
11.6:无性原核生物如何实现遗传多样性
- Page ID
- 200141

学习目标
- 比较转换、转导和共轭的过程
- 解释无性基因转移是如何产生原核生物遗传多样性的
- 解释转座子的结构和对细菌遗传多样性的影响
通常,当我们考虑遗传转移时,我们会想到垂直基因转移,即遗传信息代代相传。 到目前为止,垂直基因转移是所有细胞中遗传信息的主要传递方式。 在有性繁殖的生物体中,减数分裂期间的交叉事件和单个染色体的独立分类有助于种群的遗传多样性。 遗传多样性也是在有性生殖过程中引入的,当来自两个父母的遗传信息(每个父母都有不同的遗传信息补充)结合在一起时,在二倍体后代中产生了亲本基因型的新组合。 突变的发生也有助于人群的遗传多样性。 后代的遗传多样性在不断变化或不一致的环境中很有用,可能是有性生殖进化成功的原因之一。
当原核生物和真核生物无性繁殖时,它们通过垂直基因转移将几乎相同的遗传物质拷贝转移给后代。 尽管无性繁殖可以更快地产生更多的后代,但这些后代多样性的任何好处都消失了。 那么,以无性为主的繁殖模式的生物如何创造遗传多样性呢? 在原核生物中,水平基因转移(HGT)是引入遗传多样性的重要途径,即在同一代内将遗传物质从一个生物体引入另一个生物体。 HGT 甚至允许远距离相关的物种共享基因,从而影响其表型。 人们认为 HGT 在原核生物中更为普遍,但在任何时候,只有一小部分的原核生物基因组可以通过这种转移进行转移。 随着对这种现象的调查更加彻底,可以发现这种现象更加普遍。 许多科学家认为,HGT 和突变是原核生物遗传变异的重要来源,遗传变异是自然选择过程的原材料。 尽管 HGT 在与进化相关的生物中更为常见,但它可能发生在自然群落中共同生活的任何两个物种之间。
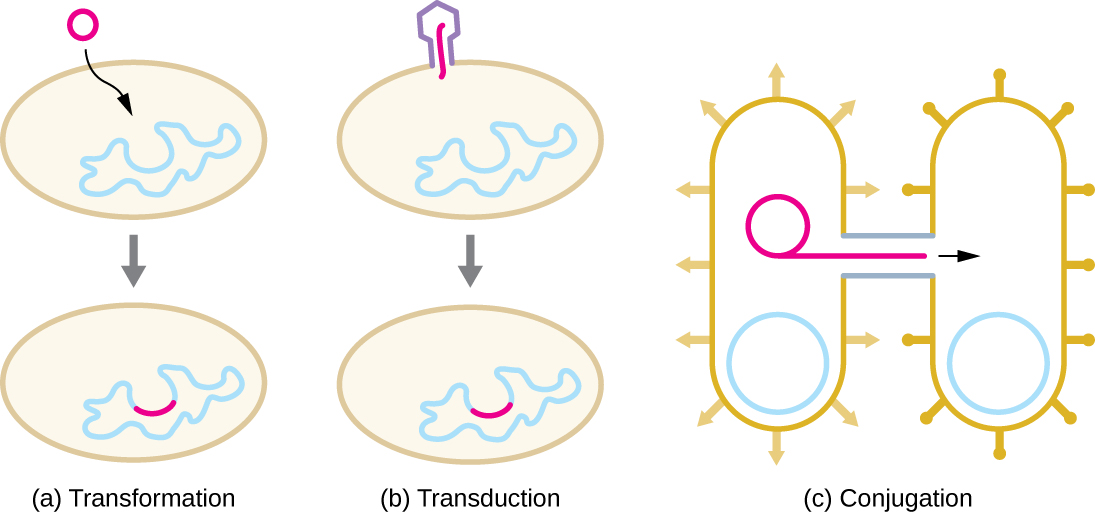
众所周知,原核生物中的 HGT 是由三种主要机制发生的,如图所示\(\PageIndex{1}\):
- 转化:裸露的 DNA 从环境中吸收
- 转导:基因在病毒的细胞之间转移(参见《病毒生命周期》)
- 共轭:使用称为共轭 pilus 的空心管在细胞之间转移基因
练习\(\PageIndex{1}\)
- 有性生殖将遗传变异引入后代的三种方式是什么?
