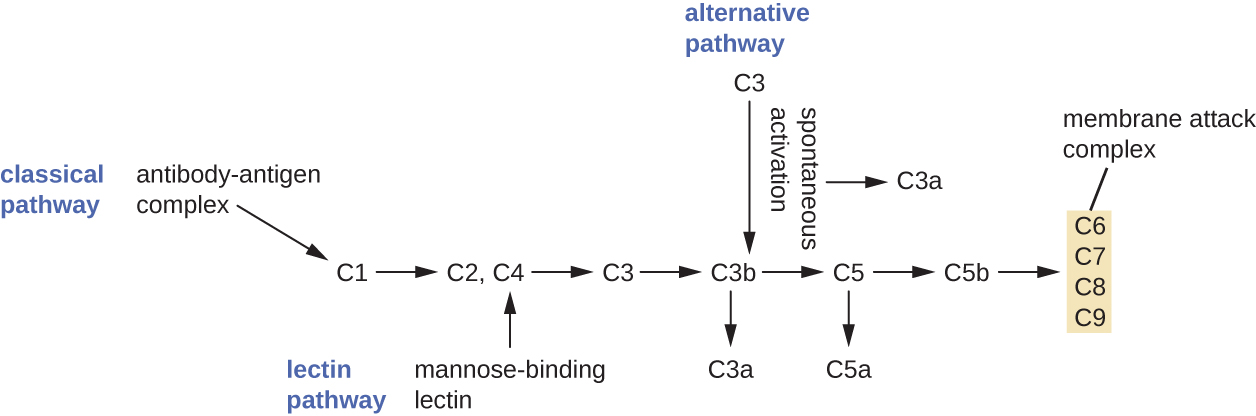
尽管每种补体激活途径的启动方式不同,但它们都提供相同的保护结果:opsonization、炎症、趋化性和细胞溶解。 opsonization 一词是指用一种化学物质(称为 opsonin)涂覆病原体,这种物质使吞噬细胞能够更容易地识别、吞没和摧毁病原体。 来自补码级联的 Opsonins 包括 C1q、C3b 和 C4b。 其他重要的 opsonins 包括甘露糖结合蛋白和抗体。 补体片段 C3a 和 C5a 是特征良好的过敏毒素,具有强大的促炎作用。 过敏毒素激活肥大细胞,导致脱颗粒和炎症化学信号的释放,包括导致血管舒张和血管通透性增加的介质。 C5a 也是中性粒细胞和其他白细胞最有效的趋化剂之一,下一节将讨论细胞防御能力。
补体蛋白 C6、C7、C8 和 C9 组合成膜攻击复合物 (MAC),允许 C9 聚合成革兰氏阴性细菌膜中的孔隙。 这些孔隙允许水、离子和其他分子自由进出靶细胞,最终导致细胞裂解和病原体死亡(图\(\PageIndex{2}\))。 但是,MAC 仅对革兰氏阴性细菌有效;它无法穿透与革兰氏阳性细菌细胞壁相关的厚厚肽聚糖层。 由于 MAC 不会对革兰氏阳性细菌病原体构成致命威胁,因此补体介导的 opsonization 对于清除革兰氏阳性细菌病原体更为重要。
细胞因子
细胞因子是可溶性蛋白,充当细胞之间的通信信号。 在非特异性先天免疫反应中,可能会释放各种细胞因子以刺激化学介质或其他细胞功能的产生,例如细胞增殖、细胞分化、抑制细胞分裂、凋亡和趋化性。
当细胞因子与其靶受体结合时,其作用可能会有很大差异,具体取决于细胞因子的类型以及它所结合的细胞或受体的类型。 特定细胞因子的功能可以描述为自分泌、旁分泌或内分泌(图\(\PageIndex{3}\))。 在自分泌功能中,释放细胞因子的同一个细胞是信号的接收者;换句话说,自分泌功能是细胞自我刺激的一种形式。 相比之下,旁分泌功能涉及将细胞因子从一个细胞释放到附近的其他细胞,从而刺激受体细胞的某些反应。 最后,当细胞将细胞因子释放到血液中并携带到更远的靶细胞时,就会产生内分泌功能。
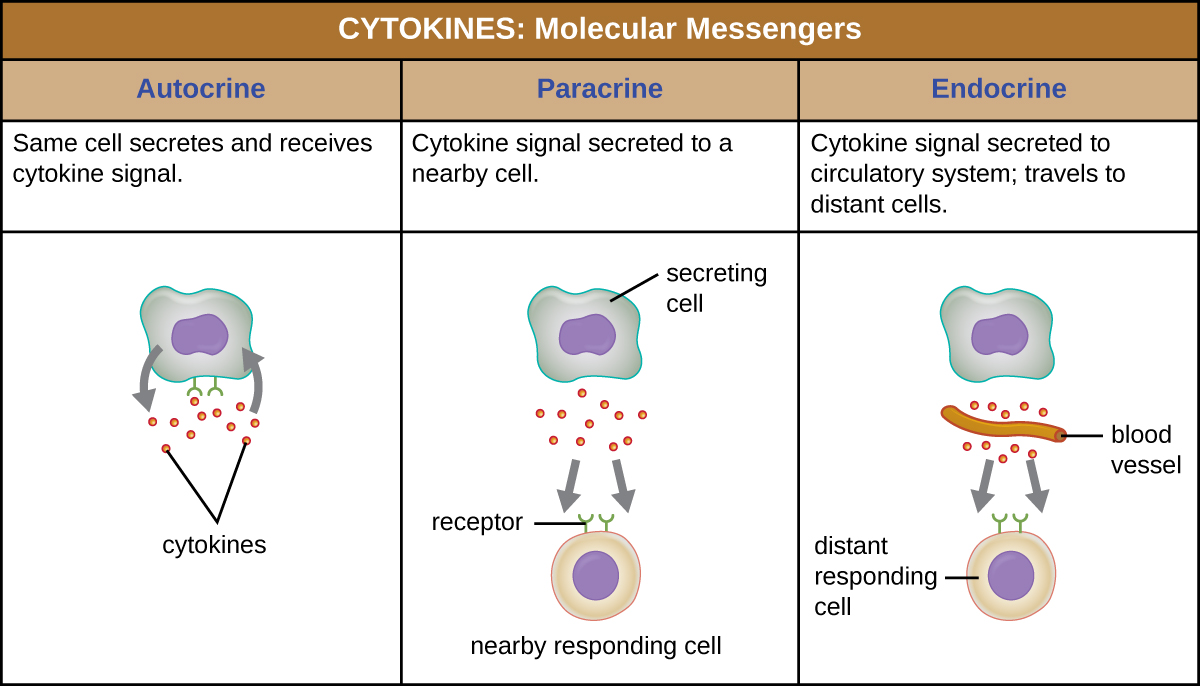
