
15.4: 真核生物中的 RNA 处理
- Page ID
- 203047

培养技能
- 描述 RNA 处理的不同步骤
- 了解外显子、内含子和拼接的意义
- 解释 tRNA 和 rRNA 是如何处理的
转录后,真核生物 pre-mRNA 必须经过多个处理步骤才能被翻译。 真核(和原核生物)tRNA 和 rRNA 也要经过处理,然后才能在蛋白质合成机制中充当成分。
mRNA 处理
真核生物 pre-mRNA 在准备翻译之前要经过大量处理。 真核生物 mRNA 成熟所涉及的额外步骤会产生一种半衰期比原核生物 mRNA 长得多的分子。 真核生物 mRNA 持续数小时,而典型的大肠杆菌 mRNA 持续时间不超过五秒钟。
pre-mRNA 首先涂有 RNA 稳定蛋白;这些蛋白可以保护 pre-mRNA 在加工和输出到细胞核外时免受降解。 mRNA 前处理的三个最重要的步骤是在分子的 5' 和 3' 末端添加稳定和信号传导因子,以及去除未指定适当氨基酸的中间序列。 在极少数情况下,mRNA转录本在转录后可以 “编辑”。
进化连接:锥虫中的 RNA 编辑
锥虫是一组原生动物,包括病原体 Trypanosoma brucei,它会导致人类昏睡病(图\(\PageIndex{1}\))。 锥虫以及几乎所有其他真核生物都有称为线粒体的细胞器,为细胞提供化学能。 线粒体是表达自身DNA的细胞器,被认为是真核生物和被吞没的原核生物之间共生关系的残余物。 锥虫的线粒体DNA对于《中央教条》有一个有趣的例外:它们之前的mRNA没有正确的信息来指定功能蛋白。 通常,这是因为 mRNA 缺少几个 U 核苷酸。 该细胞执行一个称为RNA编辑的额外RNA处理步骤来纠正这种情况。
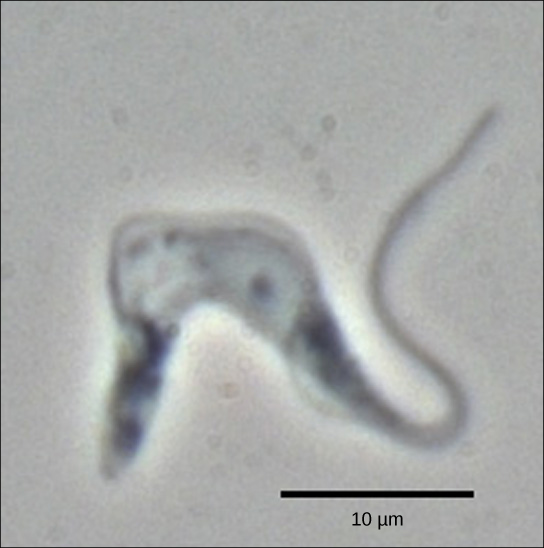
线粒体基因组中的其他基因编码 40 到 80 个核苷酸引导 RNA。 这些分子中的一个或多个通过互补碱基配对与 pre-mRNA 转录本中的某些核苷酸相互作用。 但是,引导 RNA 的 A 核苷酸比 mRNA 之前的 U 核苷酸要多。 在这些区域,引导 RNA 循环流出。 引导 RNA 的 3' 末端有一条长的 poly-U 尾巴,这些 U 碱基插入在引导 RNA 循环的 mRNA 前转录本区域。 这个过程完全由 RNA 分子介导。 也就是说,引导 RNA(而不是蛋白质)充当 RNA 编辑的催化剂。
RNA 编辑不仅仅是锥虫的现象。 在某些植物的线粒体中,几乎所有pre-mRNA都是经过编辑的。 在老鼠、兔子甚至人类等哺乳动物中也发现了RNA编辑。 mRNA前处理中这一额外步骤的进化原因可能是什么? 一种可能性是,作为古代原核生物残余物的线粒体具有同样古老的基于 RNA 的调节基因表达的方法。 为了支持这一假设,对pre-mRNA所做的编辑因细胞状况而异。 尽管是推测性的,但RNA编辑过程可能是原始时代的延迟,当时RNA分子而不是蛋白质是催化反应的原因。
5' 封顶
