
11.3: RNA 转录
- Page ID
- 200148

学习目标
- 解释如何使用 DNA 作为模板合成 RNA
- 区分原核生物和真核生物中的转录
在转录过程中,一个或多个基因的DNA序列中编码的信息被转录成一条RNA链,也称为RNA转录本。 由此产生的单链 RNA 分子由含有腺嘌呤 (A)、胞嘧啶 (C)、鸟嘌呤 (G) 和尿嘧啶 (U) 碱基的核糖核苷酸组成,充当原始 DNA 序列的移动分子拷贝。 原核生物和真核生物中的转录需要 DNA 双螺旋在 RNA 合成区域部分解开。 展开区域称为转录气泡。 特定基因的转录总是来自充当模板的两条 DNA 链中的一条,即所谓的反义链。 RNA 产物与 DNA 的模板链互补,与非模板 DNA 链或感官链几乎相同。 唯一的区别是,在 RNA 中,所有 T 核苷酸都被 U 核苷酸所取代;在 RNA 合成过程中,当互补反义链中有 A 时,U 就会被掺入。
细菌中的转录
细菌使用相同的 RNA 聚合酶来转录所有基因。 与DNA聚合酶一样,RNA聚合酶将核苷酸逐一添加到不断增长的核苷酸链的3'-OH组中。 DNA聚合酶和RNA聚合酶之间活性的一个关键区别是需要在3'-OH上添加核苷酸:DNA聚合酶需要这样的3'-OH基团,因此需要引物,而RNA聚合酶不需要。 在转录过程中,将与DNA模板链互补的核糖核苷酸添加到不断增长的RNA链中,并通过新添加的核苷酸与最后添加的核苷酸之间的脱水合成形成共价磷酸二酯键。 在大肠杆菌中,RNA聚合酶包含六个多肽亚基,其中五个构成聚合酶核心酶,负责向生长链中添加RNA核苷酸。 第六个亚单位被称为西格玛 (α)。 α 因子使 RNA 聚合酶能够与特定的启动子结合,从而允许转录各种基因。 有多种α因子允许转录各种基因。
启动
转录从启动子开始,启动子是转录机制与之结合并启动转录的 DNA 序列。 DNA双螺旋中与第一个5'RNA核苷酸转录位点相对应的核苷酸对是起始位点。 起始位点之前的核苷酸被指定为 “上游”,而起始位点之后的核苷酸被称为 “下游” 核苷酸。 在大多数情况下,启动子位于它们调节的基因的上游。 尽管启动子序列因细菌基因组而异,但一些元素是保守的。 在起始位点(指定为+1)之前的DNA中的—10和—35位置,有两个启动子共识序列或所有启动子和不同细菌物种之间相似的区域。 被称为 TATA 盒的 —10 共识序列是 TATAAT。 —35 序列被识别,并由 α 绑定。
伸长率
转录阶段的伸长始于 α 亚基与聚合酶分离,从而使核心酶能够以每秒大约 40 个核苷酸的速度在 5' 到 3' 的方向上合成与 DNA 模板互补的 RNA。 随着伸长率的推进,DNA在核心酶前面不断展开,然后在核心酶后面倒带(图\(\PageIndex{1}\))。
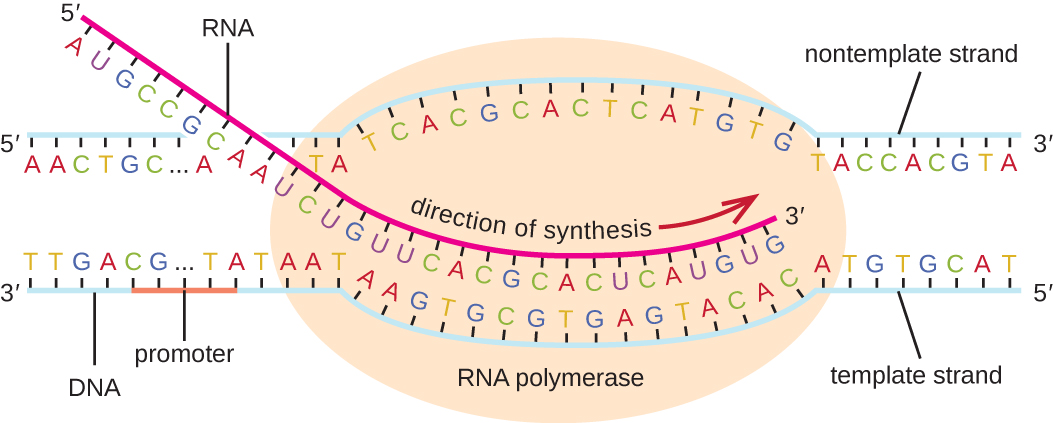
终止
基因被转录后,细菌聚合酶必须与DNA模板分离并释放出新产生的RNA。 这被称为转录的终止。 DNA 模板包含重复的核苷酸序列,这些序列充当终止信号,导致 RNA 聚合酶停滞并从 DNA 模板中释放,从而释放 RNA 转录本。
