
19.1: 人口演变
- Page ID
- 202219

培养技能
- 定义种群遗传学并描述在研究种群进化时如何使用群体遗传学
- 定义哈迪-温伯格原理并讨论其重要性
在查尔斯·达尔文和阿尔弗雷德·罗素·华莱士发展自然选择理念时,人们还不了解遗传机制或遗传学。 这种缺乏理解是理解进化许多方面的绊脚石。 事实上,当时占主导地位(且不正确的)遗传理论,即混合遗传,很难理解自然选择会如何运作。 达尔文和华莱士没有意识到奥地利僧侣格雷戈尔·孟德尔的遗传学著作,该著作于1866年出版,也就是达尔文的著作《物种起源》出版后不久。 孟德尔的作品在二十世纪初被重新发现,当时遗传学家迅速了解了继承的基础知识。 最初,新发现的基因颗粒性质使生物学家难以理解逐渐进化是如何发生的。 但是在接下来的几十年中,遗传学和进化被整合到所谓的现代合成中,即对自然选择与遗传学之间关系的连贯理解,这种理解在20世纪40年代形成,今天被普遍接受。 总而言之,现代综合描述了自然选择等进化过程如何影响种群的基因构成,进而描述了这如何导致种群和物种的逐渐进化。 该理论还将种群随时间推移而发生的变化(称为微进化)与产生具有巨大差异特征的新物种和更高分类组(称为宏观进化)的过程联系起来。
日常联系:进化与流感疫苗
每年秋天,媒体都会开始报道流感疫苗接种和潜在疫情。 科学家、健康专家和机构确定针对不同人口群体的建议,预测最佳生产和接种时间表,研制疫苗,并设立诊所提供疫苗接种。 你可能会认为一年一度的流感疫苗是媒体的大量炒作,一种重要的健康保护,或者只是手臂上短暂的不舒服刺痛。 但是你从进化的角度来看待它吗?
媒体对年度流感疫苗的炒作是基于我们对进化的理解。 每年,全球科学家都在努力预测他们预计来年最广泛和最有害的流感菌株。 这些知识基于流感菌株随着时间的推移以及过去几个流感季节的演变情况。 然后,科学家们努力研制出最有效的疫苗来对抗这些选定的菌株。 在短时间内生产了数亿剂疫苗,以便在最佳时间为关键人群提供疫苗接种。
由于像流感这样的病毒进化速度非常快(尤其是在进化时期),这构成了很大的挑战。 病毒的变异和复制速度很快,因此为预防去年的流感菌株而开发的疫苗可能无法提供抵御来年流感菌株所需的保护。 这些病毒的进化意味着持续适应以确保存活,包括适应以往疫苗存活下来。
种群遗传学
回想一下,特定角色的基因可能有多个等位基因或变体,它们编码与该角色相关的不同特征。 例如,在人类的ABO血型系统中,三个等位基因决定红细胞表面的特定血型蛋白。 二倍体生物群体中的每个人只能携带特定基因的两个等位基因,但构成该群体的个体中可能存在两个以上的等位基因。 孟德尔跟随等位基因,因为它们是从父母遗传给后代的。 二十世纪初,被称为群体遗传学的研究领域的生物学家开始研究选择性力量如何通过等位基因和基因型频率的变化改变人群。
