
19.2: 种群遗传学
- Page ID
- 202255

培养技能
- 描述总体中不同类型的变异
- 解释为什么自然选择只能对可遗传的变异起作用
- 描述遗传漂移和瓶颈效应
- 解释每种进化力如何影响种群的等位基因频率
群体中的个体通常表现出不同的表型,或者表达特定基因的不同等位基因,称为多态性。 具有两种或两种以上特定特征变体的种群被称为多态性。 表型在个体之间的分布(称为种群变异)受多种因素的影响,包括种群的遗传结构和环境(图\(\PageIndex{1}\))。 了解人群表型变异的来源对于确定种群将如何应对不同的进化压力进化非常重要。

遗传差异
自然选择和其他一些进化力量只能作用于可遗传的特征,即生物体的遗传密码。 由于等位基因是从父母传给后代的,因此可以选择那些具有有益特征或行为的等位基因,而有害的等位基因可能会被选中。 获得的特征在大多数情况下是不可遗传的。 例如,如果运动员每天在健身房锻炼身体,增强肌肉力量,那么运动员的后代不一定会长大后成为健美运动员。 另一方面,如果快速跑步的能力有遗传基础,则这种能力可能会传给孩子。
遗传力是表型变异中可归因于人群中个体之间的遗传差异或遗传差异的部分。 种群表型变异的可遗传性越大,它就越容易受到作用于可遗传变异的进化力量的影响。
人群中等位基因和基因型的多样性称为遗传变异。 当科学家参与物种的繁殖时,例如在动物园和自然保护区与动物一起繁殖,他们会努力增加种群的遗传差异,以尽可能多地保护表型多样性。 这还有助于降低与近亲繁殖相关的风险,近亲繁殖是密切相关个体的交配,这可能会产生不良影响,将可能导致异常和易患疾病的有害隐性突变聚在一起。 例如,一种由罕见的隐性等位基因引起的疾病可能存在于人群中,但只有当一个人携带两个等位基因拷贝时,它才会显现出来。 由于该等位基因在栖息地不受限制的正常、健康人群中很少见,因此两个携带者交配的机会很低,即便如此,他们的后代中也只有 25% 会从父母双方那里继承该疾病等位基因。 尽管它可能在某个时候发生,但这种情况发生的频率还不够高,自然选择无法迅速从人群中消灭等位基因,因此,基因库中的等位基因将保持在较低水平。 但是,如果一个携带者家族开始相互杂交,这将大大增加两个携带者交配并最终产生患病后代的可能性,这种现象被称为近亲繁殖抑郁症。
在人群中识别出的等位基因频率的变化可以揭示其演变情况。 除了自然选择外,还有其他进化力量可能在起作用:遗传漂移、基因流动、突变、非随机交配和环境差异。
遗传漂移
自然选择理论源于这样一种观察,即人群中的一些人比其他人更有可能存活更长时间,有更多的后代;因此,他们将把更多的基因传给下一代。 例如,一只大而强大的雄性大猩猩比一只更小、更弱的雄性大猩猩更有可能成为种群中的银背,而群体的交配量远远超过该群体中的其他雄性。 背包领袖将生下更多的后代,这些后代共享他一半的基因,并且很可能像他们的父亲一样变得更大更强壮。 随着时间的推移,较大体积的基因在人群中的频率将增加,因此,平均而言,种群将增加。 也就是说,如果这种特殊的选择压力或驱动选择性力量是唯一对人群起作用的压力,就会发生这种情况。 在其他例子中,更好的伪装或更强的抗旱能力可能会带来选择压力。
人群的等位基因和基因型频率可以改变的另一种方式是遗传漂移(图\(\PageIndex{2}\)),这只是偶然的影响。 偶然地,有些人的后代会比其他人多 —— 不是因为某种遗传编码特征所带来的优势,而仅仅是因为一只雄性碰巧在正确的时间出现在正确的地方(当接受的雌性走过时)或者因为另一只雄性碰巧在错误的地方错误的时间(狐狸狩猎的时候)。

练习\(\PageIndex{1}\)
你认为基因漂移会在岛屿或大陆上更快地发生吗?