三类重要的细胞因子是白介素、趋化因子和干扰素。 白细胞介素最初被认为只能由白细胞(白细胞)产生,只能刺激白细胞,因此它们得名的原因。 尽管白介素参与调节免疫系统的几乎所有功能,但它们在体内的作用不仅限于免疫力。 白介素也由与免疫防御无关的各种细胞产生并刺激它们。
趋化因子是趋化因子,可将白细胞招募到感染、组织损伤和炎症部位。 与更常见的趋化因子(如补体因子 C5a)相反,趋化因子在它们招募的白细胞亚群中非常特异。
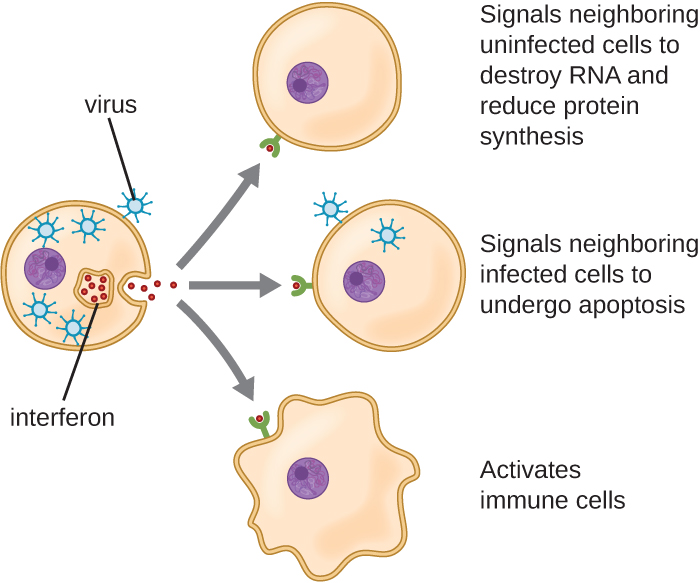
干扰素是一组多样化的免疫信号分子,对我们抵御病毒尤其重要。 I 型干扰素(干扰素 α 和 β 干扰素)由感染病毒的细胞产生和释放。 这些干扰素刺激附近的细胞停止产生 mRNA,破坏已经产生的RNA,并减少蛋白质合成。 这些细胞变化抑制了病毒的复制和成熟病毒的产生,从而减缓了病毒的传播。 I 型干扰素还刺激参与病毒清除的各种免疫细胞,从而更积极地攻击受病毒感染的细胞。 II 型干扰素(干扰素 γ)是免疫细胞的重要激活剂(图\(\PageIndex{4}\))。
引发炎症的介质
本节中讨论的许多化学介质在某种程度上会导致炎症和发热,炎症和发烧是非特异性免疫反应,详见炎症和发热。 细胞因子刺激肝脏中急性期蛋白质的产生,例如C反应蛋白和甘露糖结合凝集素。 这些急性期蛋白充当 opsonins,通过凝集素途径激活补体级联。
一些细胞因子还会结合肥大细胞和嗜碱性粒细胞,诱导它们释放组胺,一种促炎化合物。 组胺受体存在于各种细胞中,介导促炎事件,例如支气管收缩(气道收缩)和平滑肌收缩。
除组胺外,肥大细胞还可能释放其他化学介质,例如白三烯。 白三烯是基于脂质的促炎介质,由白细胞和组织细胞细胞膜中花生四烯酸的代谢产生。 与组胺的促炎作用相比,白三烯的促炎作用更有效,持续时间更长。 这些化学介质一起可以诱发咳嗽、呕吐和腹泻,从而将病原体排出体外。
某些细胞因子还刺激前列腺素的产生,前列腺素是促进激肽和组胺炎症作用的化学介质。 前列腺素还可以帮助提高体温,从而导致发烧,从而促进白细胞的活性并轻微抑制致病微生物的生长(参见炎症和发热)。
另一种炎症介质 bradykinin 会导致水肿,当液体和白细胞从血液中泄漏到组织中时,就会发生水肿。 它与毛细血管壁中细胞上的受体结合,导致毛细血管扩张并变得更容易渗透液体。
练习\(\PageIndex{3}\)
- 这三种补体激活途径有什么共同点?
