
8.2: 碳水化合物的分解代谢
- Page ID
- 200213

学习目标
- 描述为什么糖酵解不依赖氧气
- 定义和描述糖酵解产生的三碳分子、ATP 和 NADH 的净产量
- 解释三碳丙酮酸分子是如何转化为双碳乙酰基团的,这些基团可以汇入克雷布斯循环。
- 定义和描述克雷布斯周期中二氧化碳、GTP/ATP、FADH 2 和 NADH 的净收益率
- 解释如何在细胞中使用克雷布斯循环的中间碳分子
有大量的酶途径可以分解碳水化合物以捕获 ATP 键中的能量。 此外,许多分解代谢途径会产生中间分子,这些中间分子也被用作合成代谢的基石。 了解这些过程很重要,原因有很多。 首先,由于所涉及的主要代谢过程是各种化学异养生物所共有的,因此我们可以通过研究大肠杆菌等更容易操纵的细菌的新陈代谢来学到很多关于人类新陈代谢的知识。 其次,由于动物和人类病原体也是化学异养生物,因此了解这些细菌中新陈代谢的细节,包括细菌和人类途径之间可能存在的差异,对于诊断病原体以及发现靶向抗微生物疗法很有用特定病原体。 最后,专门了解化学异养代谢所涉及的途径也可以作为比较微生物使用的其他更不寻常的代谢策略的基础。 尽管 chemoheterorophs 和 chemoautotrophs 之间引发电子转移的电子的化学来源不同,但这两种生物中都使用了许多相似的过程。
向学生介绍新陈代谢概念的典型例子是碳水化合物分解代谢。 对于 chemoheterotrophs 来说,我们的新陈代谢例子从糖原、淀粉或纤维素等多糖的分解代谢开始。 诸如分解糖原或淀粉的淀粉酶和分解纤维素的纤维素酶等酶可以导致这些聚合物中葡萄糖单体之间的糖苷键水解,从而释放葡萄糖以进一步分解代谢。
