
8.6:光合作用和光的重要性
- Page ID
- 200198

学习目标
- 描述真核生物和原核生物中光合色素的功能和位置
- 描述光依赖和光无关反应的主要产物
- 描述光合细胞中产生葡萄糖的反应
- 比较和对比循环和非环光磷酸化
从大肠杆菌到人类的异养生物依赖主要存在于碳水化合物分子中的化学能。 这些碳水化合物中有许多是由光合作用产生的,光合作用是光养生物将太阳能(阳光)转化为化学能的生化过程。 尽管光合作用最常与植物有关,但微生物光合作用也是化学能的重要来源,为许多不同的生态系统提供了动力。 在本节中,我们将重点介绍微生物的光合作用。
光合作用分为两个连续阶段:依赖光的反应和与光无关的反应(图\(\PageIndex{1}\))。 在依赖光的反应中,来自阳光的能量被光合膜中的色素分子吸收,并转化为储存的化学能。 在与光无关的反应中,光依赖反应产生的化学能量用于使用二氧化碳驱动糖分子的组装;但是,这些反应仍然是光依赖性的,因为驱动它们所需的光依赖反应的产物很短-活过。 依赖光的反应会产生 ATP 和 NADPH 或 NADH 来暂时储存能量。 这些能量载体用于与光无关的反应,以驱动能量上不利的过程,即将无机二氧化碳 “固定” 为有机形式,即糖。
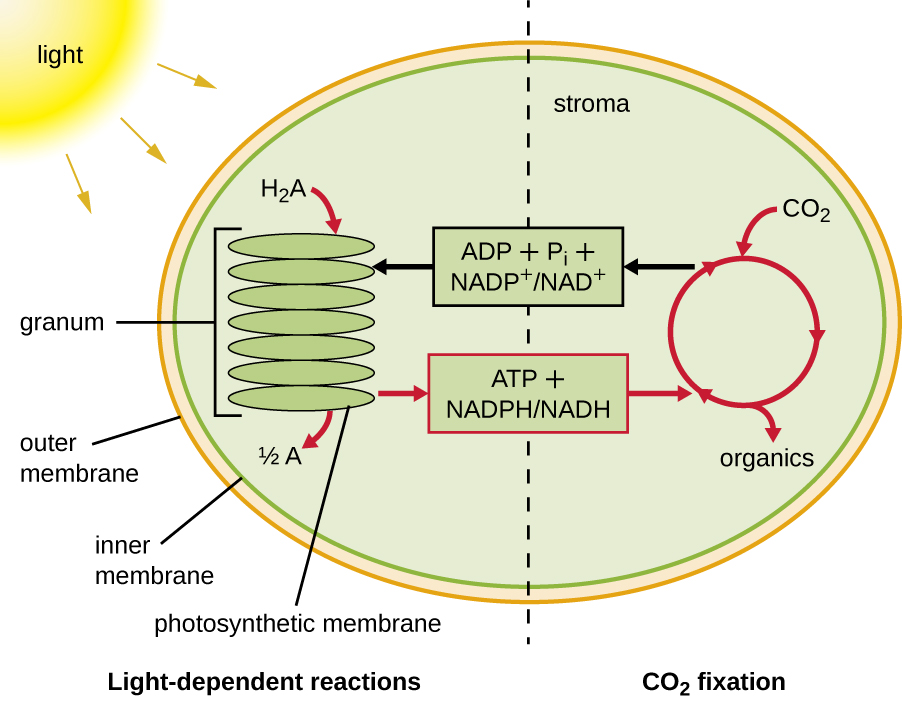
真核生物和原核生物中的光合结构
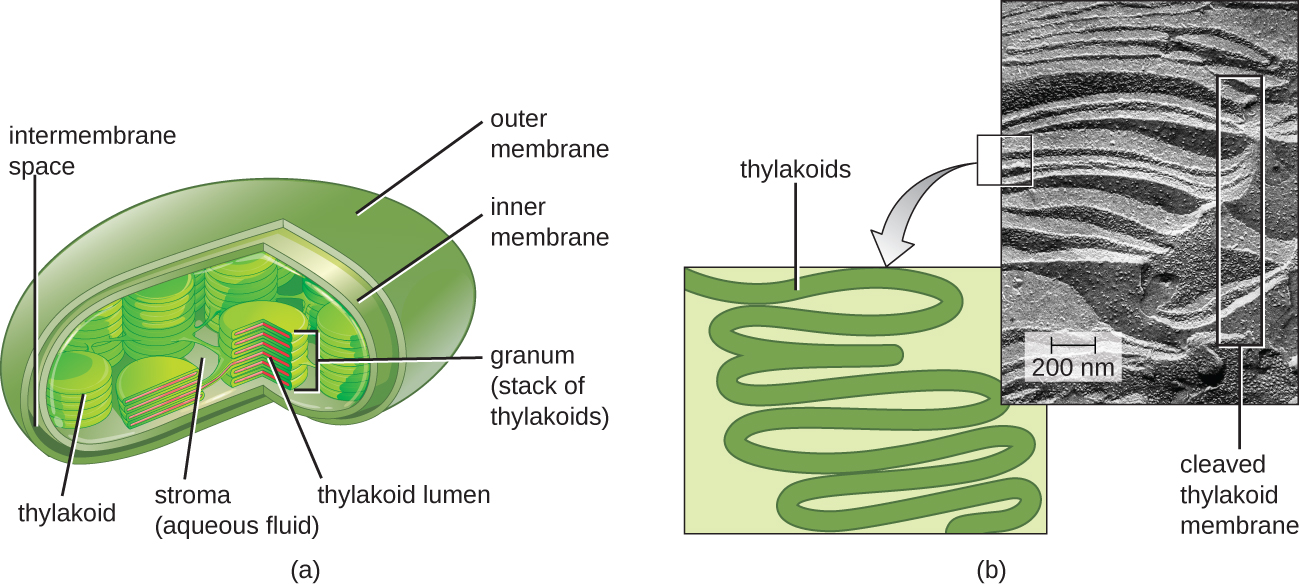