- 回答
-
在预计人口较少的岛屿上,遗传漂移的发生速度可能会更快。
少数群体更容易受到遗传漂移的影响。 另一方面,大量人口会受到偶然影响的缓冲。 如果10个人中的一个人碰巧在将任何后代留给下一代之前在年轻时死亡,那么其所有基因(占人群基因库的1/10)都将突然消失。 在100人群中,这仅占总基因库的1%;因此,它对人群遗传结构的影响要小得多。
自然事件也可以放大遗传漂移,例如自然灾害随机造成很大一部分人口死亡。 它被称为瓶颈效应,它导致很大一部分基因突然被消灭(图\(\PageIndex{3}\))。 幸存者的遗传结构一举变成了整个人群的遗传结构,这可能与灾前人口大不相同。
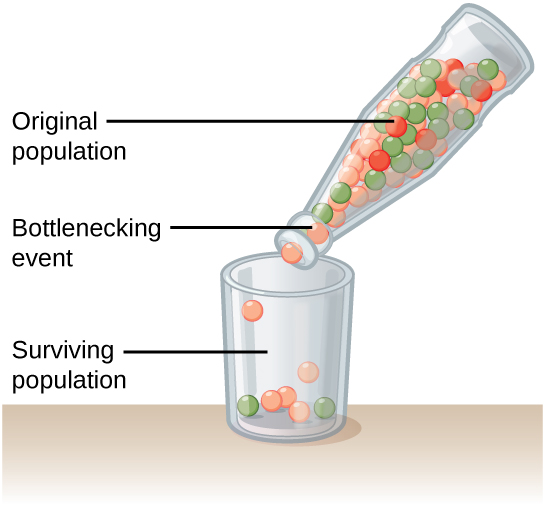
种群可能会受到遗传漂移强烈影响的另一种情景是,如果一部分人口离开新地点开始新的种群,或者如果一个种群被某种物理屏障分开。 在这种情况下,这些人不太可能代表整个人口,从而产生创始效应。 当遗传结构发生变化以匹配新人群开国元勋和母亲的遗传结构时,就会产生创始人效应。 创始人效应被认为是南非荷兰定居者南非荷兰语群体遗传史中的关键因素,南非荷兰语中常见但在大多数其他人群中很少见的突变就证明了这一点。 这可能是因为创始殖民者携带这些突变的比例高于正常水平。 因此,该人群的亨廷顿氏病(HD)和范可尼贫血(FA)发病率异常高,这是一种已知会导致血髓和先天性异常(甚至癌症)的遗传性疾病。 1
链接到学习
观看这段简短的视频,详细了解创始人和瓶颈效应。
科学方法连接:测试瓶颈效应
问题:自然灾害如何影响人口的遗传结构?
背景:当大部分人口突然被地震或飓风摧毁时,在事件中幸存下来的个人通常是原始群体的随机抽样。 因此,人群的基因构成可能会发生巨大变化。 这种现象被称为瓶颈效应。
假设:反复发生的自然灾害将产生不同的种群遗传结构;因此,每次进行此实验时,结果都会有所不同。
检验假设:使用不同颜色的珠子计算出原始种群。 例如,红色、蓝色和黄色的珠子可能代表红色、蓝色和黄色的个体。 在记录了原始种群中每个人的数量后,将它们全部放入脖子较窄的瓶子中,一次只能容纳几颗珠子。 然后,将瓶中三分之一的内容物倒入碗中。 这代表了在自然灾害造成大多数人口死亡之后的幸存者。 计算碗中不同颜色珠子的数量,并记录下来。 然后,将所有珠子放回瓶中,再重复四次实验。
分析数据:比较实验得出的五个种群。 种群中是否都包含相同数量的不同颜色的珠子,还是各不相同? 请记住,这些种群都来自完全相同的父母群体。
得出结论:由此产生的五个种群很可能会有很大的不同。 这是因为自然灾害不是选择性的,它们会随机杀死和拯救个人。 现在想想这会如何影响实际人口。 飓风袭击密西西比州墨西哥湾沿岸时会发生什么? 生活在海滩上的海鸟怎么样?
