
8.3: 细胞呼吸
- Page ID
- 200189

学习目标
- 比较和对比电子传输系统在原核细胞和真核细胞中的位置和功能
- 比较和对比底物水平和氧化磷酸化之间的差异
- 解释化学渗透和质子动力之间的关系
- 描述原核细胞与真核细胞中 ATP 合成酶的功能和位置
- 比较和对比有氧呼吸和厌氧呼吸
我们刚刚讨论了葡萄糖分解代谢的两种途径——糖酵解和克雷布斯循环,它们通过底物水平的磷酸化产生 ATP。 但是,大多数 ATP 是在称为氧化磷酸化的单独过程中产生的,该过程发生在细胞呼吸期间。 细胞呼吸始于电子从 NADH 和 FADH 2(糖酵解、过渡反应和克雷布斯循环中产生)通过一系列化学反应转移到最终的无机电子受体(要么是有氧呼吸中的氧气,要么是非氧无机分子)厌氧呼吸)。 这些电子转移发生在原核细胞细胞膜的内部或真核细胞线粒体内膜中的特殊蛋白质复合物中。 收集电子的能量以在膜上产生电化学梯度,该梯度用于通过氧化磷酸化产生 ATP。
电子传输系统
电子传输系统(ETS)是细胞呼吸过程中涉及的最后一个组件;它包括一系列与膜相关的蛋白质复合物和相关的移动辅助电子载体。 电子传输是一系列类似于桶装旅的化学反应,因为来自NADH和FADH 2 的电子从一个ETS电子载体迅速传递到另一个ETS电子载体。 这些载流子可以在 ETS 中传递电子,因为它们具有氧化还原电位。 要使蛋白质或化学物质接受电子,它必须具有比电子供体更正的氧化还原电位。 因此,电子从具有更负氧化还原电位的电子载流子转移到氧化还原电位更高的电子载流子。 真核和原核电子传输系统中涉及的四大类电子载流子是细胞色素、黄蛋白、铁硫蛋白和喹酮。
在有氧呼吸中,ETS末端的最终电子受体(即具有最大正氧化还原电位的那个)是氧分子(O 2),它被最终的ETS载体还原为水(H 2 O)。 这种电子载体细胞色素氧化酶因细菌类型而异,可用于区分密切相关的细菌以进行诊断。 例如,革兰氏阴性机会主义者铜绿假单胞菌和引起革兰氏阴性霍乱的霍乱弧菌使用细胞色素 c 氧化酶,这可以通过氧化酶测试检测,而其他革兰氏阴性肠杆菌科,如大肠杆菌,则对此呈阴性测试,因为它们会产生不同的细胞色素氧化酶类型。
在许多情况下无法进行有氧呼吸,包括以下任何一种或多种情况:
- 该细胞缺少编码适当的细胞色素氧化酶的基因,该酶用于在电子传输系统末端将电子转移到氧气中。
- 该细胞缺乏编码酶的基因,无法最大限度地减少有氧呼吸过程中产生的危险氧自由基(例如过氧化氢(H 2 O 2)或超氧化物)的严重破坏作用\((\ce{O2-})\)。
- 该细胞缺乏足够的氧气进行有氧呼吸。
有氧呼吸的一种可能的替代方法是厌氧呼吸,使用氧气以外的无机分子作为最终的电子受体。 在细菌和古细菌中发现了许多类型的厌氧呼吸。 反硝化物是重要的土壤细菌,它们使用硝酸盐\((\ce{NO3-})\)和亚硝酸盐\((\ce{NO2-})\)作为最终电子受体,产生氮气(N 2)。 许多有氧呼吸细菌,包括大肠杆菌,转而使用硝酸盐作为最终电子受体,并在氧气含量耗尽时产生亚硝酸盐。
使用厌氧呼吸的微生物通常具有完整的克雷布斯循环,因此这些生物可以获得形成的NADH和FADH 2 分子的能量。 但是,厌氧呼吸器使用由其基因组编码的改变的 ETS 载体,包括将电子转移到最终电子受体的不同复合物。 这些电子转移系统产生的电化学梯度较小,因此通过厌氧呼吸形成的ATP较少。
练习\(\PageIndex{1}\)
有氧呼吸和厌氧呼吸都使用电子传输链吗?