糖酵解
对于细菌、真核生物和大多数古细菌来说,糖酵解是葡萄糖分解代谢的最常见途径;它产生能量、减少的电子载体和细胞代谢的前体分子。 每个活生物体都会进行某种形式的糖酵解,这表明这种机制是一种古老的普遍代谢过程。 该过程本身不使用氧气;但是,糖酵解可以与有氧或厌氧的其他代谢过程相结合。 糖酵解发生在原核和真核细胞的细胞质中。 它从一个单一的六碳葡萄糖分子开始,最后是两个叫做丙酮酸的三碳糖分子。 丙酮酸在糖酵解后可能会被进一步分解,以便通过有氧或厌氧呼吸利用更多能量,但是许多生物,包括许多微生物,可能无法呼吸;对于这些生物来说,糖酵解可能是它们产生 ATP 的唯一来源。
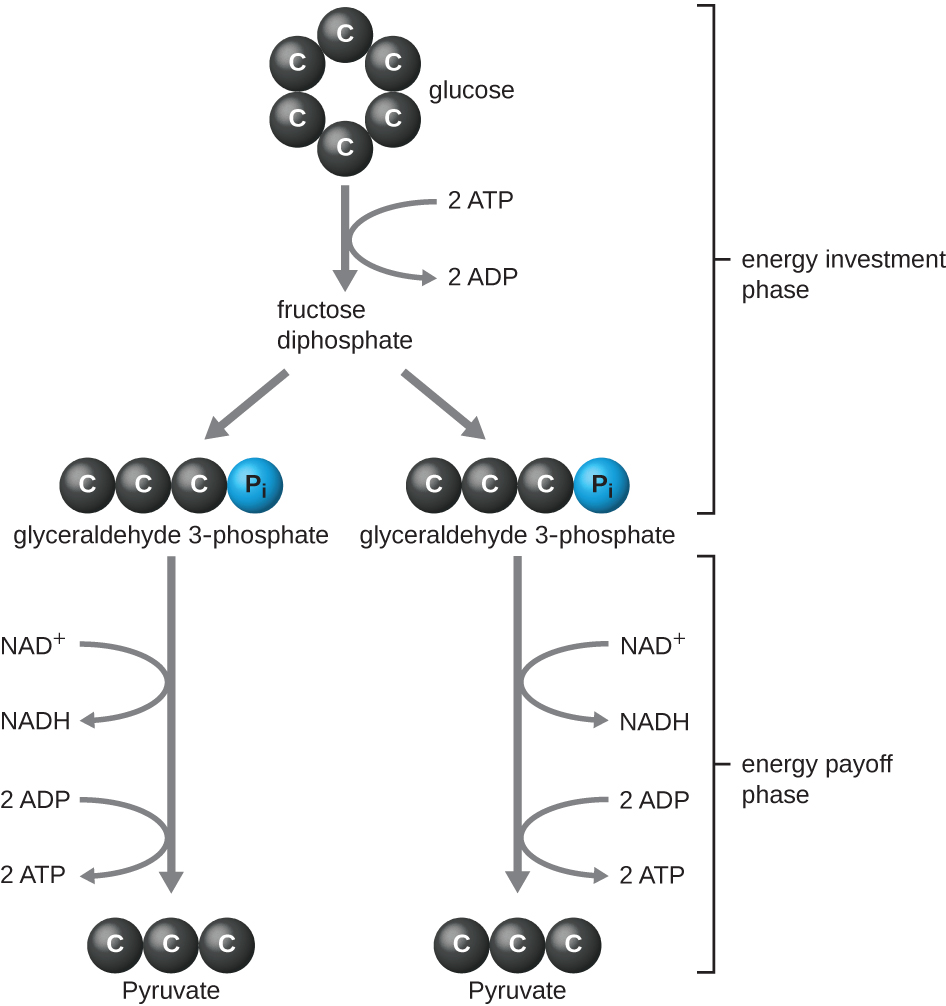
动物体内发现的、在微生物中最常见的糖酵解类型是 Embden-Meyerhof-Parnas(EMP)途径,它以古斯塔夫·恩布登(1874—1933)、奥托·迈耶霍夫(1884—1951)和雅各布·帕纳斯(1884—1949)命名。 使用EMP途径进行糖酵解包括两个不同的阶段(图\(\PageIndex{1}\))。 该路径的第一部分称为能量投资阶段,它利用来自两个ATP分子的能量来修饰葡萄糖分子,这样六碳糖分子就可以均匀地分裂成两个磷酸化三碳分子,称为甘油醛3-磷酸(G3P)。 该路径的第二部分称为能量回报阶段,它通过将 G3P 氧化为丙酮酸来提取能量,使用源自葡萄糖的电子,产生四个 ATP 分子,并将两个 NAD + 分子还原为两个 NADH 分子。 (附录C 中有关于具有化学结构和酶名称的完整EMP途径的讨论和说明。)