- 无性繁殖有什么好处?
- 原核生物中水平基因转移的三种机制是什么?
转型
弗雷德里克·格里菲斯是第一个演示转型过程的人。 1928年,他表明,活的、非致病性的肺炎链球菌细菌可以通过暴露于热杀死的致病菌株转化为致病细菌。 他总结说,某种病原体,他称之为 “转化原理”,已经从死亡的致病细菌传递给活的非致病细菌。 1944 年,奥斯瓦尔德·艾弗里(1877—1955 年)、科林·麦克劳德(1909—1972 年)和麦克琳·麦卡蒂(1911—2005 年)证明转化原理是 DNA(参见 “利用微生物学发现生命的秘密”)。
在转化过程中,原核生物吸收了在其环境中发现的裸DNA,这些DNA来自其他细胞,这些细胞在死亡时会裂解并将其内容物(包括基因组)释放到环境中。 许多细菌具有天生能力,这意味着它们会主动与环境DNA结合,将其通过细胞包膜运送到细胞质中,然后使其成为单链。 通常,细胞内的双链外来DNA会被核酸酶破坏,以防病毒感染。 但是,这些核酸酶通常对单链 DNA 无效,因此细胞内的这种单链 DNA 有机会重组到细菌基因组中。 含有来自不同生物的 DNA 片段的 DNA 分子称为重组 DNA。 (重组DNA将在《微生物和基因工程工具》中进行更详细的讨论。) 如果细菌通过重组将新的 DNA 纳入自己的基因组,则细菌细胞可能会获得新的表型特性。 例如,如果非致病性细菌从病原体中吸收毒素基因的DNA,然后将其纳入染色体,则它也可能具有致病性。 质粒 DNA 也可能被合格细菌吸收,赋予细胞新的特性。 总体而言,自然界中的转化是一个相对低效的过程,因为环境中的DNA水平很低,这是因为细胞裂解过程中也会释放出来的核酸酶的活性。 此外,基因重组在将新的 DNA 序列纳入基因组方面效率低下。
在自然界中,细菌转化是获取编码毒力因子和抗生素耐药性的遗传元素的重要机制。 事实证明,编码抗微生物化合物耐药性的基因在自然界中很普遍,即使在不受人类影响的环境中也是如此。 这些基因允许生活在混合群落中的微生物争夺有限的资源,它们可以通过转化以及HGT的其他过程在种群中转移。 在实验室中,我们可以利用细菌转化的自然过程进行基因工程,制造出各种各样的药品,正如《微生物与基因工程工具》中所讨论的那样。
练习\(\PageIndex{2}\)
为什么细菌细胞会将带入细胞的环境 DNA 变成单链形式?
转导
感染细菌的病毒(噬菌体)也可能通过一种称为转导的过程将一小段染色体DNA从一种细菌转移到另一种细菌(见图6.2.3)。 回想一下,在广义转导中,任何染色体DNA片段都可以通过在噬菌体组装过程中意外将染色体DNA封装到噬菌体头部来转移到新的宿主细胞。 相比之下,专门的转导是由于不精确地从细菌染色体上切除溶酶原预言,从而将其从噬菌体整合位点两侧携带到新的宿主细胞。 因此,房东可能会获得新的财产。 这个过程被称为溶原转化。 具有医学意义的是,溶血性噬菌体可能携带毒力基因到其新宿主。 一旦插入到新宿主的染色体中,新宿主可能会获得致病性。 几种致病细菌,包括白喉棒状杆菌(白喉的病原体)和肉毒梭菌(肉毒中毒的病原体),都具有毒性,因为溶解性噬菌体引入了毒素编码基因,这证实了临床转导与传染病有关的基因交换中的相关性。 古细菌有自己的病毒,可以将遗传物质从一个人转移到另一个个体。
练习\(\PageIndex{3}\)
- 原核细胞转导的因子是什么?