化学渗透、质子动力与氧化磷酸化
在电子通过 ETS 的每次转移中,电子都会失去能量,但是在某些转移中,通过使用能量将氢离子(H +)泵送到膜上,从而将能量存储为势能。 在原核细胞中,H + 被泵送到细胞质膜外部(在革兰氏阴性和革兰氏阳性细菌中称为周质空间);在真核细胞中,它们从线粒体基质通过线粒体内部膜泵送到膜间空间。 H + 在膜上的分布不均匀,这就形成了电化学梯度,因为 H + 离子具有正电荷(电),并且膜一侧的浓度(化学物质)更高。 这种由膜一侧的H +(也称为质子)与另一侧相比积累而形成的电化学梯度被称为质子动力(PMF)。 由于所涉及的离子是 H +,因此还会建立 pH 梯度,H + 浓度较高的膜侧的酸性更强。 正如本章所讨论的那样,除了使用 PMF 制造 ATP 之外,PMF 还可用于驱动其他能量不利的过程,包括营养物输送和鞭毛旋转以提高运动力。
ETS 产生的这种电化学梯度的势能导致 H + 在膜(原核细胞中的质膜和真核细胞中线粒体中的内膜)中扩散。 这种氢离子穿过膜的流动,称为化学渗透,必须通过一种称为ATP合成酶的膜结合酶复合物通过膜中的通道发生(图\(\PageIndex{1}\))。 以这种方式移动的趋势很像大坝一侧积聚的水,打开时会穿过大坝。 ATP 合成酶(比如水力发电大坝的摄入量和生成器的组合)是一种复杂的蛋白质,它起着微小的生成器的作用,在 H + 通过酶扩散的力量下,它们的电化学梯度从许多相互排斥的 H + 向下移动其中 H + 较少。 在原核细胞中,H + 从细胞质膜外部流入细胞质,而在真核线粒体中,H + 从膜间空间流向线粒体基质。 该分子机器各部分的转动通过氧化磷酸化从ADP和无机磷酸盐(P i)中再生 ATP,氧化磷酸化是产生 ATP 的第二种机制,可以收集存储在电化学梯度中的势能。
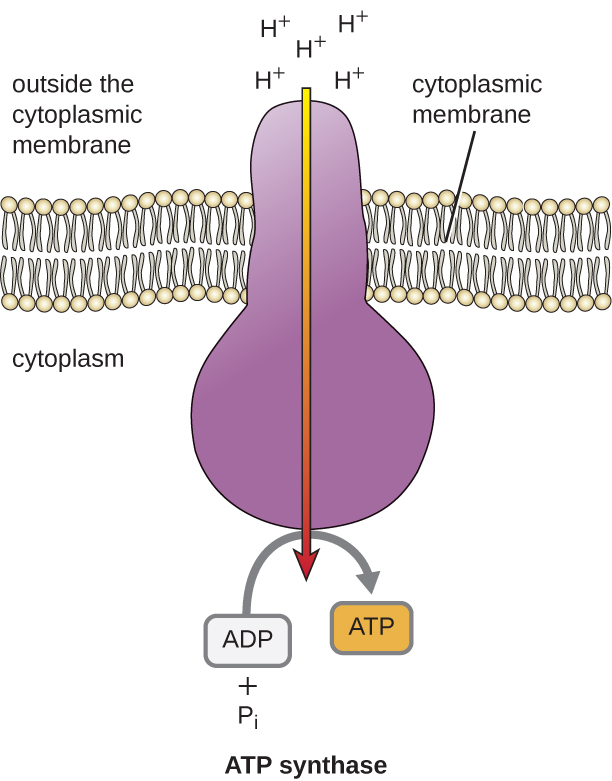
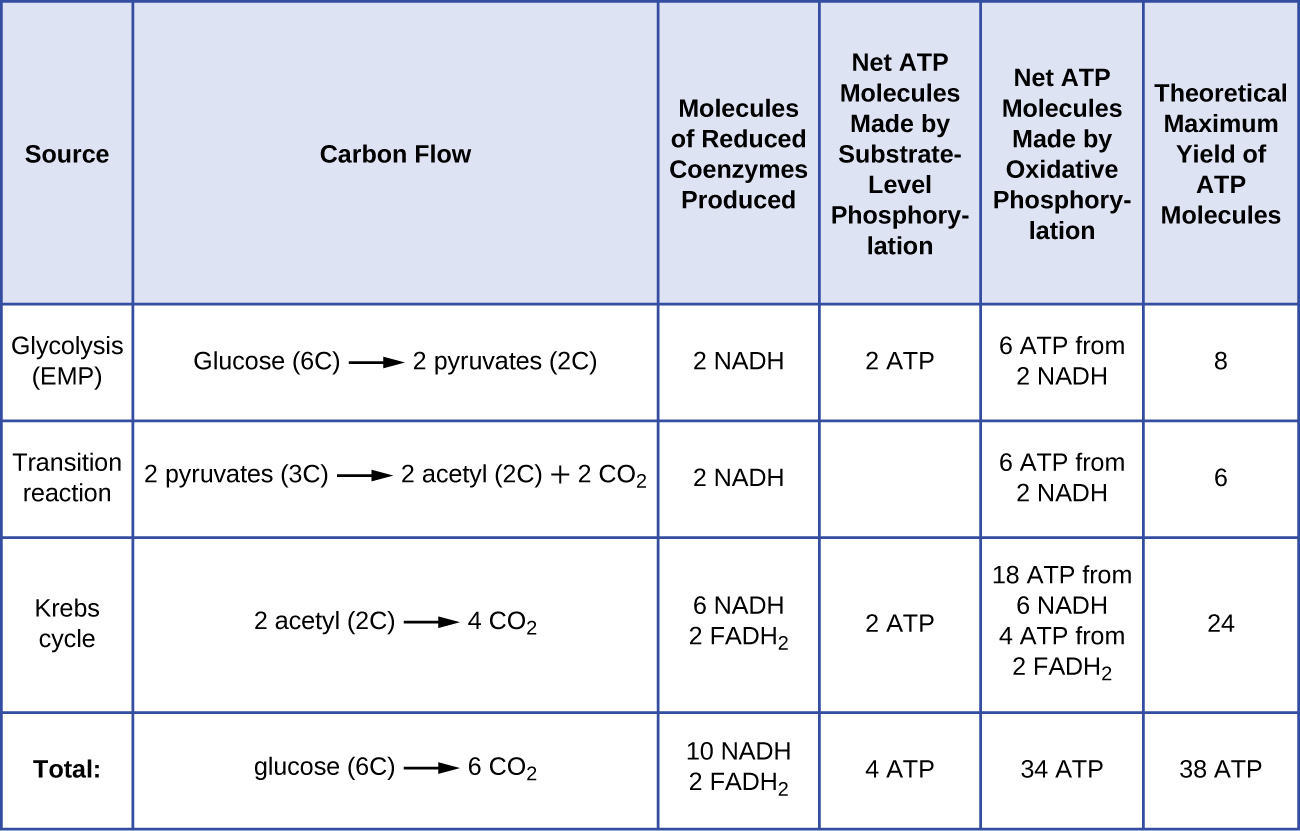