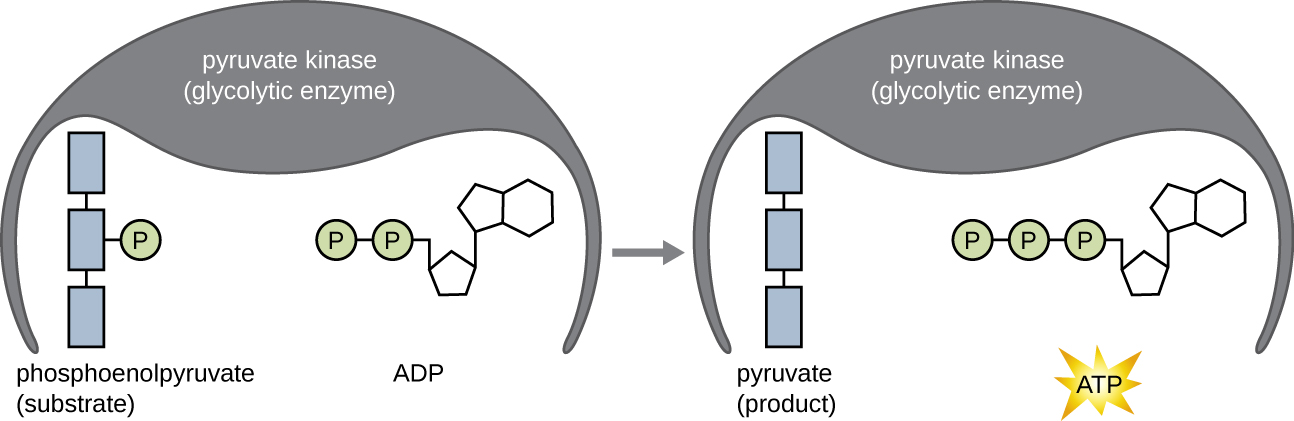
在糖酵解的能量回报阶段产生的ATP分子是由底物水平的磷酸化形成的(图\(\PageIndex{1}\)),这是产生ATP的两种机制之一。 在底物级磷酸化中,从有机分子中去除磷酸基团,然后直接转移到可用的 ADP 分子中,产生 ATP。 在糖酵解过程中,来自中间分子的高能磷酸基团被添加到 ADP 中以产生 ATP。
总体而言,在这个糖酵解过程中,单个葡萄糖分子分解的净收益为:
- 两个 ATP 分子
- 两个 NADH 分子,以及
- 两个丙酮酸分子。
其他糖酵解途径
当我们提到糖酵解时,除非另有说明,否则我们指的是动物和许多细菌使用的EMP途径。 但是,一些原核生物使用其他糖酵解途径。 一个重要的替代方案是恩特纳-杜多罗夫(ED)路径,它以其发现者内森·恩特纳和迈克尔·杜多罗夫(1911-1975 年)的名字命名。 尽管某些细菌,包括机会性革兰氏阴性病原体铜绿假单胞菌,仅含有糖酵解的 ED 途径,但其他细菌,例如大肠杆菌,有能力使用 ED 途径或 EMP 途径。
存在于所有细胞中的第三种糖酵解途径是磷酸戊糖途径(PPP),也称为磷酸葡萄糖酸盐途径或己糖单磷酸盐分流通路,与前两种途径大不相同。 有证据表明,PPP可能是最古老的通用糖酵解途径。 来自PPP的中间体用于核苷酸和氨基酸的生物合成。 因此,当细胞分别需要核酸和/或蛋白质合成时,这种糖酵解途径可能会受到青睐。 附录 C 中对完整的 ED 途径和具有化学结构和酶名称的 PPP 进行了讨论和说明。