- 在专业转导中,DNA的转导片段来自哪里?
转导的临床后果
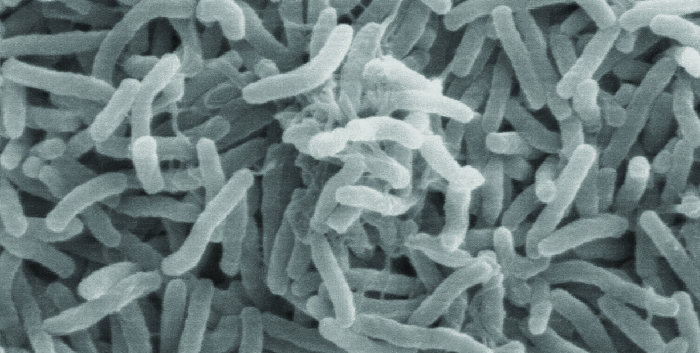
保罗是来自亚特兰大的23岁救济工作者,在2010年地震后于2011年前往海地提供援助。 在那里工作了几个星期后,他突然开始出现腹部不适,包括严重的抽筋、恶心、呕吐和水样腹泻。 他还开始经历剧烈的肌肉抽筋。 在当地一家诊所,医生怀疑保罗的症状是由霍乱引起的,因为地震后爆发了霍乱。 由于霍乱是通过粪口途径传播的,因此卫生基础设施的破坏(通常发生在自然灾害之后)可能会引发疫情。 医生使用霍乱量油尺测试证实了推定诊断。 然后,他给保罗开了单剂量的强力霉素和口服补液盐,指示他喝大量的干净水。
霍乱是由革兰氏阴性曲杆霍乱弧菌引起的(图\(\PageIndex{2}\))。 其症状主要源于霍乱毒素(CT)的产生,该毒素最终激活氯化物转运蛋白,将氯离子从上皮细胞中泵入肠腔。 然后,水会跟随氯离子,导致霍乱所特有的多产水样腹泻。 编码霍乱毒素的基因被整合到 V 的细菌染色体中。 霍乱是通过使用溶原丝状CTX噬菌体感染细菌引起的,该噬菌体携带CT基因,并在预言整合后将其引入染色体。 因此,霍乱杆菌的致病菌株是由专门转导的水平基因转移产生的。
练习\(\PageIndex{4}\)
- 为什么自然灾害导致霍乱疫情更常见?
- 为什么肌肉抽筋是霍乱的常见症状? 为什么口服补液盐治疗对霍乱的治疗如此重要?
- 在受霍乱影响的地区,人们可以使用哪些策略来预防疾病传播?
共轭
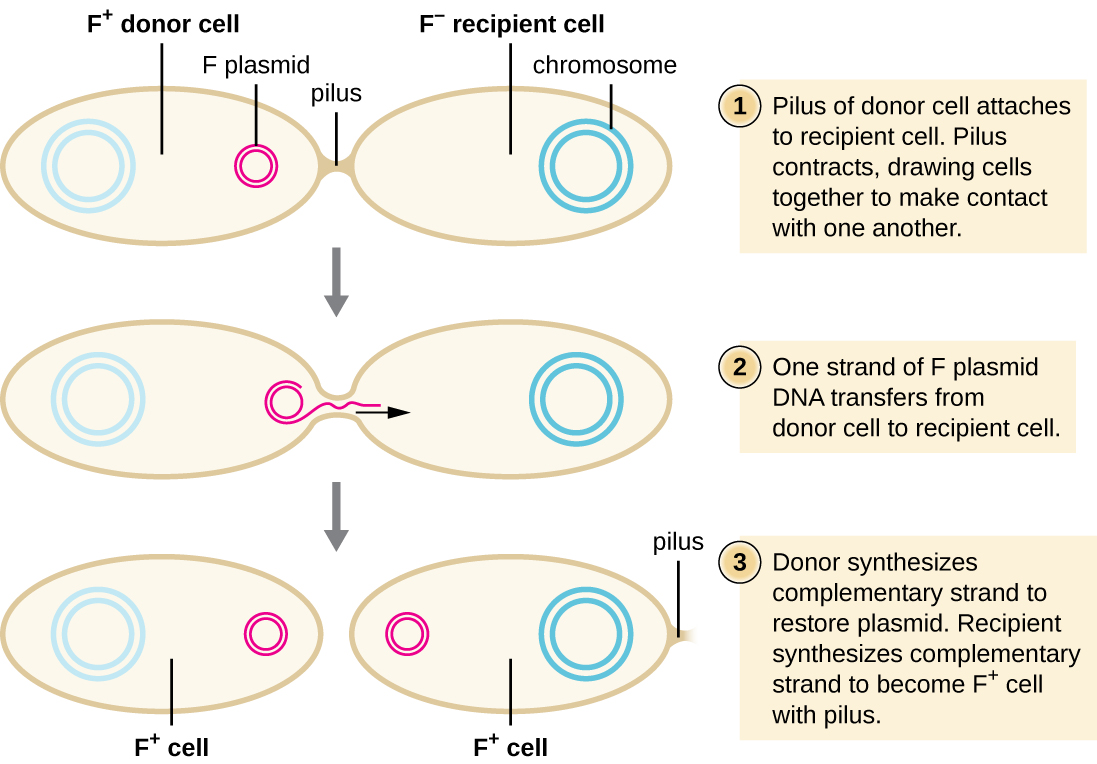
在偶联中,DNA通过偶联pilus直接从一种原核生物转移到另一种原核生物,这使生物相互接触。 在大肠杆菌中,编码偶联能力的基因位于一种叫做 F 质粒(也称为生育因子)的细菌质粒上,而共轭 pilus 被称为 F pilus。 F-plasmid 基因既编码构成 F pilus 的蛋白质,也编码参与质粒滚动循环复制的蛋白质。 含有能够形成 F pilus 的 F 质粒的细胞被称为 F + 细胞或供体细胞,缺少 F 质粒的细胞被称为 F − 细胞或受体细胞 s。
