
46.2:能量流经生态系统
- Page ID
- 202879

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)学习目标
- 描述生物如何在食物网和相关的食物链中获取能量
- 解释营养等级之间能量转移的效率如何影响生态系统结构和动态
- 讨论营养等级以及如何使用生态金字塔对其进行建模
所有生物都需要这种或那种形式的能量。 大多数复杂的代谢途径(通常以三磷酸腺苷和三磷酸腺苷的形式)都需要能量,尤其是那些负责从较小的化合物中生成大分子的代谢途径,而生命本身就是一个能量驱动的过程。 如果没有恒定的能量输入,活生物体将无法从其单体亚基中组装大分子(蛋白质、脂质、核酸和复合碳水化合物)。
重要的是要了解生物是如何获得能量的,以及这种能量是如何通过食物网及其组成食物链从一个生物传递到另一个生物的。 食物网说明了能量是如何定向流经生态系统的,包括生物获取和使用能量的效率以及食物网中剩余的能量可供其他生物使用。
生物如何在食物网中获取能量
生物通过三种方式获得能量:光合作用、化学合作用以及异养生物消耗和消化其他活体或以前活着的生物。
光合和化合生物都分为一个称为自养的类别:能够合成自己的食物(更具体地说,能够使用无机碳作为碳源)的生物。 光合自养物(photoautotrophs)使用阳光作为能量来源,而化合自养物(chemoautotrophs)使用无机分子作为能量来源。 Autotrophs 对所有生态系统都至关重要。 没有这些生物,其他活生物就无法获得能量,生命本身也是不可能的。
光自养物,例如植物、藻类和光合细菌,是世界上大多数生态系统的能量来源。 这些生态系统通常用放牧的食物网来描述。 Photoautotrophs 利用太阳的太阳能,将其转化为 ATP(和 NADP)形式的化学能。 ATP 中储存的能量用于合成复杂的有机分子,例如葡萄糖。
Chemoautotrophs 主要是在没有阳光的稀有生态系统中发现的细菌,例如与黑暗洞穴或海底热液喷口相关的生态系统(图\(\PageIndex{1}\))。 热液喷口中的许多化学自养生物使用从喷口释放的硫化氢(H 2 S)作为化学能源。 这使chemoautotrophs能够合成复杂的有机分子,例如葡萄糖,以获取自身的能量,进而为生态系统的其余部分提供能量。

在营养水平之内的生产力
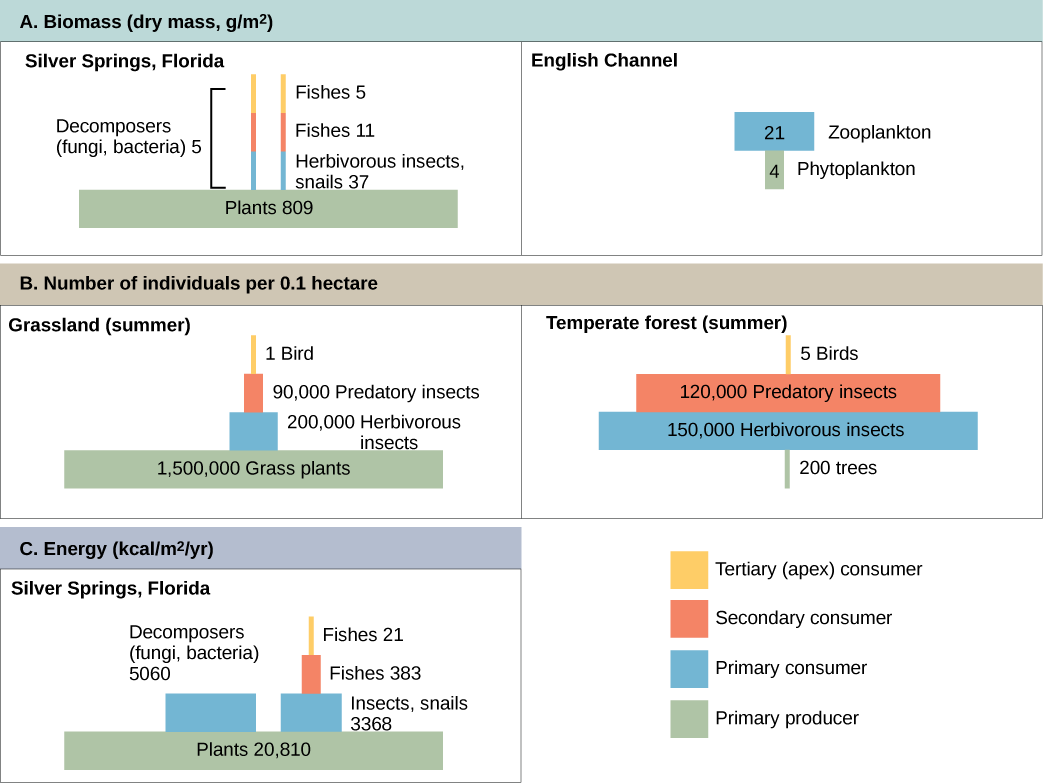
生态系统内的生产力可以定义为进入生态系统的能量在特定营养层面上融入生物质的百分比。 生物量是指在测量时单位面积内在营养层面上活体或先前活过的生物体的总质量。 生态系统在每个营养层面都有典型的生物量。 例如,在英吉利海峡生态系统中,初级生产者的生物量为4 g/m 2(克/平方米),而主要消费者的生物量为21 g/m 2。
初级生产者的生产力在任何生态系统中都尤为重要,因为这些生物通过光自养或化学自养为其他活生物体带来能量。 光合初级生产者吸收来自太阳的能量的速度称为总初级生产力。 Silver Springs 水生生态系统内能量流的分隔图显示了初级生产力总值的示例,如图所示(图 46.1.7)。 在这个生态系统中,初级生产者累积的总能量(初级生产力总值)为20,810 kcal/m 2 /年。