葡萄糖分解代谢产生的ATP分子的数量各不相同。 例如,电子传输系统复合物可以通过膜泵送的氢离子数量因生物种类的不同而有所不同。 在线粒体有氧呼吸中,电子从 NADH 的一个分子中通过会产生足够的质子动力,通过氧化磷酸化产生三个 ATP 分子,而电子从 FADH 2 的一个分子中通过会产生足够的质子动力,只能产生两个 ATP 分子ATP 分子。 因此,糖酵解、过渡反应和克雷布斯循环过程中每葡萄糖产生的 10 个 NADH 分子携带足够的能量来产生 30 个 ATP 分子,而在这些过程中每葡萄糖产生的两个 FADH 2 分子提供了足够的能量来制造四个 ATP 分子。 总体而言,葡萄糖完全有氧呼吸过程中产生的ATP的理论最大产量为38个分子,其中4个由底物级磷酸化产生,34个由氧化磷酸化产生(图\(\PageIndex{2}\))。 实际上,总ATP产量通常较低,从1到34个ATP分子不等,具体取决于细胞使用的是有氧呼吸还是厌氧呼吸;在真核细胞中,消耗一些能量将中间体从细胞质运送到线粒体中,从而影响ATP产量。
该图\(\PageIndex{2}\)总结了一个葡萄糖分子在完全有氧呼吸过程中各种过程产生的 ATP 的理论最大产量。
练习\(\PageIndex{1}\)
质子动力有什么作用?
摘要
- 葡萄糖细胞呼吸过程中产生的大多数ATP都是通过氧化磷酸化产生的。
- 电子传输系统 (ETS) 由一系列膜相关蛋白复合物和相关的移动辅助电子载流子组成。 ETS 嵌入原核生物的细胞质膜和真核生物的线粒体内部膜中。
- 每个 ETS 复合物都有不同的氧化还原电位,电子从氧化还原电位较为负的电子载流子转移到氧化还原电位较高的电子载体。
- 为了进行有氧呼吸,细胞需要氧气作为最终的电子受体。 细胞还需要完整的克雷布斯周期、适当的细胞色素氧化酶和氧气解毒酶,以防止有氧呼吸过程中产生的氧自由基的有害影响。
- 进行厌氧呼吸的生物使用替代电子传输系统载体将电子最终转移到最终的非氧电子受体。
- 微生物的电子传输系统的组成差异很大,可用于诊断目的,以帮助识别某些病原体。
- 当电子从 NADH 和 FADH 2 通过 ETS 传递时,电子会失去能量。 这种能量是通过将 H + 泵送到膜上来存储的,从而产生质子动力。
- 通过使用 ATP合成酶的化学渗透作用允许氢离子扩散回膜,可以利用这种质子动力的能量。 当氢离子通过其电化学梯度向下扩散时,ATP 合成酶的成分会旋转,从 ADP 中产生 ATP,通过氧化磷酸化产生 P i。
- 有氧呼吸在氧化磷酸化过程中形成的 ATP(最多 34 个 ATP 分子)比厌氧呼吸(介于 1 到 32 个 ATP 分子之间)多。