虽然前mRNA仍在合成中,但通过磷酸盐连接在生长转录本的5'末端添加了一个7-甲基鸟苷帽子。 该部分(功能组)保护新生的 mRNA 免受降解。 此外,蛋白质合成所涉及的因子可以识别上限,从而帮助启动核糖体的翻译。
3' Poly-A Tail
伸长完成后,pre-mRNA就会被AAUAA共识序列和富含GU的序列之间的核酸内切酶分开,而AAUAAA序列保留在pre-mRNA上。 然后,一种叫做 Poly-A 聚合酶的酶会添加一串大约 200 个 A 残基,称为 Poly-A 尾巴。 这种修饰进一步保护pre-mRNA免受降解,并发出转录本所需的细胞因子输出到细胞质的信号。
mRNA 前拼接
真核生物基因由外显子和称为内含子的 int ervening 序列(int - ron 表示它们是 int ervening 角色)和称为内含子的 int ervening 序列(int- ron 表示它们的 int ervening 作用)组成可能参与基因调控,但在处理过程中会从 pre-mRNA 中移除。 mRNA 中的内含子序列不编码功能蛋白。
内含子的发现令20世纪70年代的研究人员感到惊讶,他们预计,pre-mRNA可以在不经过进一步处理的情况下指定蛋白质序列,就像他们在原核生物中观察到的那样。 高等真核生物的基因通常包含一个或多个内含子。 这些区域可能对应于调控序列;但是,基因中有许多内含子或具有很长内含子的生物学意义尚不清楚。 内含子可能会减缓基因表达,因为转录含有大量内含子的 pre-mRNA 需要更长的时间。 或者,内含子可能是整个进化过程中古代基因融合遗留的非功能序列残留物。 单独的外显子通常编码单独的蛋白质亚单位或结构域这一事实支持了这一点。 在大多数情况下,内含子的序列可以在不影响蛋白质产物的情况下发生突变。
在蛋白质合成之前,必须完全、精确地去除所有 pre-mRNA 的内含子。 如果这个过程出现了哪怕是单个核苷酸的错误,那么重新连接的外显子的读取框架就会发生变化,由此产生的蛋白质就会失调。 移除内含子和重新连接外显子的过程称为拼接(图\(\PageIndex{2}\))。 当 pre-mRNA 仍在细胞核中时,内含子被移除并降解。 拼接是通过序列特异性机制进行的,该机制确保内含子被移除,外显子以单核苷酸的准确性和精度重新连接。 pre-mRNA 的拼接是由称为 spliceosomes 的蛋白质和 RNA 分子复合物进行的。
艺术连接
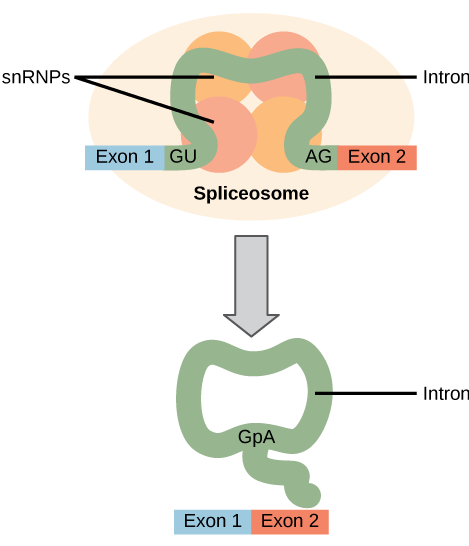
拼接错误与癌症和其他人类疾病有关。 哪些类型的突变可能导致拼接错误? 如果出现拼接错误,请考虑不同的可能结果。
请注意,可以存在 70 多个单独的内含子,每个内含子都必须经过拼接过程,除了 5' 封顶和添加 Poly-A 尾巴外,才能生成单个、可翻译的 mRNA 分子。
链接到学习
在此网站上查看如何在RNA拼接过程中去除内含子。
处理 tRNA 和 rRNA