F 质粒的共轭
在大肠杆菌的典型偶联过程中,F + 细胞的 F pilus 与 F — 细胞接触并缩回,使两个细胞包膜接触(图\(\PageIndex{3}\))。 然后,在共轭 pilus 部位的两个细胞之间形成细胞质桥。 当 F 质粒在 F + 细胞中发生滚动环复制时,F 质粒的单链拷贝通过细胞质桥转移到 F − 细胞,然后由后者合成互补链,使其成为双链。 F − 细胞现在变成了能够产生自己的共轭 pilus 的 F + 细胞。 最终,在同时包含 F + 和 F − 细胞的混合细菌群中,所有细胞都将变成 F + 细胞。 大肠杆菌 F 质粒上的基因还编码阻止 F + 细胞之间偶联的蛋白质。
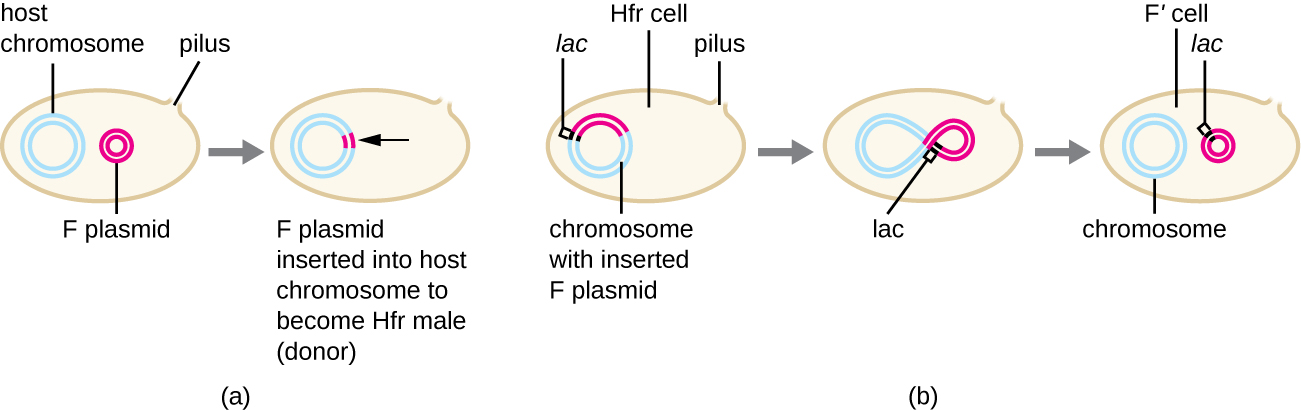
F'和Hfr细胞的偶联
尽管大肠杆菌中的典型偶联仅导致 F 质粒 DNA 的转移,但偶联也可能转移染色体 DNA。 这是因为 F 质粒偶尔会通过质粒和染色体之间的重组整合到细菌染色体中,形成 Hfr 细胞(图\(\PageIndex{4}\))。 “Hfr” 是指受体 F − 细胞通过偶联从 Hfr 细胞接收遗传信息时出现的高重组频率。 与在专业转导过程中不精确地切除预言者类似,整合的 F 质粒也可能被不精确地从染色体上切除,从而产生一个 F' 质粒,该质粒携带一些与整合位点相邻的染色体 DNA。 偶联后,这种 DNA 被引入受体细胞,既可以作为 F' 质粒的一部分维持,也可以重组到受体细胞的细菌染色体中。