由于所有生物都需要将其中的一部分能量用于其自身功能(例如呼吸和由此产生的代谢热损失),因此科学家们通常指的是生态系统的净初级生产力。 净初级生产力是指初级生产者在考虑了生物的呼吸和热量损失后剩余的能量。 然后,净生产率就可以在下一个营养层面上提供给主要消费者。 在我们的 Silver Spring 示例中,20,810 kcal/m 2 /yr 中的 13,187 用于呼吸或作为热量流失,剩下 7,632 kcal/m 2 /yr 的能量供主要消费者使用。
生态效率:营养等级之间的能量转移
如图 46.1.7 所示,随着能量从初级生产者流向消费者和分解者的各个营养级别,大量能量从一个营养层流向下一个营养级别。 造成这种损失的主要原因是热力学第二定律,该定律指出,每当能量从一种形式转化为另一种形式时,系统中就会有混乱(熵)的趋势。 在生物系统中,这意味着当来自一个营养层面的生物消耗下一个等级时,大量的能量会作为代谢热量流失。 在 Silver Springs 生态系统示例(图 46.1.7)中,我们看到,初级消费者每年可从初级生产者那里获得的 7618 kcal/m 2 /y 的能量中生产了 1103 kcal/m 2 /y。 两个连续营养等级之间的能量传递效率的测量称为营养级传递效率(TLTE),由以下公式定义:
\[\text{TLTE} = \frac{\text{production at present trophic level}}{\text{production at previous trophic level}} * 100 \nonumber\]
在 Silver Springs,前两个营养等级之间的TLTE约为14.8%。 营养级之间的能量转移效率低通常是限制食物网中观察到的食物链长度的主要因素。 事实是,经过四到六次能量转移后,剩下的能量不足以支持另一个营养等级。 在图 46.1.5 所示的安大略湖示例中,初级生产者(绿藻)和主要消费者(奇努克鲑鱼)之间仅发生了三次能量转移。
生态学家有许多不同的方法来测量生态系统内的能量传输。 有些转移更容易或更难测量,这取决于生态系统的复杂性以及科学家观察生态系统的途径。 换句话说,有些生态系统比其他生态系统更难研究,有时必须对能量转移的量化进行估计。
在表征生态系统内能量流方面,另一个重要参数是净生产效率。 净生产效率(NPE)使生态学家能够量化特定营养水平的生物将其获得的能量转化为生物质的效率;它是使用以下公式计算的:
\[\text{NPE} = \frac{\text{net consumer productivity}}{\text{assimilation}} * 100 \nonumber\]
净消费者生产率是指下一个营养级生物可获得的能量含量。 同化是指当前营养水平的生物质(每单位面积产生的能量含量),其中考虑了由于食物摄入不完全导致的能量损失、用于呼吸的能量以及作为废物而损失的能量。 不完全摄取是指一些消费者只吃一部分食物的事实。 例如,当狮子杀死羚羊时,它会吃掉除皮革和骨头之外的所有东西。 狮子缺少骨内富含能量的骨髓,因此狮子没有利用猎物所能提供的所有卡路里。