等位基因频率(或基因频率)是特定等位基因在群体中出现的速率。 到目前为止,我们将进化作为生物群体特征的变化来讨论,但表型变化的背后是遗传变化。 在种群遗传学中,“进化” 一词被定义为群体中等位基因频率的变化。 以 ABO 血型系统为例,其中一个等位基因 I A 的频率是该等位基因的拷贝数除以群体中 ABO 基因的所有拷贝数。 例如,约旦 1 的一项研究发现 I A 的频率为 26.1%。 I B 和 I 0 等位基因分别占等位基因的 13.4% 和 60.5%,所有频率加起来达到 100%。 随着时间的推移,这种频率的变化将构成人口的演变。
给定种群中的等位基因频率可能因环境因素而变化;因此,在自然选择过程中,某些等位基因比其他等位基因更为广泛。 自然选择可以改变种群的基因构成;例如,如果给定的等位基因赋予了允许个体更好地存活或拥有更多后代的表型。 由于这些后代中有许多也会携带有益的等位基因,通常还有相应的表型,因此它们会有更多的自己的后代也携带等位基因,从而延续周期。 随着时间的推移,等位基因将在人群中传播。 一些等位基因将很快以这种方式固定,这意味着人群中的每个人都将携带该等位基因,而如果来自基因库中的显性等位基因,则有害的突变可能会很快被消除。 基因库是一个群体中所有等位基因的总和。
有时,群体中的等位基因频率会随机变化,与现有等位基因频率相比,对群体没有任何好处。 这种现象被称为遗传漂移。 自然选择和遗传漂移通常在人群中同时发生,不是孤立的事件。 很难确定哪个过程占主导地位,因为通常几乎不可能确定每次发生时等位基因频率变化的原因。 在人群中孤立部分引发等位基因频率变化的事件称为创始人效应,这在原始人群中并不典型。 自然选择、随机漂移和创始人效应可能导致人群基因组的重大变化。
哈迪-温伯格平衡原理
二十世纪初,英国数学家戈弗雷·哈迪(Godfrey Hardy)和德国医生威廉·温伯格(Wilhelm Weinberg)提出了描述人群基因构成的平衡原理。 该理论后来被称为哈迪-温伯格平衡原理,它指出,种群的等位基因和基因型频率本质上是稳定的,除非某种进化力量作用于种群,否则等位基因和基因型频率都不会改变。 Hardy-Weinberg原理假设条件没有突变、迁移、移民或支持或反对基因型的选择性压力,再加上无限种群;尽管没有人群能够满足这些条件,但该原理为比较实际人口变化提供了一个有用的模型。
根据这一理论,群体遗传学家在其数学模型中将不同的等位基因表示为不同的变量。 例如,变量 p 通常代表特定等位基因的频率,例如 Y 代表孟德尔豌豆中的黄色特征,而变量 q 表示赋予绿色的 y 等位基因的频率。 如果这些是群体中给定轨迹的仅有的两个可能的等位基因,则 p + q = 1。 换句话说,所有 p 等位基因和所有 q 等位基因构成了在种群中发现的该轨迹的所有等位基因。
但是,大多数生物学家最终感兴趣的不是不同等位基因的频率,而是由此产生的基因型的频率,即种群的遗传结构,科学家可以从中推测表型的分布。 如果观察到表型,则只能知道纯合隐性等位基因的基因型;计算结果提供了剩余基因型的估计值。 由于每个人每个基因携带两个等位基因,如果已知等位基因频率(p 和 q),预测这些基因型的频率是一种简单的数学计算,可以确定如果从基因库中随机抽取两个等位基因,则获得这些基因型的概率。 因此,在上述情景中,单个豌豆植物可能是 pp (YY),从而生产黄豌豆;pq (Yy),也是黄豌豆;或 qq (yy),从而生产青豆(图\(\PageIndex{1}\))。 换句话说,pp 个体的频率仅为 p 2;pq 个体的频率为 2pq;而 qq 个体的频率为 q 2。 而且,再说一遍,如果 p 和 q 是人群中给定特征的仅有的两个可能的等位基因,则这些基因型频率总和为一:p 2 + 2pq + q 2 = 1。