在所有光养真核生物中,光合作用发生在叶绿体内,叶绿体是一种通过光合细菌的内共生作用在真核生物中产生的细胞器(参见真核细胞的独特特征)。 这些叶绿体被具有内层和外层的双层膜所包围。 叶绿体内有第三层膜,它形成堆叠的圆盘状光合结构,称为 thylakoids(图\(\PageIndex{2}\))。 一堆 thylakoids 被称为颗粒,叶绿体内颗粒周围的空间称为基质。
相比之下,原核生物中的光合膜不是组织成不同的膜封闭细胞器;相反,它们是质膜的折叠区域。 例如,在蓝细菌中,这些折叠区域也被称为 thylakoids。 无论哪种情况,嵌入在 tylakoid 膜或其他光合细菌膜中的都是组织成一个或多个光系统的光合色素分子,其中光能实际上被转化为化学能。
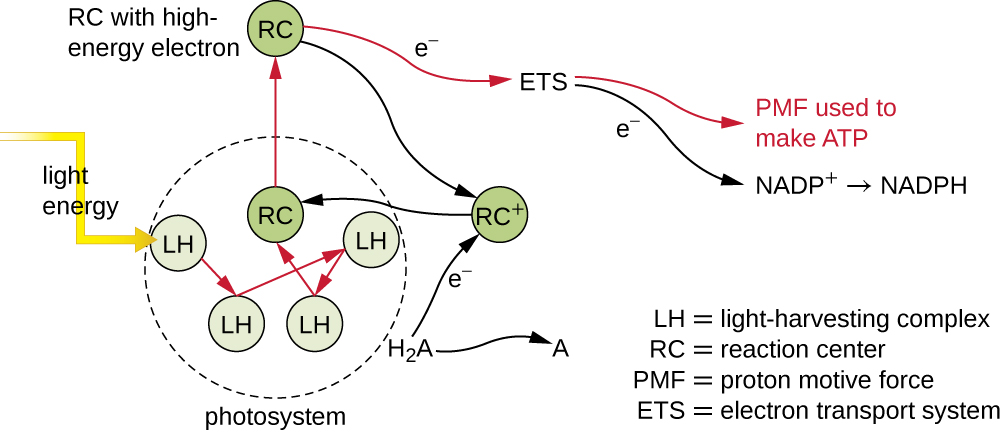
光合膜内的光合色素被组织成光系统,每个光系统都由光收集(触角)复合物和反应中心组成。 光采复合物由多种蛋白质和相关色素组成,每种蛋白质和相关色素都可能吸收光能,从而变得兴奋。 这种能量从一个色素分子转移到另一个色素分子,直到最终(大约百万分之一秒后)它被输送到反应中心。 到目前为止,只有能量——不是电子——在分子之间传输。 反应中心含有一种色素分子,它在激发时会发生氧化,实际上会放弃电子。 正是在光合作用的这一步,光能被转化为激发的电子。