Hfr 细胞还可以像对待巨大的 F 质粒一样对待细菌染色体,并尝试将其副本转移给受体 F 细胞。 由于细菌染色体非常大,整个染色体的转移需要很长时间(图\(\PageIndex{5}\))。 但是,在偶联过程中,细菌细胞之间的接触是短暂的,因此整个染色体被转移的情况并不常见。 与更远的宿主基因相比,F质粒整合位点附近的宿主染色体DNA被滚动圈复制的单向过程所取代,更有可能被转移和重组到受体细胞的染色体中。 因此,可以根据细菌基因何时通过偶联转移来绘制细菌基因在Hfr细胞基因组中的相对位置。 因此,在广泛进行细菌基因组测序的时代之前,原核生物基因组图谱上的距离通常是在几分钟内测量的。
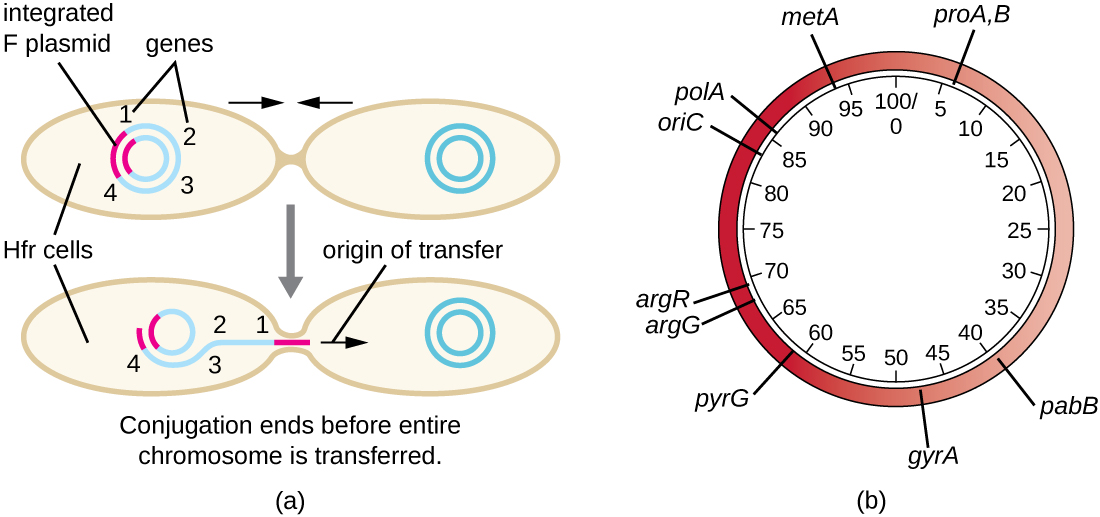
共轭的后果和应用
质粒是细菌中一种重要的染色体外DNA元素,在含有质粒的细胞中,质粒被认为是细菌基因组的一部分。 从临床角度来看,质粒通常编码与毒力有关的基因。 例如,编码使细菌细胞对特定抗生素产生抗药性的蛋白质的基因编码在 R 质粒上。 R 质粒除了具有抗微生物药物耐药性的基因外,还含有控制质粒偶联和转移的基因。 R 质粒能够在相同物种的细胞之间和不同物种的细胞之间转移。 单 R 质粒通常包含多个基因,可对多种抗生素产生耐药性。
还可以在质粒上发现产生对感染期间定植至关重要的各种毒素和分子所需的基因。 例如,产维罗毒素的大肠杆菌(VTEC)菌株似乎是通过获取编码该毒素的大型质粒从革兰氏阴性亲属志贺氏痢疾志贺氏菌那里获得了编码志贺毒素的基因。 VTEC 会导致严重的腹泻疾病,可能导致溶血尿毒症综合征 (HUS),从而可能导致肾衰竭和死亡。
在非临床环境中,编码降解多环芳烃(PAH)等特殊非典型化合物所需的代谢酶的细菌基因也经常编码在质粒上。 此外,某些质粒能够通过与偶联不同的机制从细菌细胞转移到其他细胞类型,例如动植物细胞。 微生物遗传学的现代应用涵盖了此类机制及其在基因工程中的应用。
练习\(\PageIndex{5}\)
- 共轭期间会发生什么类型的复制?
- 产生 Hfr 大肠杆菌细胞会发生什么?