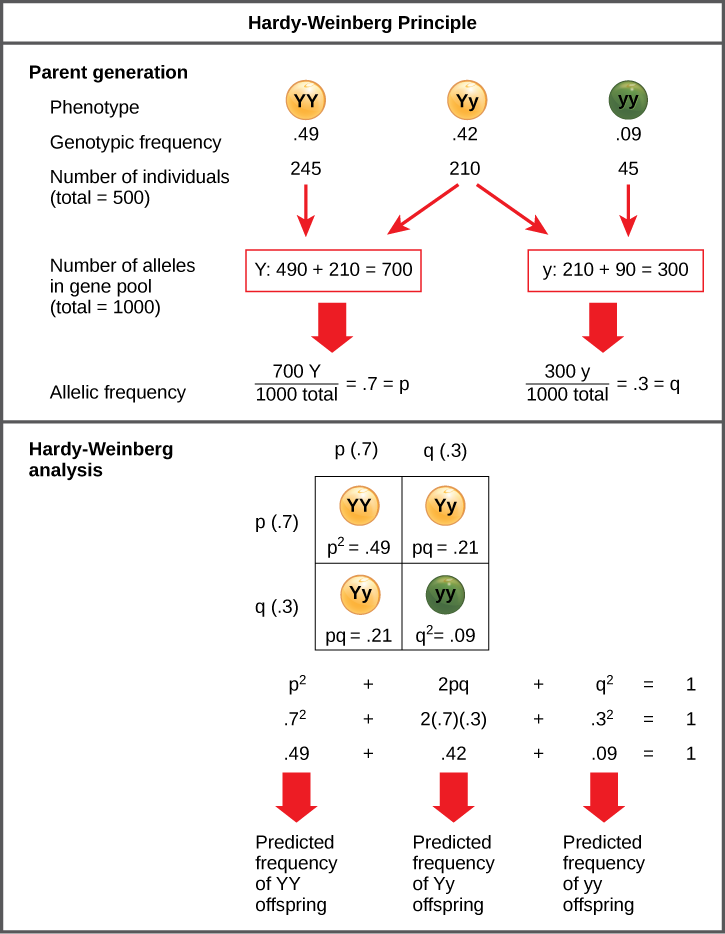
练习\(\PageIndex{1}\)
在植物中,紫罗兰花的颜色(V)比白色(v)占主导地位。 如果在 500 株植物种群中 p = 0.8 且 q = 0.2,那么你预计有多少个体会成为纯合显性 (VV)、杂合子 (Vv) 和纯合隐性遗传 (vv)? 你预计有多少植物会有紫罗兰花,有多少会有白花?
- 回答
-
预计配电量为 320 VV、160VV 和 20 VV 电厂。 具有VV或Vv基因型的植物将有紫罗兰花,而具有vv基因型的植物将有白花,因此预计总共有480株植物会有紫罗兰花,20株植物将有白花。
从理论上讲,如果一个种群处于平衡状态,也就是说,没有进化力量作用于它,那么一代又一代地将具有相同的基因库和遗传结构,而这些方程式将一直成立。 当然,即使是哈迪和温伯格也认识到,没有自然种群能够幸免于进化。 由于漂移、突变、可能的迁移和选择,自然界种群的基因构成不断变化。 因此,确定人群中表型的确切分布的唯一方法就是出去计算表型。 但是哈迪-温伯格原理为科学家提供了非进化人口的数学基线,他们可以将不断变化的人群与之进行比较,从而推断出哪些进化力量可能在起作用。 如果等位基因或基因型的频率偏离了哈迪-温伯格方程的预期值,那么种群正在演变。
摘要
进化论的现代综合源于达尔文、华莱士和孟德尔关于进化和遗传的思想的凝聚力,以及对种群遗传学的更现代的研究。 它描述了种群和物种的演变,从个体之间的小规模变化到古生物学时期的大规模变化。 为了了解生物是如何进化的,科学家可以追踪种群在一段时间内的等位基因频率。 如果它们代代相传,科学家们可以得出结论,人口不在哈迪-温伯格均衡状态,因此正在演变。
脚注
- 1 Sahar S. Hanania、Dhia S. Hassawi 和 Nidal M. Irshaid,“约旦人群中 ABO 血型系统的等位基因频率和分子基因型”,《医学科学杂志》 7(2007):51-58,doi: 10.3923/jms.2007.51.58。
词汇表
- 等位基因频率
- (也称基因频率)特定等位基因在人群中出现的速率
- 创始者效应
- 引发部分人群等位基因频率变化的事件,这在原始人群中并不典型
- 基因库
- 人群中所有个体携带的所有等位基因
- 遗传结构
- 不同可能的基因型在人群中的分布
- 宏观进化
- 古生物学时代出现的更大规模的进化变化
- 微进化
- 人口遗传结构的变化
- 现代合成
- 20世纪40年代形成并在今天被普遍接受的总体进化范式
- 种群遗传学
- 研究选择性力如何随着时间的推移改变人群中的等位基因频率