不同种类的采光颜料会吸收可见光的独特波长(颜色)模式。 颜料会反射或传输它们无法吸收的波长,使它们呈现出相应的颜色。 光合色素(用于吸收太阳能的分子)的例子包括细菌叶绿素(绿色、紫色或红色)、类胡萝卜素(橙色、红色或黄色)、叶绿素(绿色)、藻蓝素(蓝色)和藻红素(红色)。 通过混合颜料,生物体可以吸收来自更多波长的能量。 由于光合细菌通常在争夺阳光时生长,因此每种类型的光合细菌都经过优化,可以收集其通常暴露的波长的光,从而导致水生和土壤生态系统中的微生物群落因光质和渗透率而分层。
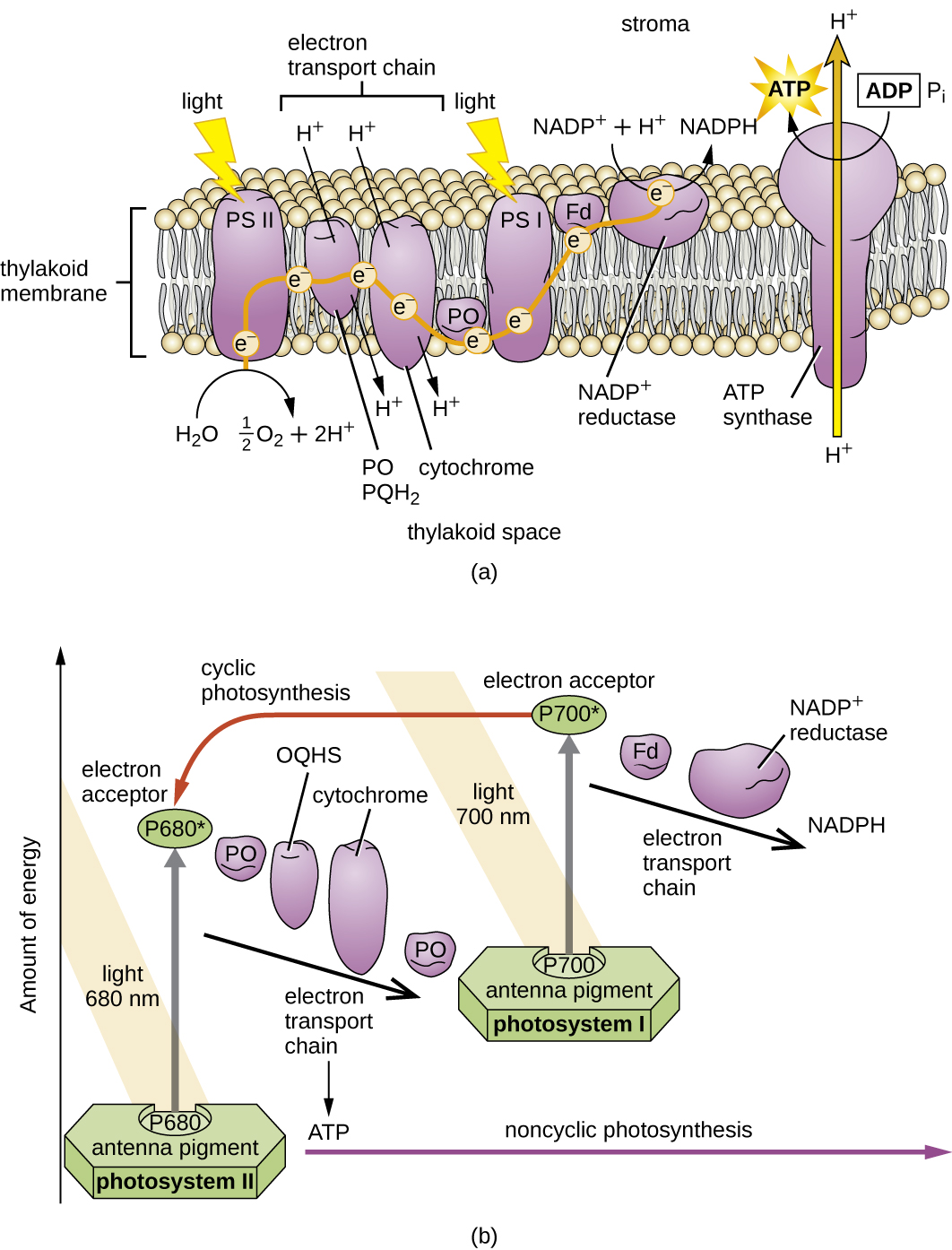
一旦光采集复合体将能量转移到反应中心,反应中心就会将其高能电子一个接一个地输送到电子传输系统中的电子载体,然后开始通过 ETS 进行电子传输。 ETS 与用于细胞呼吸的 ETS 类似,嵌入在光合膜中。 最终,电子被用来产生 NADH 或 NADPH。 在光合膜上形成的电化学梯度用于通过光磷酸化过程通过化学渗透产生 ATP,这是氧化磷酸化的另一个例子(图\(\PageIndex{3}\))。
练习\(\PageIndex{1}\)
在光养真核生物中,光合作用发生在哪里?
氧气和缺氧光合作用
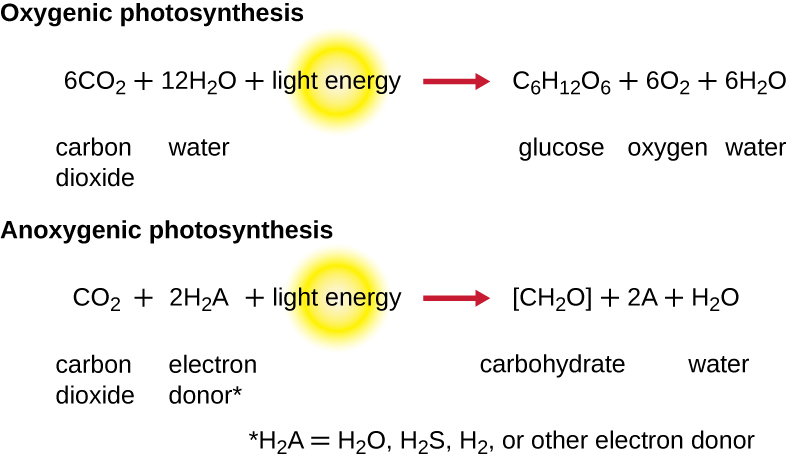
为了继续进行光合作用,必须更换从反应中心色素中丢失的电子。 这种电子的来源(H 2 A)将植物和蓝细菌的氧气光合作用与其他类型的细菌光养体进行的缺氧光合作用区分开来(图\(\PageIndex{4}\))。 