tRNA 和 rRNA 是在蛋白质合成中起作用的结构分子;但是,这些 RNA 本身并没有被翻译。 pre-rRNA 在核仁中被转录、处理并组装成核糖体。 pre-tRNA 在细胞核中被转录和处理,然后释放到细胞质中,在那里它们与游离氨基酸连接以进行蛋白质合成。
真核生物和原核生物中的大多数 tRNA 和 rRNA 首先被转录为跨越多个 rRNA 或 tRNA 的长前体分子。 然后,酶将前体分解成与每个结构 RNA 相对应的亚单位。 pre-RRNA 的一些碱基是甲基化的;也就是说,为了稳定起见,添加了 —CH 3 部分(甲基官能团)。 tRNA 之前的分子也会经历甲基化。 与 pre-mRNA 一样,亚基切除发生在注定要成为 tRNA 或 rRNA 的真核生物前 RNA 中。
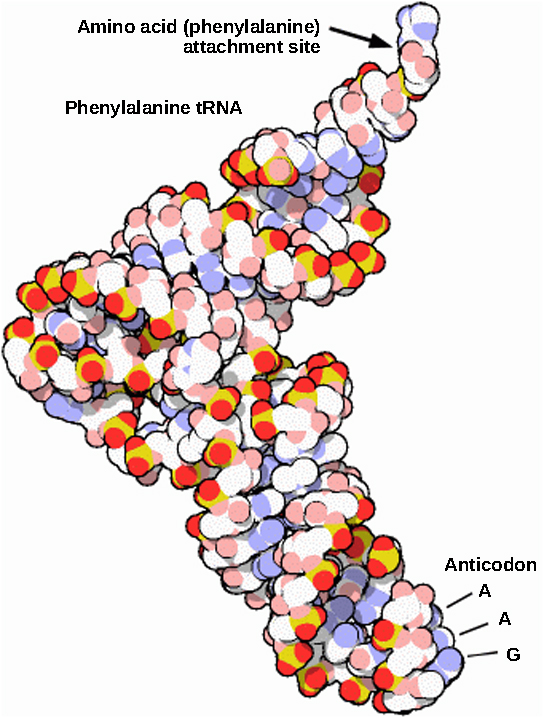
成熟的 rRNA 约占每个核糖体的50%。 核糖体的某些RNA分子纯粹是结构性的,而另一些则具有催化或结合活性。 成熟的 tRNA 通过分子内氢键呈现三维结构,将氨基酸结合位点定位在一端,将抗密码子定位在另一端(图\(\PageIndex{3}\))。 抗密码子是 tRNA 中的三核苷酸序列,通过互补碱基配对与 mRNA 密码子相互作用。
摘要
真核生物前 mRNA 经过修饰,带有 5' 甲基鸟苷帽和 poly-A 尾巴。 这些结构可保护成熟的 mRNA 免受降解,并有助于将其从细胞核中导出。 pre-mRNA 也会经过拼接,其中内含子被移除,外显子以单核苷酸的准确性重新连接。 只有经过5'封盖、3'聚腺苷酸化和内含子拼接的成品mRNA才能从细胞核输出到细胞质。 pre-rRNA 和 pre-tRNA 可以通过核苷酸的分子内裂解、剪接、甲基化和化学转化来处理。 在极少数情况下,在合成 mRNA 后,还会进行 RNA 编辑以插入缺失的碱基。
艺术联系
图\(\PageIndex{2}\):拼接错误与癌症和其他人类疾病有关。 哪些类型的突变可能导致拼接错误? 如果出现拼接错误,请考虑不同的可能结果。
- 回答
-
内含子两端的拼接体识别序列中的突变,或者构成拼接体的蛋白质和RNA中的突变,可能会损害拼接。 突变还可能添加新的剪接体识别位点。 拼接错误可能导致内含子被保留在拼接的RNA中,外显子被切除,或者拼接位点的位置发生变化。
词汇表
- 7-甲基鸟苷帽子
- 在 pre-mRNA 的 5' 末端添加了修改,以保护 mRNA 免受降解并协助翻译
- anticodon
- 对应于 mRNA 密码子的 tRNA 分子中的三核苷酸序列
- 外显子
- mRNA 前拼接完成后,蛋白质编码 mRNA 中存在序列
- 内含子
- 处理过程中从 mRNA 拼接的非蛋白质编码介入序列
- poly-a 尾巴
- 在 pre-mRNA 的 3' 末端添加了修饰,以保护 mRNA 免受降解并帮助 mRNA 从细胞核导出
- RNA 编辑
- 直接改变已经合成的 mRNA 中的一个或多个核苷酸
- 粘接
- 在 mRNA 之前移除内含子和重新连接外显子的过程