练习\(\PageIndex{1}\)
生物体什么时候可以使用 ED 途径或 PPP 进行糖酵解?
过渡反应、辅酶 A 和克雷布斯循环
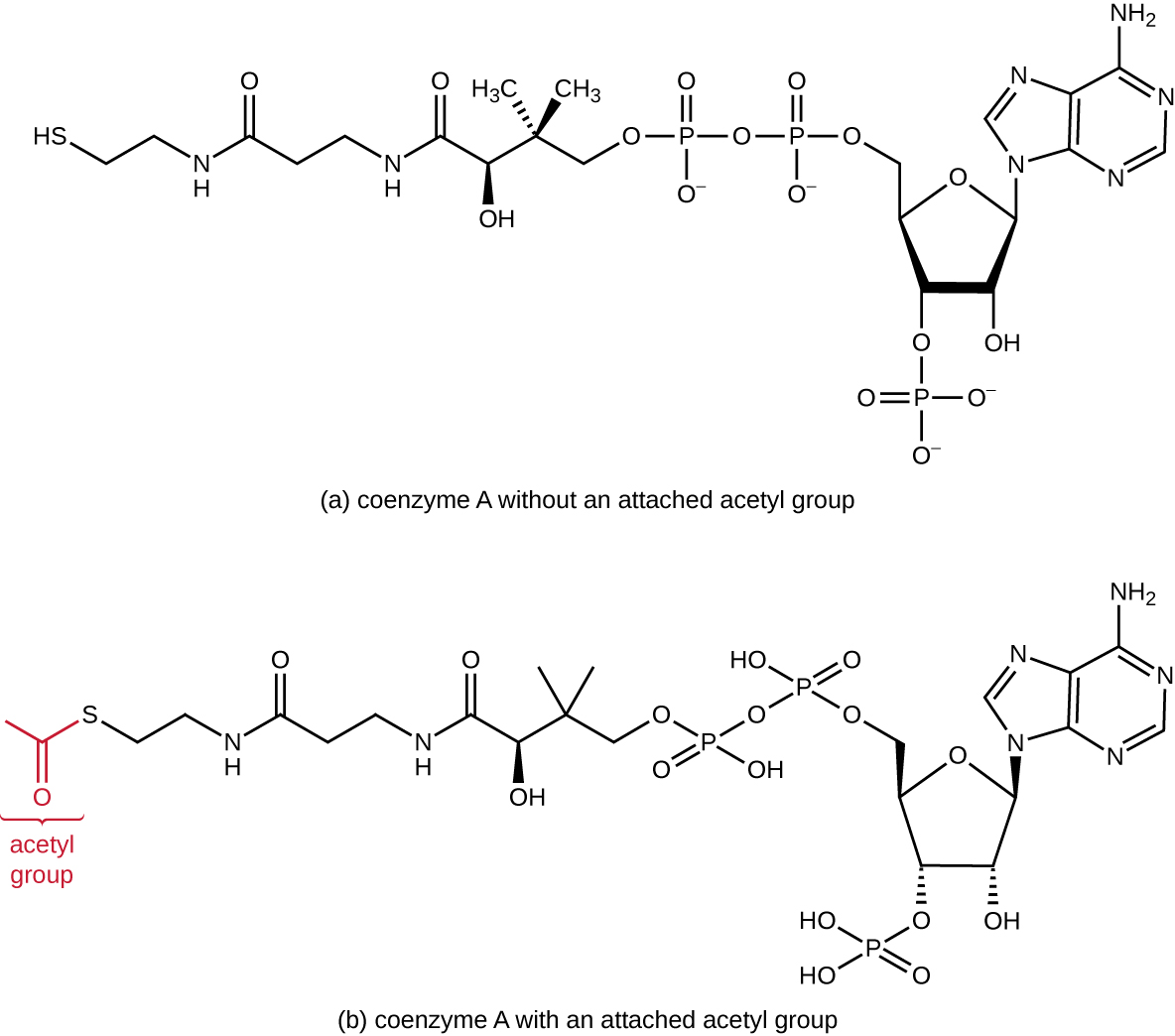
糖酵解会产生丙酮酸,丙酮酸可以进一步氧化以捕获更多能量。 要使丙酮酸进入下一个氧化途径,必须首先通过酶复合物丙酮酸脱氢酶在过渡反应(也称为桥反应)中将其脱羧为双碳乙酰基(见附录C 和图\(\PageIndex{3}\))。 在过渡反应中,电子也被转移到 NAD + 以形成 NADH。 要进入代谢过程的下一阶段,必须将相对较小的二碳乙酰附着在一种名为辅酶A(CoA)的非常大的载体化合物上。 过渡反应发生在真核生物的线粒体基质中;在原核生物中,它发生在细胞质中,因为原核生物缺乏膜封闭的细胞器。
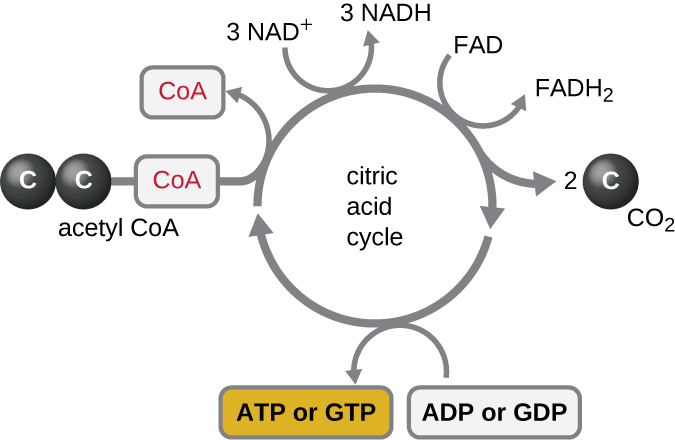
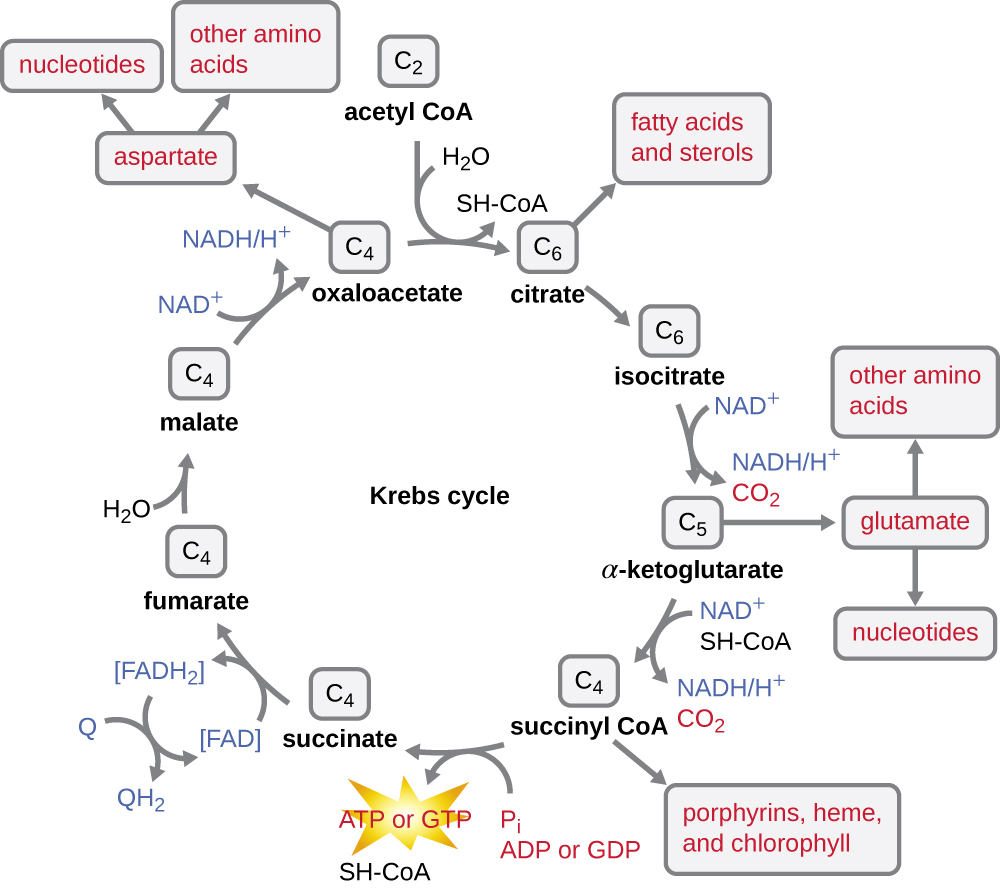
克雷布斯循环将过渡反应期间产生的乙酰基中的剩余电子转移到电子载体分子,从而减少它们。 克雷布斯循环也与糖酵解和过渡反应一起发生在原核生物的细胞质中,但它发生在真核细胞的线粒体基质中,过渡反应也发生在真核细胞的线粒体基质中。 克雷布斯循环以其发现者英国科学家汉斯·阿道夫·克雷布斯(Hans Adolf Krebs)(1900—1981)命名,也被称为柠檬酸循环或三羧酸循环(TCA),因为柠檬酸的结构中有三个羧基。 