在氧气光合作用中,H 2 O 被分裂并将电子供应到反应中心。 由于氧气作为副产品产生并被释放,因此这种光合作用被称为氧气光合作用。 但是,当其他还原化合物作为电子供体时,不会产生氧气;这些类型的光合作用被称为缺氧光合作用。 硫化氢(H 2 S)或硫代硫酸盐\(\ce{(S2O3^{2-})}\)可以作为电子供体,从而分别产生元素硫和硫酸根\(\ce{(SO4^{2-})}\)离子。
光系统分为两种类型:光系统 I(PSI)和光系统 II(PSII)(图\(\PageIndex{5}\))。 蓝细菌和植物叶绿体同时具有光系统,而缺氧光合细菌仅使用其中一种光系统。 两个光系统同时被光能激发。 如果细胞同时需要 ATP 和 NADPH 进行生物合成,则它将进行非环光磷酸化。 将PSII反应中心电子传递给连接PSII和PSI的ETS后,从PSII反应中心丢失的电子将被水分裂所取代。 激发的 PSI 反应中心电子用于将 NADP + 还原为 NADPH,取而代之的是退出 ETS 的电子。 这种方式的电子流动被称为 Z 方案。
如果细胞对ATP的需求明显超过其对NADPH的需求,则它可能会绕过通过循环光磷酸化产生的还原力。 在循环光磷酸化过程中只使用 PSI;PSI 反应中心的高能电子被传递到 ETS 载体,然后最终返回到氧化的 PSI 反应中心颜料,从而还原它。
练习\(\PageIndex{2}\)
为什么光合细菌会有不同的色素?