- 解释自分泌、旁分泌和内分泌信号。
- 列举两个重要的引起炎症的介质。
临床重点:第 2 部分
为了缓解呼吸道收缩,安吉拉立即接受抗组胺药治疗,通过吸入器服用皮质类固醇,然后进行一段时间的监测。 尽管她的病情没有恶化,但药物似乎并不能缓解她的病情。 她被送进医院接受进一步观察、检测和治疗。
入院后,临床医生进行过敏测试,试图确定其环境中是否有东西可能触发过敏性炎症反应。 医生下令进行血液分析以检查特定细胞因子的水平。 还会采集痰液样本并将其送往实验室进行微生物染色、培养和鉴定可能导致感染的病原体。
练习\(\PageIndex{4}\)
- 先天免疫系统的哪些方面可能导致安吉拉的气道收缩?
- 为什么安吉拉用抗组胺药治疗?
- 为什么医生会对安吉拉血液中的细胞因子水平感兴趣?
\(\PageIndex{3}\)该表概述了本节中讨论的化学防御措施。
| 防御 | 示例 | 函数 |
|---|---|---|
| 体液中的化学物质和酶 | 来自皮脂腺的皮脂 | 提供油屏障,保护毛囊毛孔免受病原体侵害 |
| 来自皮脂和皮肤微生物群的油酸 | 降低 pH 值以抑制病原体 | |
| 分泌物中的溶菌酶 | 通过攻击细胞壁杀死细菌 | |
| 胃、尿和阴道中的酸 | 抑制或杀死细菌 | |
| 消化酶和胆汁 | 杀死细菌 | |
| 乳铁蛋白和转铁蛋白 | 结合并封存铁,抑制细菌生长 | |
| 肺部表面活性剂 | 杀死细菌 | |
| 抗菌肽 | 防御素、细菌素、dermicidin、cathelicidin、组织抑制素、 | 通过攻击膜或干扰细胞功能来杀死细菌 |
| 血浆蛋白介质 | 急性期蛋白(C 反应蛋白、血清淀粉样蛋白 A、铁蛋白、纤维蛋白原、转铁蛋白和甘露糖结合凝集素) | 抑制细菌的生长,帮助捕获和杀死细菌 |
| 补充 c3b 和 c4b | 对病原体进行消化以帮助吞噬作用 | |
| 补充 c5a | 吞噬细胞的趋化剂 | |
| 补充 c3a 和 c5a | 促炎过敏毒素 | |
| 细胞因子 | 白介素 | 刺激和调节免疫系统的大多数功能 |
| 趋化因子 | 将白细胞招募到感染区域 | |
| 干扰素 | 提醒细胞注意病毒感染,诱导病毒感染细胞凋亡,在受感染和附近未感染的细胞中诱导抗病毒防御,刺激免疫细胞攻击病毒感染细胞 | |
| 引发炎症的介质 | 组胺 | 促进血管舒张、支气管收缩、平滑肌收缩、增加分泌和粘液产生 |
| Leukotrienes | 促进炎症;比组胺更强且更持久 | |
| 前列腺素 | 促进炎症和发烧 | |
| Bradykinin | 增加血管舒张和血管通透性,导致水肿 |
关键概念和摘要
- 许多内源和外源产生的化学介质表现出非特异性抗微生物功能。
- 许多化学介质存在于体液中,例如皮脂、唾液、粘液、胃液和肠液、尿液、眼泪、耳垢和阴道分泌物。
- 存在于皮肤和身体其他部位的@@ 抗菌肽(AMP)主要是为了应对病原体的存在而产生的。 这些包括 dermcidin、cathelicidin、防御素、组抑素和细菌素。
- 血浆含有各种用作化学介质的蛋白质,包括急性期蛋白、补体蛋白和细胞因子。
- 补体系统涉及许多在血浆中循环的前体蛋白。 这些蛋白质在存在微生物的情况下以级联序列被激活,从而通过形成膜攻击复合体(MAC),导致病原体的消化、白细胞的化学吸引、炎症的诱导和细胞溶解。
- 细胞因子是促进先天免疫细胞的各种非特异性反应的蛋白质,包括其他化学介质的产生、细胞增殖、细胞死亡和分化。
- 细胞因子在炎症反应中起着关键作用,引发炎症的介质的产生,例如急性期蛋白、组胺、白三烯、前列腺素和缓激素。