- 质粒上编码了哪些类型的性状?
换位
称为转座子(转座元素)或 “跳跃基因” 的遗传元素是DNA分子,其末端包括特殊的反向重复序列和编码转座酶的基因(图\(\PageIndex{6}\))。 转座子允许整个序列从 DNA 分子中的一个位置独立切除,然后通过一种称为移位的过程整合到其他地方的 DNA 中。 转座子最初是由美国遗传学家芭芭拉·麦克林托克(1902—1992 年)在 20 世纪 40 年代在玉米(玉米)中发现的。 此后,在所有类型的生物中都发现了转座子,包括原核生物和真核生物。 因此,与前面讨论的三种机制不同,移位不是原核生物特异性的。 大多数转座子是不可复制的,这意味着它们以 “剪切粘贴” 的方式移动。 但是,有些可能是复制的,在制作副本以插入其他地方(“复制粘贴”)的同时,保留了它们在DNA中的位置。 因为转座子可以在 DNA 分子内移动,从一个 DNA 分子移动到另一个 DNA 分子,甚至从一个细胞移到另一个细胞,所以它们有能力引入遗传多样性。 同一 DNA 分子内的运动可以通过使基因失活或激活来改变表型。
转座子可能携带额外的基因,从而将这些基因从一个位置转移到另一个位置。 例如,细菌转座子可以重新定位抗生素耐药基因,将其从染色体转移到质粒。 事实证明,这种机制是导致细菌性痢疾的志贺氏菌菌株中多个抗生素耐药基因在单个 R 质粒上共定位的原因。 这样的 R 质粒就可以很容易地通过偶联过程在细菌群体中转移。
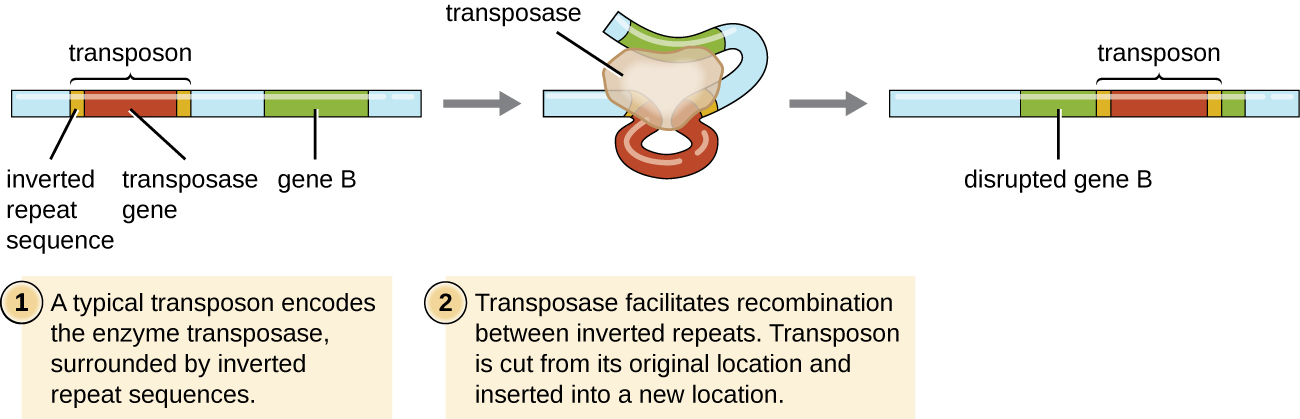
练习\(\PageIndex{6}\)
转座子可以通过哪两种方式影响其移动到的细胞的表型?
| 任期 | 定义 |
|---|---|
| 共轭 | 使用共轭 pilus 通过直接接触转移 DNA |
| 转导 | 细菌中水平基因转移的机制,其中基因是通过病毒感染转移的 |