与糖酵解不同,克雷布斯循环是一个闭环:该路径的最后一部分可再生第一步中使用的化合物(图\(\PageIndex{4}\))。 循环的八个步骤是一系列化学反应,它们从过渡反应中捕获双碳乙酰基(CoA 载体不进入克雷布斯循环),过渡反应被添加到克雷布斯循环中的四碳中间体中,产生六碳中间体柠檬酸(给出此周期的备用名称)。 当循环的一圈回到四碳中间体的起点时,该循环会产生两个二氧化碳分子,一个由底物级磷酸化产生的 ATP 分子(或同等分子,例如三磷酸鸟苷 [GTP]),以及三个 NADH 分子和一个 FADH 2 分子 。 (整个克雷布斯周期的讨论和详细说明见附录C。)
尽管许多生物使用克雷布斯循环作为葡萄糖代谢的一部分,但克雷布斯循环中的几种中间化合物可用于合成各种重要的细胞分子,包括氨基酸、叶绿素、脂肪酸和核苷酸;因此,该循环是合成代谢和分解代谢(图\(\PageIndex{5}\))。
关键概念和摘要
- 糖酵解是葡萄糖分解的第一步,它会形成由底物级磷酸化、NADH 和两个丙酮酸分子产生的 ATP。 糖酵解不使用氧气,也不依赖氧气。
- 糖酵解后,三碳丙酮酸被脱羧形成双碳乙酰基,同时形成 NADH。 乙酰基附着在一种叫做辅酶 A 的大型载体化合物上
- 过渡步骤结束后,辅酶A将二碳乙酰输送到克雷布斯循环,两个碳进入循环。 每循环一圈,一个源自糖酵解的乙酰基被进一步氧化,通过底物级磷酸化产生三个 NADH 分子、一个 FADH 2 和一个 ATP,并释放出两个 CO 2 分子。
- 克雷布斯循环可用于其他目的。 许多中间体用于合成重要的细胞分子,包括氨基酸、叶绿素、脂肪酸和核苷酸。