练习\(\PageIndex{1}\)
- RNA 聚合酶的 β 因子在哪里结合 DNA 开始转录?
- 什么会启动 RNA 聚合酶的聚合活性?
- 结束转录的信号来自哪里?
真核生物中的转录
原核生物和真核生物的转录过程基本相同,但有一些显著差异(见表\(\PageIndex{1}\))。 真核生物使用三种不同的聚合酶,即 RNA 聚合酶 I、II 和 III,它们在结构上都与细菌 RNA 聚合酶不同。 每种转录不同的基因子集。 有趣的是,古细菌含有一种单一的RNA聚合酶,该聚合酶与真核生物RNA聚合酶II的关系比与细菌对应物的关系更为密切。 真核生物 mRNA 通常也是 monocistronic,这意味着它们各自只编码一种多肽,而细菌和古细菌的原核 mRNA 通常是多环共体,这意味着它们编码多种多肽。
原核生物和真核生物之间最重要的区别是后者的膜结合核,这影响了使用RNA分子进行蛋白质合成的便捷性。 当基因结合在细胞核中后,真核细胞必须将编码蛋白质的 RNA 分子运送到要翻译的细胞质。 编码蛋白质的初级转录本,即由RNA聚合酶直接合成的RNA分子,必须经过多个处理步骤,以保护这些RNA分子在从细胞核转移到细胞质并转化为蛋白质期间免受降解。 例如,真核生物 mRNA 可能持续几个小时,而典型的原核生物 mRNA 的持续时间不超过 5 秒。
初级转录本(也称为 pre-mRNA)首先涂有 RNA 稳定蛋白,以保护其在处理和输出到细胞核外时免受降解。 第一种处理在初级转录本仍在合成过程中开始;在生长转录本的5'末端添加一种特殊的7-甲基鸟苷核苷酸,称为5'上限。 除了防止降解外,后续蛋白质合成所涉及的因素还可以识别上限,这有助于启动核糖体的翻译。 伸长完成后,另一种加工酶会在 3' 末端添加一串大约 200 个腺嘌呤核苷酸,称为 Poly-A 尾巴。 这种修饰进一步保护 pre-mRNA 免受降解,并向细胞因子发出信号,表明转录本需要输出到细胞质中。
编码多肽的真核生物基因由称为外显子(ex-on 表示它们被压制)的编码序列和称为内含子的中间序列(int-ron 表示它们的 int ervening 作用)组成。 与内含子对应的转录RNA序列不编码功能性多肽的区域,并在处理过程中从pre-mRNA中移除。 在蛋白质合成之前,必须将所有内含子编码的 RNA 序列从前 mRNA 中完全精确地去除,这样外显子编码的 RNA 序列才能正确连接在一起编码功能性多肽。 如果该过程出现单个核苷酸的错误,则重新连接的外显子的序列就会发生转移,由此产生的多肽将失效。 去除内含子编码的 RNA 序列并重新连接外显子编码的 RNA 序列的过程称为 RNA 拼接,由含有小核核糖核蛋白 (snRNP) 的剪接体的作用促进了这种过程。 内含子编码的 RNA 序列在 pre-mRNA 中时被移除。 尽管内含子没有被翻译,但它们似乎具有各种功能,包括基因调控和mRNA转运。 完成这些修饰后,成熟的转录本,即编码多肽的mRNA,被运出细胞核,运往细胞质进行翻译。 内含子可以用不同的方式拼接出来,从而在最终的mRNA产物中包含或排除各种外显子。 此过程被称为替代拼接。 替代拼接的优点是可以生成不同类型的 mRNA 转录本,所有转录本都来自相同的 DNA 序列。 近年来,研究表明,一些古细菌也有能力拼接其之前的mRNA。