基因流
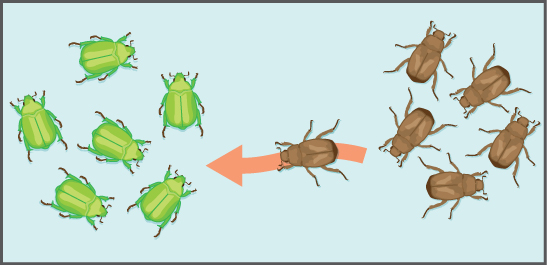
另一个重要的进化力量是基因流:由于个体或配子的迁移,等位基因流入和流出人群(图\(\PageIndex{4}\))。 虽然有些人口相当稳定,但另一些人口的变化更大。 例如,许多植物通过风或鸟将花粉送到很远的地方,以便在一定距离之外为同一物种的其他种群授粉。 即使是最初看似稳定的种群,例如狮子的骄傲,也可以经历相当一部分的移民和移民,因为发育中的雄性离开母亲,与遗传无关的雌性寻找新的自豪感。 这种个体进出群体的可变流动不仅改变了种群的基因结构,而且还可能为不同地质位置和栖息地的种群带来新的遗传变异。
突变
突变是生物体DNA的改变,是种群多样性的重要驱动力。 物种的进化是由于随着时间的推移而发生的突变的积累。 新突变的出现是引入新的基因型和表型变异的最常见方式。 有些突变是不利或有害的,通过自然选择很快就会从人群中消除。 其他则是有益的,将在人群中传播。 突变是有益还是有害取决于它能否帮助生物存活到性成熟和繁殖。 有些突变没有任何作用,可以在基因组中持续存在,不受自然选择的影响。 有些会对基因和由此产生的表型产生巨大影响。
非随机交配
如果个体与同龄人非随机交配,则结果可能是人口变化。 发生非随机交配的原因有很多。 原因之一是简单的伴侣选择;例如,雌性豌豆可能更喜欢尾巴更大、更亮的孔雀。 自然选择会选择能为个人带来更多交配的特征。 一种常见的配偶选择形式,称为分类交配,是个人倾向于与表型上与自己相似的伴侣交配。
非随机交配的另一个原因是物理位置。 在分布在遥远地理距离的大量人口中尤其如此,因为并非所有人都有平等的机会彼此接触。 有些人可能相隔数英里穿过树林或崎岖的地形,而另一些人则可能住在附近。
环境差异
基因并不是决定种群变异的唯一参与者。 表型还受到其他因素的影响,例如环境(图\(\PageIndex{5}\))。 例如,由于经常暴露在阳光下,这是一种环境因素,海滩游客的皮肤可能比城市居民更黑。 一些主要特征,例如性别,是由某些物种的环境决定的。 例如,一些海龟和其他爬行动物具有取决于温度的性别决定(TSD)。 TSD 意味着,如果个体的卵子在一定的温度范围内孵育,则会发育成雄性,或者在不同的温度范围内孵育雌性。

人群之间的地理分离可能导致这些人群之间表型变异的差异。 大多数人群之间可以看到这种地理差异,并且可能很明显。 一种叫做 cline 的地理变异可以看作是给定物种的种群在生态梯度上逐渐变化。 例如,在更靠近地球两极的凉爽气候中,温血动物的身体往往更大,这使它们能够更好地节约热量。 这被认为是纬度斜线。 或者,开花植物往往在不同的时间开花,具体取决于它们在被称为海拔斜坡的山坡上的位置。
如果人群之间存在基因流动,则个体在临床上表现出逐渐的表型差异。 另一方面,受限的基因流动可能导致突然的差异,甚至物种形成。
摘要
遗传和环境因素都可能导致人群的表型变异。 不同的等位基因可以赋予不同的表型,不同的环境也可能导致个体的外观或行为有所不同。 但是,只有那些编码在个体基因中的差异才能传递给其后代,从而成为自然选择的目标。 自然选择的工作原理是选择具有有益特征或行为的等位基因,而选择具有有害特质的等位基因。 遗传漂移源于生殖系中某些个体可能比其他人有更多的后代。 当个体离开或加入人群时,等位基因频率可能会因基因流动而发生变化。 个体DNA的突变可能会给群体带来新的变异。 当个体不与组中的其他人随机交配时,等位基因频率也可以改变。
脚注
- 1 A.J. Tipping 等人,“南非荷兰语人口范可尼贫血家族产生创始效应的分子和家谱证据”,PNAS 98,第 10 期(2001):5734-5739,doi:10.1073/pnas.091402398。
词汇表
- 分类交配
- 当个人倾向于与表型上与自己相似的人交配时
- 瓶颈效应
- 放大自然事件或灾难造成的遗传漂移
- cline
- 生态梯度上的逐渐地理差异
- 基因流
- 由于个体或配子的迁移,等位基因流入和流出种群
- 遗传漂移
- 机会对人群基因库的影响
- 遗传差异
- 人群中等位基因和基因型的多样性
- 地理差异
- 地理上分离的人群之间表型变异的差异
- 遗传力
- 可归因于其遗传差异的种群变异比例
- 近亲繁殖
- 密切相关的人交配
- 近亲繁殖抑郁症
- 近亲繁殖种群的异常和疾病增加
- 非随机交配
- 由于伴侣选择或其他力量导致个体比其他人更容易与某些表型交配,人群的基因库发生变化
- 种群变异
- 人群中表型的分布
- 选择性压力
- 导致一种表型优于另一种表型的环境因素