与光无关的反应
在来自太阳的能量转化为化学能并暂时储存在ATP和NADPH分子(寿命为百万分之一秒)中之后,photoautotrophs就有了制造多碳碳水化合物分子所需的燃料,这种分子可以长期存活数亿年能量存储。 碳来自二氧化碳,这种气体是细胞呼吸的废物。
卡尔文-本森循环(以梅尔文·加尔文 [1911—1997] 和安德鲁·本森 [1917—2015] 命名)是用于固定二氧化碳的生化途径,位于光合细菌的细胞质和真核生物叶绿体的基质中。 加尔文循环中与光无关的反应可以分为三个基本阶段:固定、还原和再生(有关加尔文循环的详细说明,请参阅附录C)。
- 固定:核酮糖二磷酸羧化酶(Rubisco)酶催化向二磷酸核酮糖(ruBP)中添加二氧化碳。 这导致产生 3-磷酸甘油酯 (3-PGA)。
- 还原:使用六个 ATP 和 NADPH 分子(来自光依赖反应)将 3-PGA 转化为 3-磷酸甘油醛(G3P)。 然后使用一些 G3P 来生成葡萄糖。
- 再生:未用于合成葡萄糖的剩余的 G3P 用于再生 RuBP,使系统能够继续固定 CO 2。 在这些再生反应中又使用了三个 ATP 分子。
加尔文循环被植物和光自养细菌广泛使用,据说RuBisco是地球上最丰富的酶,占植物叶绿体总可溶性蛋白的30%至50%。 1 但是,除了在 photoautotrophs 中普遍使用加尔文循环外,许多非光合化学自养生物还使用加尔文循环来修复二氧化碳。 此外,其他细菌和古细菌使用替代系统固定 CO 2。 尽管大多数使用加尔文循环替代品的细菌都是化学自养的,但某些绿色硫光自养细菌也被证明使用替代的二氧化碳固定途径。
练习\(\PageIndex{3}\)
描述加尔文循环的三个阶段。
关键概念和摘要
- Heterotrophs 依赖于 autotrophs 产生的碳水化合物,其中许多是光合的,可以将太阳能转化为化学能。
- 不同的光合生物使用不同的光合颜料混合物,这增加了生物体可以吸收的光的波长范围。
- 光系统(PSI 和 PSII)均包含一种光采复合物,由多种蛋白质和吸收光能的相关色素组成。 光合作用的光依赖反应将太阳能转化为化学能,产生ATP和NADPH或NADH来暂时储存这种能量。
- 在氧气光合作用中,H 2 O 充当电子供体,取代反应中心电子,氧气作为副产物形成。 在缺氧光合作用中,其他还原分子,如H 2 S或硫代硫酸盐可用作电子供体;因此,氧气不是作为副产物形成的。
- 当需要同时产生 ATP 和 NAD@@ PH 时,非环光磷酸化用于氧气光合作用。 如果细胞对 ATP 的需求超过其对 NADPH 的需求,那么它可能会进行循环光磷酸化,只产生 ATP。
- 光合作用与光无关的反应使用光依赖反应中的ATP和NADPH将二氧化碳固定到有机糖分子中。
脚注
- 1 A. Dhingra 等人。 “增强叶绿体表达的 Rbc S 基因的翻译可恢复核 Rb c S 反义植物中的小亚基水平和光合作用。” 《美利坚合众国国家科学院院刊》 101 第 16 期 (2004): 6315—6320。