因此,NPE衡量每个营养等级使用食物中的能量并将其转化为生物质以为下一个营养等级提供燃料的效率。 一般而言,冷血动物(外热),例如无脊椎动物、鱼类、两栖动物和爬行动物,它们为呼吸和加热消耗的能量少于温血动物(吸热),例如鸟类和哺乳动物。 吸热产生的额外热量虽然对于这些生物在寒冷环境中的活性而言是一种优势,但就NPE而言,这是一个主要的缺点。 因此,许多吸热体必须比外温更频繁地进食才能获得生存所需的能量。 一般而言,外热的 NPE 比吸热的 NPE 高出一个数量级(10 倍)。 例如,据测量,毛毛虫吃树叶的NPE为18%,而松鼠吃橡子的NPE可能低至1.6%。
温血动物的能量利用效率低下对世界食物供应产生了广泛影响。 人们普遍认为,肉类行业使用大量农作物来喂养牲畜,而且由于NPE很低,动物饲料产生的大部分能量都流失了。 例如,生产1000卡路里(kcal)的玉米或大豆的膳食卡路里(kcal)的成本约为1美分,但生产类似数量的卡路里种植牛以供食用牛肉的成本约为0.19美元。 牛奶中同样的能量含量也很昂贵,约为每 1000 千卡0.16美元。 这种差异在很大程度上是由于牛的NPE较低。 因此,全世界越来越多地推动非肉类和非乳制品食品的消费,从而减少为肉类行业喂养动物所浪费的能源。
建模生态系统能量流:生态金字塔
生态系统的结构可以通过生态金字塔进行可视化,查尔斯·埃尔顿在20世纪20年代的开创性研究中首次描述了生态金字塔。 生态金字塔显示了各个营养层面上各种参数(例如生物数量、能量和生物量)的相对数量。
数字金字塔可以是直立的也可以是倒置的,具体取决于生态系统。 如图所示\(\PageIndex{2}\),夏季的典型草原以许多植物为基础,每个营养级的生物数量都会减少。 但是,在夏季,在温带森林中,与主要消费者的数量(主要是昆虫)相比,金字塔底部的树木很少。 因为树木很大,所以它们具有很强的光合能力,并在这个生态系统中的其他植物中占主导地位以获得阳光。 即使数量较少,森林的初级生产者仍然能够支持其他营养级别。
另一种可视化生态系统结构的方法是使用生物质金字塔。 这座金字塔测量在不同营养等级转化为活组织的能量。 以 Silver Springs 生态系统为例,此数据显示的是直立的生物质金字塔(图\(\PageIndex{2}\)),而英吉利海峡示例中的金字塔是倒置的。 银泉生态系统的植物(初级生产者)占那里发现的生物量的很大一部分。 但是,英吉利海峡例子中的浮游植物所占的生物量少于主要消费者,即浮游动物。 与倒置的数字金字塔一样,这座倒金字塔不是因为初级生产者缺乏生产力,而是浮游植物的高周转率造成的。 主要消费者会迅速消耗浮游植物,从而最大限度地减少其在任何特定时间点的生物量。 但是,浮游植物繁殖速度很快,因此它们能够支持生态系统的其余部分。
金字塔生态系统建模还可用于显示通过营养等级的能量流。 请注意,这些数字与图 46.1.7 中能量流室图中使用的数字相同。 能量金字塔总是直立的,没有足够初级生产力的生态系统是无法支持的。 所有类型的生态金字塔都可用于表征生态系统结构。 但是,在对生态系统能量流的研究中,能量金字塔是生态系统结构中最一致和最具代表性的模型(图\(\PageIndex{2}\))。
练习
描绘生物或生物量数量的金字塔可以是倒置的、直立的,甚至是菱形的。 但是,能量金字塔总是直立的。 为什么?