| 财产 | 细菌 | 真核生物 |
|---|---|---|
| 每个 mRNA 编码的多肽数量 | Monocistronic 或 polycistroni | 只有 monocistronic |
| 股线伸长率 | 核心 + α = 全酵素 | RNA 聚合酶 I、II 或 III |
| 增加了 5 英尺的上限 | 不 | 是的 |
| 添加 3' Poly-A 尾巴 | 不 | 是的 |
| pre-mRNA 的拼接 | 不 | 是的 |
练习\(\PageIndex{2}\)
- 在真核细胞中,蛋白质基因的RNA转录本在转录后如何修改?
- 外显子或内含子是否包含蛋白质序列的信息?
临床重点:第 2 部分
在急诊室,一位护士告诉马克,他做出了去医院的明智决定,因为他的症状表明感染已经失控。 马克的症状有所改善,皮肤区域受到影响,肿胀量增加。 在患处,皮疹已经开始,起泡,皮肤最外层下方形成了小气穴,有些皮肤变灰了。 基于其中一个水泡排出的脓液的腐臭气味、感染的迅速进展以及受影响皮肤的视觉外观,医生立即开始治疗坏死性筋膜炎。 马克的医生下令对从水泡中排出的液体进行培养,还下令进行血液检查,包括白细胞计数。
马克被送进重症监护室并开始静脉注射广谱抗生素,以尽量减少感染的进一步传播。 尽管接受了抗生素治疗,但马克的病情迅速恶化。 马克变得困惑和头晕。 在他住院后的几个小时内,他的血压明显下降,呼吸变得更浅更快。 此外,水泡增加,水泡的颜色变为紫黑色,伤口本身似乎正在迅速向上移动 Mark 的腿部。
练习\(\PageIndex{3}\)
- 马克坏死性筋膜炎的可能病因有哪些?
- 为什么抗生素治疗似乎不起作用,有哪些可能的解释?
关键概念和摘要
- 在转录过程中,DNA 中编码的信息用于制造 RNA。
- RNA聚合酶使用DNA的反义链作为模板,通过在生长链的3'末端添加互补的RNA核苷酸来合成RNA。
- 在转录开始期间,RNA 聚合酶以称为启动子的序列与 DNA 结合。
- 编码具有相关功能的蛋白质的基因经常在原核生物中的单个启动子的控制下被转录,从而形成编码多个多肽的多环共振mRNA 分子。
- 与DNA聚合酶不同,RNA聚合酶不需要3'-OH基团即可添加核苷酸,因此启动时不需要引物。
- 当 RNA 聚合酶遇到导致聚合酶失效的特定 DNA 序列时,就会@@ 终止细菌中的转录。 这会导致从 DNA 模板链中释放 RNA 聚合酶,从而释放 RNA 转录本。
- 真核生物有三种不同的RNA聚合酶。 真核生物还有 monocistronic mRNA,每种只编码一种多肽。
- 真核生物初级转录本通过多种方式进行处理,包括添加 5' 帽和 3' -poly-A 尾巴,以及拼接,以生成成熟的 mRNA 分子,该分子可以从细胞核中输出,并且不受降解。