| 转型 | 水平基因转移机制,其中裸露的环境 DNA 被细菌细胞吸收 |
| 换位 | DNA 从 DNA 分子中的一个位置独立切除并在其他位置整合的过程 |
临床重点:第 3 部分
尽管抗生素治疗仍在继续,但马克的感染继续迅速发展。 感染区域继续扩大,他必须安装呼吸机以帮助他呼吸。 马克的医生下令手术切除受感染的组织。 初次手术后,每天对马克的伤口进行监测,以确保感染不会复发,但会继续扩散。
经过另外两轮手术,感染似乎终于得到控制。 几天后,马克被从呼吸机上移开,得以自行呼吸。 但是,他的小腿失去了大量皮肤和软组织。
练习\(\PageIndex{7}\)
- 为什么去除受感染的组织会阻止感染?
- 这种治疗方法可能有哪些并发症?
关键概念和摘要
- 水平基因转移是无性繁殖生物(如原核生物)获得新性状的重要途径。
- 细菌通常使用三种水平基因转移机制:转化、转导和偶联。
- 转化允许有能力的细胞吸收死亡时从其他细胞释放的裸体DNA进入细胞质,在那里它可以与宿主基因组重组。
- 在广义转导中,任何染色体DNA片段都可能通过意外将降解的宿主染色体封装到噬菌体头中而转移。 在专门的转导中,只有与溶血性噬菌体整合位点相邻的染色体DNA才能由于前体切除不精确而转移。
- 共轭由 F 质粒介导,该质粒编码共轭 pilus,使含有 F 质粒的 F + 细胞与 F 细胞接触。
- F 质粒罕见地整合到细菌染色体中,产生 Hfr 细胞,允许染色体 DNA 从捐赠者转移到受体。 此外,不精确地从染色体上切除 F 质粒可能会产生 F' 质粒,该质粒可以通过偶联转移给受者。
- R 质粒的共轭转移是细菌群落中抗生素耐药性传播的重要机制。
- 转座子是末端有倒置重复序列的 DNA 分子,它还对酶转座酶进行编码,允许它们从 DNA 中的一个位置移动到另一个位置。 尽管转座子存在于原核生物和真核生物中,但转座子在细菌病原体中与毒力因子(包括抗生素耐药基因)的运动具有临床相关性。