- 回答
-
生物金字塔可能是倒置的或菱形的,因为大型生物,例如树,可以维持许多较小的生物。 同样,生物的低生物量可以在下一个营养层面维持更大的生物量,因为生物繁殖速度快,从而提供持续的营养。 但是,由于热力学定律的原因,能量金字塔必须始终是直立的。 热力学第一定律指出,能量既不能产生也不能被摧毁;因此,每个营养等级都必须从下面的营养等级获得能量。 热力学第二定律指出,在能量传递过程中,一些能量总是作为热量流失;因此,每一个较高的营养等级可用的能量就会减少。
食物网的后果:生物放大倍率
生态系统动态最重要的环境后果之一是生物放大作用。 生物放大作用是指从初级生产者到主要消费者的各个营养级生物体内的持久性有毒物质浓度不断增加。 许多物质已被证明具有生物蓄积性,包括对杀虫剂 d ichloro d ibeny t richlorethane(DD T)的经典研究,该研究发表在雷切尔·卡森(Rachel Carson)的1960年代畅销书《寂静的春天》中。 滴滴涕在危害为人所知之前是一种常用的杀虫剂。 在一些水生生态系统中,每个营养级的生物消耗了许多较低水平的生物,这导致食用鱼类的鸟类(顶点消费者)中的滴滴涕增加。 因此,鸟类积累了足够数量的滴滴涕,使其蛋壳变得脆弱。 这种效应增加了筑巢期间的卵子破损率,并被证明对这些鸟类种群有不利影响。 1970年代,美国禁止使用滴滴涕。
其他具有生物放大作用的物质包括多氯联苯(PCB)和重金属,如汞、铅和镉,前者在美国一直用于冷却液中,直至1979年被禁止使用。 这些物质最好在水生生态系统中进行研究,在水生生态系统中,处于不同营养水平的鱼类会积聚初级生产者带入生态系统的有毒物质。 正如美国国家海洋和大气管理局(NOAA)在休伦湖萨吉诺湾进行的一项研究(图\(\PageIndex{3}\))所示,从生态系统的初级生产者(浮游植物)到鱼类物种的营养水平不同,多氯联苯的浓度有所增加。 主要消费者(角膜白斑)的多氯联苯含量是浮游植物的四倍多。 此外,根据其他研究的结果,食用这些鱼的鸟类的多氯联苯含量可能比湖鱼中发现的多氯联苯水平至少高出一个数量级。
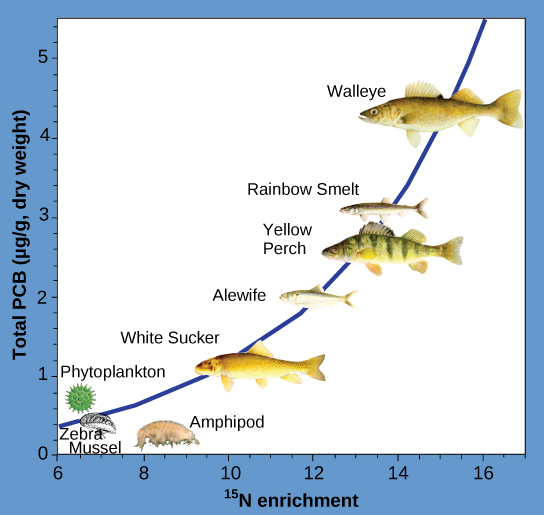
某些类型的海产品中汞和镉等重金属的积累也引起了其他担忧。 美国环境保护署(EPA)建议,孕妇和幼儿不应食用任何剑鱼、鲨鱼、鲭鱼或方头鱼,因为它们的汞含量很高。 建议这些人吃汞含量低的鱼:鲑鱼、罗非鱼、虾、鳕鱼和鲶鱼。 生物放大作用是一个很好的例子,说明生态系统动态如何影响我们的日常生活,甚至影响我们所吃的食物。
摘要
生态系统中的生物通过多种方式获取能量,当能量从食物网的底部流向顶部时,能量会在营养层之间转移,每次转移时能量都会流失。 这些转移的效率对于理解温血动物和冷血动物的不同行为和饮食习惯非常重要。 生态系统能量建模最好使用生态能量金字塔来完成,尽管其他生态金字塔提供了有关生态系统结构的其他重要信息。
词汇表
- 同化
- 考虑到由于食物摄入不完全而损失的能量、用于呼吸的能量以及作为废物的能量损失后,生物质从以前的营养水平消耗和吸收
- 生物放大作用
- 从初级生产者到主要消费者,每个营养级生物体内持久性有毒物质的浓度都在增加
- 有机燃料
- 在测量时,单位面积内营养级内活体或以前活过的生物体的总重量
- chemoautotroph
- 生物能够利用来自无机分子的能量合成自己的食物
- 生态金字塔
- (也是 Eltonian Pyramid)根据生物数量、生物量或能量含量以图形表示生态系统中不同营养等级
- 初级生产力总值
- 光合初级生产者吸收来自太阳的能量的速率
- 净消费者生产率
- 下一个营养级生物可获得的能量含量
- 净初级生产率
- 考虑到生物的呼吸和热量损失后仍留在初级生产者中的能量
- 净生产效率 (NPE)
- 衡量营养等级将其从前一个营养等级获得的能量转化为生物质的能力
- 营养等级转移效率 (TLTE)
- 两个连续营养等级之间的能量传递效率